

An Overview of D7 Protein Structure and Physiological Roles in Blood-Feeding Nematocera

Abstract
:Simple Summary
Abstract
1. Introduction
2. D7s: From the Description of the First Gene to Clues Regarding Their Diversity, Distribution and Functions
3. Salivary D7 Proteins Are Anti-Inflammatory and Anti-Hemostatic Molecules
3.1. Contact Pathway Inhibitors
3.2. Biogenic Amine Binding D7s
3.3. Eicosanoid Binding D7s (the N-Terminal Domain of D7Ls)
3.4. ADP Binding D7s
4. Mosquito Juvenile Hormone-Binding Protein (mJHBP): What Is a D7-like Protein Doing in Mosquito Hemolymph?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ribeiro, J.M. Role of saliva in blood-feeding by arthropods. Annu. Rev. Entomol. 1987, 32, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Francischetti, I.M. Role of arthropod saliva in blood feeding: Sialome and post-sialome perspectives. Annu. Rev. Entomol. 2003, 48, 73–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graca-Souza, A.V.; Maya-Monteiro, C.; Paiva-Silva, G.O.; Braz, G.R.; Paes, M.C.; Sorgine, M.H.; Oliveira, M.F.; Oliveira, P.L. Adaptations against heme toxicity in blood-feeding arthropods. Insect Biochem. Mol. Biol. 2006, 36, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M. Blood-feeding arthropods: Live syringes or invertebrate pharmacologists? Infect. Agents Dis. 1995, 4, 143–152. [Google Scholar] [PubMed]
- Ribeiro, J.M.; Mans, B.J.; Arca, B. An insight into the sialome of blood-feeding Nematocera. Insect Biochem. Mol. Biol. 2010, 40, 767–784. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.C.; Arcà, B. Chapter 2 from Sialomes to the Sialoverse. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 59–118. [Google Scholar]
- Andersen, J.F. Structure and mechanism in salivary proteins from blood-feeding arthropods. Toxicon 2010, 56, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Arca, B.; Ribeiro, J.M. Saliva of hematophagous insects: A multifaceted toolkit. Curr. Opin. Insect Sci. 2018, 29, 102–109. [Google Scholar] [CrossRef]
- Cornwall, J.; Patton, W. Some observations on the salivary secretion of the commoner blood-sucking insects and ticks. Indian J. Med. Res. 1914, 2, 569–593. [Google Scholar]
- Lester, H.; Lloyd, L. Notes on the Process of Digestion in Tsetse-flies. Bull. Entomol. Res. 1928, 19, 39–60. [Google Scholar] [CrossRef]
- Ribeiro, J.; Garcia, E. The role of the salivary glands in feeding in Rhodnius prolixus. J. Exp. Biol. 1981, 94, 219–230. [Google Scholar] [CrossRef]
- Ribeiro, J.M.; Rossignol, P.A.; Spielman, A. Role of mosquito saliva in blood vessel location. J. Exp. Biol. 1984, 108, 1–7. [Google Scholar] [CrossRef]
- Ribeiro, J.; Rossignol, P.; Spielman, A. Salivary gland apyrase determines probing time in anopheline mosquitoes. J. Insect Physiol. 1985, 31, 689–692. [Google Scholar] [CrossRef]
- Valenzuela, J.G. High-throughput approaches to study salivary proteins and genes from vectors of disease. Insect Biochem. Mol. Biol. 2002, 32, 1199–1209. [Google Scholar] [CrossRef]
- Mans, B.; Francischetti, I. Sialomic perspectives on the evolution of blood-feeding behavior in Arthropods: Future therapeutics by natural design. In Toxins and Hemostasis; Springer: Dordrecht, The Netherlands, 2010; pp. 21–44. [Google Scholar]
- Mans, B.J. Evolution of vertebrate hemostatic and inflammatory control mechanisms in blood-feeding arthropods. J. Innate Immun. 2011, 3, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, J.G.; Charlab, R.; Gonzalez, E.C.; de Miranda-Santos, I.K.; Marinotti, O.; Francischetti, I.M.; Ribeiro, J.M. The D7 family of salivary proteins in blood sucking diptera. Insect Mol. Biol. 2002, 11, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Calvo, E.; de Bianchi, A.G.; James, A.A.; Marinotti, O. The major acid soluble proteins of adult female Anopheles darlingi salivary glands include a member of the D7-related family of proteins. Insect Biochem. Mol. Biol. 2002, 32, 1419–1427. [Google Scholar] [CrossRef]
- Malafronte Rdos, S.; Calvo, E.; James, A.A.; Marinotti, O. The major salivary gland antigens of Culex quinquefasciatus are D7-related proteins. Insect Biochem. Mol. Biol. 2003, 33, 63–71. [Google Scholar] [CrossRef]
- Rihani, K.; Ferveur, J.F.; Briand, L. The 40-Year Mystery of Insect Odorant-Binding Proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. Camb. Philos. Soc. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.S.; Xiao, S.; Carlson, J.R. The diverse small proteins called odorant-binding proteins. Open Biol. 2018, 8, 180208. [Google Scholar] [CrossRef] [Green Version]
- Manoharan, M.; Ng Fuk Chong, M.; Vaitinadapoule, A.; Frumence, E.; Sowdhamini, R.; Offmann, B. Comparative genomics of odorant binding proteins in Anopheles gambiae, Aedes aegypti, and Culex quinquefasciatus. Genome Biol. Evol. 2013, 5, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Wogulis, M.; Morgan, T.; Ishida, Y.; Leal, W.S.; Wilson, D.K. The crystal structure of an odorant binding protein from Anopheles gambiae: Evidence for a common ligand release mechanism. Biochem. Biophys. Res. Commun. 2006, 339, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Arca, B.; Lombardo, F.; Lanfrancotti, A.; Spanos, L.; Veneri, M.; Louis, C.; Coluzzi, M. A cluster of four D7-related genes is expressed in the salivary glands of the African malaria vector Anopheles gambiae. Insect Mol. Biol. 2002, 11, 47–55. [Google Scholar] [CrossRef]
- Mans, B.J.; Calvo, E.; Ribeiro, J.M.; Andersen, J.F. The crystal structure of D7r4, a salivary biogenic amine-binding protein from the malaria mosquito Anopheles gambiae. J. Biol. Chem. 2007, 282, 36626–36633. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, P.H.; Dias, D.R.; Xu, X.; Francischetti, I.M.B.; Gittis, A.G.; Arp, G.; Garboczi, D.N.; Ribeiro, J.M.C.; Andersen, J.F. Functional aspects of evolution in a cluster of salivary protein genes from mosquitoes. Insect Biochem. Mol. Biol. 2022, 146, 103785. [Google Scholar] [CrossRef]
- Alvarenga, P.H.; Francischetti, I.M.; Calvo, E.; Sa-Nunes, A.; Ribeiro, J.M.; Andersen, J.F. The function and three-dimensional structure of a thromboxane A2/cysteinyl leukotriene-binding protein from the saliva of a mosquito vector of the malaria parasite. PLoS Biol. 2010, 8, e1000547. [Google Scholar] [CrossRef] [Green Version]
- James, A.A.; Blackmer, K.; Marinotti, O.; Ghosn, C.R.; Racioppi, J.V. Isolation and characterization of the gene expressing the major salivary gland protein of the female mosquito, Aedes aegypti. Mol. Biochem. Parasitol. 1991, 44, 245–253. [Google Scholar] [CrossRef]
- Arca, B.; Lombardo, F.; de Lara Capurro, M.; della Torre, A.; Dimopoulos, G.; James, A.A.; Coluzzi, M. Trapping cDNAs encoding secreted proteins from the salivary glands of the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. USA 1999, 96, 1516–1521. [Google Scholar] [CrossRef] [Green Version]
- Suwan, N.; Wilkinson, M.C.; Crampton, J.M.; Bates, P.A. Expression of D7 and D7-related proteins in the salivary glands of the human malaria mosquito Anopheles stephensi. Insect Mol. Biol. 2002, 11, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Calvo, E.; Mans, B.J.; Andersen, J.F.; Ribeiro, J.M. Function and evolution of a mosquito salivary protein family. J. Biol. Chem. 2006, 281, 1935–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isawa, H.; Yuda, M.; Orito, Y.; Chinzei, Y. A mosquito salivary protein inhibits activation of the plasma contact system by binding to factor XII and high molecular weight kininogen. J. Biol. Chem. 2002, 277, 27651–27658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo, E.; Mans, B.J.; Ribeiro, J.M.; Andersen, J.F. Multifunctionality and mechanism of ligand binding in a mosquito antiinflammatory protein. Proc. Natl. Acad. Sci. USA 2009, 106, 3728–3733. [Google Scholar] [CrossRef] [Green Version]
- Martin-Martin, I.; Kern, O.; Brooks, S.; Smith, L.B.; Valenzuela-Leon, P.C.; Bonilla, B.; Ackerman, H.; Calvo, E. Biochemical characterization of AeD7L2 and its physiological relevance in blood feeding in the dengue mosquito vector, Aedes aegypti. FEBS J. 2021, 288, 2014–2029. [Google Scholar] [CrossRef]
- Martin-Martin, I.; Smith, L.B.; Chagas, A.C.; Sa-Nunes, A.; Shrivastava, G.; Valenzuela-Leon, P.C.; Calvo, E. Aedes albopictus D7 Salivary Protein Prevents Host Hemostasis and Inflammation. Biomolecules 2020, 10, 1372. [Google Scholar] [CrossRef]
- Martin-Martin, I.; Paige, A.; Valenzuela Leon, P.C.; Gittis, A.G.; Kern, O.; Bonilla, B.; Chagas, A.C.; Ganesan, S.; Smith, L.B.; Garboczi, D.N.; et al. ADP binding by the Culex quinquefasciatus mosquito D7 salivary protein enhances blood feeding on mammals. Nat. Commun. 2020, 11, 2911. [Google Scholar] [CrossRef]
- Jablonka, W.; Kim, I.H.; Alvarenga, P.H.; Valenzuela, J.G.; Ribeiro, J.M.C.; Andersen, J.F. Functional and structural similarities of D7 proteins in the independently-evolved salivary secretions of sand flies and mosquitoes. Sci. Rep. 2019, 9, 5340. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, P.H.; Xu, X.; Oliveira, F.; Chagas, A.C.; Nascimento, C.R.; Francischetti, I.M.; Juliano, M.A.; Juliano, L.; Scharfstein, J.; Valenzuela, J.G.; et al. Novel family of insect salivary inhibitors blocks contact pathway activation by binding to polyphosphate, heparin, and dextran sulfate. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2759–2770. [Google Scholar] [CrossRef] [Green Version]
- Isawa, H.; Orito, Y.; Iwanaga, S.; Jingushi, N.; Morita, A.; Chinzei, Y.; Yuda, M. Identification and characterization of a new kallikrein-kinin system inhibitor from the salivary glands of the malaria vector mosquito Anopheles stephensi. Insect Biochem. Mol. Biol. 2007, 37, 466–477. [Google Scholar] [CrossRef]
- Kato, H.; Anderson, J.M.; Kamhawi, S.; Oliveira, F.; Lawyer, P.G.; Pham, V.M.; Sangare, C.S.; Samake, S.; Sissoko, I.; Garfield, M.; et al. High degree of conservancy among secreted salivary gland proteins from two geographically distant Phlebotomus duboscqi sandflies populations (Mali and Kenya). BMC Genom. 2006, 7, 226. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Jochim, R.C.; Gomez, E.A.; Uezato, H.; Mimori, T.; Korenaga, M.; Sakurai, T.; Katakura, K.; Valenzuela, J.G.; Hashiguchi, Y. Analysis of salivary gland transcripts of the sand fly Lutzomyia ayacuchensis, a vector of Andean-type cutaneous leishmaniasis. Infect. Genet. Evol. 2013, 13, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, J.G.; Garfield, M.; Rowton, E.D.; Pham, V.M. Identification of the most abundant secreted proteins from the salivary glands of the sand fly Lutzomyia longipalpis, vector of Leishmania chagasi. J. Exp. Biol. 2004, 207, 3717–3729. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, J.G.; Belkaid, Y.; Garfield, M.K.; Mendez, S.; Kamhawi, S.; Rowton, E.D.; Sacks, D.L.; Ribeiro, J.M. Toward a defined anti-Leishmania vaccine targeting vector antigens: Characterization of a protective salivary protein. J. Exp. Med. 2001, 194, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, F.; Rowton, E.; Aslan, H.; Gomes, R.; Castrovinci, P.A.; Alvarenga, P.H.; Abdeladhim, M.; Teixeira, C.; Meneses, C.; Kleeman, L.T.; et al. A sand fly salivary protein vaccine shows efficacy against vector-transmitted cutaneous leishmaniasis in nonhuman primates. Sci. Transl. Med. 2015, 7, 290ra90. [Google Scholar] [CrossRef] [Green Version]
- Renne, T. The procoagulant and proinflammatory plasma contact system. Semin. Immunopathol. 2012, 34, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Vogler, E.A.; Siedlecki, C.A. Contact activation of blood-plasma coagulation. Biomaterials 2009, 30, 1857–1869. [Google Scholar] [CrossRef] [Green Version]
- Renne, T.; Pozgajova, M.; Gruner, S.; Schuh, K.; Pauer, H.U.; Burfeind, P.; Gailani, D.; Nieswandt, B. Defective thrombus formation in mice lacking coagulation factor XII. J. Exp. Med. 2005, 202, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Grover, S.P.; Mackman, N. Intrinsic Pathway of Coagulation and Thrombosis. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 331–338. [Google Scholar] [CrossRef]
- Gailani, D.; Renne, T. Intrinsic pathway of coagulation and arterial thrombosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2507–2513. [Google Scholar] [CrossRef]
- Mailer, R.K.; Rangaswamy, C.; Konrath, S.; Emsley, J.; Renne, T. An update on factor XII-driven vascular inflammation. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119166. [Google Scholar] [CrossRef] [PubMed]
- Oschatz, C.; Maas, C.; Lecher, B.; Jansen, T.; Bjorkqvist, J.; Tradler, T.; Sedlmeier, R.; Burfeind, P.; Cichon, S.; Hammerschmidt, S.; et al. Mast cells increase vascular permeability by heparin-initiated bradykinin formation in vivo. Immunity 2011, 34, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Dray, A.; Perkins, M. Bradykinin and inflammatory pain. Trends Neurosci. 1993, 16, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E.; Kitching, I.J. The phylogeny of Anophelinae revisited: Inferences about the origin and classification of Anopheles (Diptera: Culicidae). Zool. Scr. 2016, 45, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Giraldo-Calderon, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; VectorBase, C.; Madey, G.; et al. VectorBase: An updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef]
- Neafsey, D.E.; Waterhouse, R.M.; Abai, M.R.; Aganezov, S.S.; Alekseyev, M.A.; Allen, J.E.; Amon, J.; Arca, B.; Arensburger, P.; Artemov, G.; et al. Mosquito genomics. Highly evolvable malaria vectors: The genomes of 16 Anopheles mosquitoes. Science 2015, 347, 1258522. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.M.C.; Wides, R.; et al. The genome sequence of the malaria mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef]
- Arca, B.; Lombardo, F.; Struchiner, C.J.; Ribeiro, J.M. Anopheline salivary protein genes and gene families: An evolutionary overview after the whole genome sequence of sixteen Anopheles species. BMC Genom. 2017, 18, 153. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Horsmanheimo, L.; Harvima, I.T.; Harvima, R.J.; Brummer-Korvenkontio, H.; Francois, G.; Reunala, T. Histamine and leukotriene C4 release in cutaneous mosquito-bite reactions. J. Allergy Clin. Immunol. 1996, 98, 408–411. [Google Scholar] [CrossRef]
- Bisgaard, H.; Lerche, A.; Kristensen, J.K. Leukotriene- and histamine-induced increases in vascular permeability and interstitial transport in the skin. J. Investig. Dermatol. 1985, 84, 427–429. [Google Scholar] [CrossRef] [PubMed]
- Ashina, K.; Tsubosaka, Y.; Nakamura, T.; Omori, K.; Kobayashi, K.; Hori, M.; Ozaki, H.; Murata, T. Histamine Induces Vascular Hyperpermeability by Increasing Blood Flow and Endothelial Barrier Disruption In Vivo. PLoS ONE 2015, 10, e0132367. [Google Scholar] [CrossRef] [PubMed]
- Vander Does, A.; Labib, A.; Yosipovitch, G. Update on mosquito bite reaction: Itch and hypersensitivity, pathophysiology, prevention, and treatment. Front. Immunol. 2022, 13, 1024559. [Google Scholar] [CrossRef] [PubMed]
- Twycross, R.; Greaves, M.W.; Handwerker, H.; Jones, E.A.; Libretto, S.E.; Szepietowski, J.C.; Zylicz, Z. Itch: Scratching more than the surface. QJM 2003, 96, 7–26. [Google Scholar] [CrossRef]
- Ohtsuka, E.; Kawai, S.; Ichikawa, T.; Nojima, H.; Kitagawa, K.; Shirai, Y.; Kamimura, K.; Kuraishi, Y. Roles of mast cells and histamine in mosquito bite-induced allergic itch-associated responses in mice. Jpn. J. Pharmacol. 2001, 86, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.B.; Duge, E.; Valenzuela-Leon, P.C.; Brooks, S.; Martin-Martin, I.; Ackerman, H.; Calvo, E. Novel salivary antihemostatic activities of long-form D7 proteins from the malaria vector Anopheles gambiae facilitate hematophagy. J. Biol. Chem. 2022, 298, 101971. [Google Scholar] [CrossRef]
- Abdeladhim, M.; Coutinho-Abreu, I.V.; Townsend, S.; Pasos-Pinto, S.; Sanchez, L.; Rasouli, M.; Guimaraes-Costa, A.B.; Aslan, H.; Francischetti, I.M.B.; Oliveira, F.; et al. Molecular Diversity between Salivary Proteins from New World and Old World Sand Flies with Emphasis on Bichromomyia olmeca, the Sand Fly Vector of Leishmania mexicana in Mesoamerica. PLoS Negl. Trop. Dis. 2016, 10, e0004771. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, D.A.; Engel, M.S. Evolution of the Insects; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2005; p. xv. 755p. [Google Scholar]
- Xu, X.; Oliveira, F.; Chang, B.W.; Collin, N.; Gomes, R.; Teixeira, C.; Reynoso, D.; My Pham, V.; Elnaiem, D.E.; Kamhawi, S.; et al. Structure and function of a “yellow” protein from saliva of the sand fly Lutzomyia longipalpis that confers protective immunity against Leishmania major infection. J. Biol. Chem. 2011, 286, 32383–32393. [Google Scholar] [CrossRef] [Green Version]
- Luna-Gomes, T.; Bozza, P.T.; Bandeira-Melo, C. Eosinophil recruitment and activation: The role of lipid mediators. Front. Pharmacol. 2013, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Camp, R.D.; Coutts, A.A.; Greaves, M.W.; Kay, A.B.; Walport, M.J. Responses of human skin to intradermal injection of leukotrienes C4, D4 and B4. Br. J. Pharmacol. 1983, 80, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Soter, N.A.; Lewis, R.A.; Corey, E.J.; Austen, K.F. Local effects of synthetic leukotrienes (LTC4, LTD4, LTE4, and LTB4) in human skin. J. Investig. Dermatol. 1983, 80, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Offermanns, S. Activation of platelet function through G protein-coupled receptors. Circ. Res. 2006, 99, 1293–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francischetti, I.M. Platelet aggregation inhibitors from hematophagous animals. Toxicon 2010, 56, 1130–1144. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Ebina, S.; Nakanishi, H.; Nakahata, N. Thromboxane A2 receptor-mediated signal transduction in rabbit aortic smooth muscle cells. Gen. Pharmacol. 1995, 26, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, N. Thromboxane A2: Physiology/pathophysiology, cellular signal transduction and pharmacology. Pharmacol. Ther. 2008, 118, 18–35. [Google Scholar] [CrossRef]
- Andoh, T.; Nishikawa, Y.; Yamaguchi-Miyamoto, T.; Nojima, H.; Narumiya, S.; Kuraishi, Y. Thromboxane A2 induces itch-associated responses through TP receptors in the skin in mice. J. Investig. Dermatol. 2007, 127, 2042–2047. [Google Scholar] [CrossRef] [Green Version]
- Andoh, T.; Li, S.; Uta, D. Involvement of thromboxane A2 in interleukin-31-induced itch-associated response in mice. Pharmacol. Rep. 2018, 70, 251–257. [Google Scholar] [CrossRef]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori Odorant-binding proteins reveals that a general odorant-binding protein discriminates between sex pheromone components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Bucci, B.K.; Kruse, S.W.; Thode, A.B.; Alvarado, S.M.; Jones, D.N. Effect of n-alcohols on the structure and stability of the Drosophila odorant binding protein LUSH. Biochemistry 2006, 45, 1693–1701. [Google Scholar] [CrossRef]
- Kim, M.S.; Repp, A.; Smith, D.P. LUSH odorant-binding protein mediates chemosensory responses to alcohols in Drosophila melanogaster. Genetics 1998, 150, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Murphy, E.J.; Nix, J.C.; Jones, D.N.M. Aedes aegypti Odorant Binding Protein 22 selectively binds fatty acids through a conformational change in its C-terminal tail. Sci. Rep. 2020, 10, 3300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Picimbon, J.F.; Ji, S.; Kan, Y.; Chuanling, Q.; Zhou, J.J.; Pelosi, P. Multiple functions of an odorant-binding protein in the mosquito Aedes aegypti. Biochem. Biophys. Res. Commun. 2008, 372, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Sarti, A.C.; Coutinho-Silva, R. Purinergic signaling, DAMPs, and inflammation. Am. J. Physiol. Cell Physiol. 2020, 318, C832–C835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugappa, S.; Kunapuli, S.P. The role of ADP receptors in platelet function. Front. Biosci. 2006, 11, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Pham, V.; Jablonka, W.; Goodman, W.G.; Ribeiro, J.M.C.; Andersen, J.F. A mosquito hemolymph odorant-binding protein family member specifically binds juvenile hormone. J. Biol. Chem. 2017, 292, 15329–15339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, R.; Fujimoto, Z.; Shiotsuki, T.; Tsuchiya, W.; Momma, M.; Tase, A.; Miyazawa, M.; Yamazaki, T. Structural mechanism of JH delivery in hemolymph by JHBP of silkworm, Bombyx mori. Sci. Rep. 2011, 1, 133. [Google Scholar] [CrossRef] [Green Version]
- Jindra, M.; Palli, S.R.; Riddiford, L.M. The juvenile hormone signaling pathway in insect development. Annu. Rev. Entomol. 2013, 58, 181–204. [Google Scholar] [CrossRef]
- Nakamura, A.; Stiebler, R.; Fantappie, M.R.; Fialho, E.; Masuda, H.; Oliveira, M.F. Effects of retinoids and juvenoids on moult and on phenoloxidase activity in the blood-sucking insect Rhodnius prolixus. Acta Trop. 2007, 103, 222–230. [Google Scholar] [CrossRef]
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory Pathways Controlling Female Insect Reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef]
- Hernandez-Martinez, S.; Cardoso-Jaime, V.; Nouzova, M.; Michalkova, V.; Ramirez, C.E.; Fernandez-Lima, F.; Noriega, F.G. Juvenile hormone controls ovarian development in female Anopheles albimanus mosquitoes. Sci. Rep. 2019, 9, 2127. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, D.O.; de Paula, S.O.; Zanuncio, J.C.; Martinez, L.C.; Serrao, J.E. Juvenile hormone downregulates vitellogenin production in Ectatomma tuberculatum (Hymenoptera: Formicidae) sterile workers. J. Exp. Biol. 2016, 219, 103–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.M.; Wang, Y.H.; Yang, Q.T.; Wang, X.L.; Wang, M.; Raikhel, A.S.; Zou, Z. Regulation of antimicrobial peptides by juvenile hormone and its receptor, Methoprene-tolerant, in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2021, 128, 103509. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Guo, E.; Diao, Y.; Zhou, S.; Peng, Q.; Cao, Y.; Ling, E.; Li, S. Genome-wide regulation of innate immunity by juvenile hormone and 20-hydroxyecdysone in the Bombyx fat body. BMC Genom. 2010, 11, 549. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Castillo, J.C.; Aryan, A.; Martin-Martin, I.; Nouzova, M.; Noriega, F.G.; Barletta, A.B.F.; Calvo, E.; Adelman, Z.N.; Ribeiro, J.M.C.; et al. A mosquito juvenile hormone binding protein (mJHBP) regulates the activation of innate immune defenses and hemocyte development. PLoS Pathog. 2020, 16, e1008288. [Google Scholar] [CrossRef] [Green Version]
- Mans, B.J.; Ribeiro, J.M.; Andersen, J.F. Structure, function, and evolution of biogenic amine-binding proteins in soft ticks. J. Biol. Chem. 2008, 283, 18721–18733. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.F.; Francischetti, I.M.; Valenzuela, J.G.; Schuck, P.; Ribeiro, J.M. Inhibition of hemostasis by a high affinity biogenic amine-binding protein from the saliva of a blood-feeding insect. J. Biol. Chem. 2003, 278, 4611–4617. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.F.; Gudderra, N.P.; Francischetti, I.M.; Ribeiro, J.M. The role of salivary lipocalins in blood feeding by Rhodnius prolixus. Arch. Insect Biochem. Physiol. 2005, 58, 97–105. [Google Scholar] [CrossRef]
- Jablonka, W.; Pham, V.; Nardone, G.; Gittis, A.; Silva-Cardoso, L.; Atella, G.C.; Ribeiro, J.M.; Andersen, J.F. Structure and Ligand-Binding Mechanism of a Cysteinyl Leukotriene-Binding Protein from a Blood-Feeding Disease Vector. ACS Chem. Biol. 2016, 11, 1934–1944. [Google Scholar] [CrossRef] [Green Version]
- Assumpcao, T.C.; Alvarenga, P.H.; Ribeiro, J.M.; Andersen, J.F.; Francischetti, I.M. Dipetalodipin, a novel multifunctional salivary lipocalin that inhibits platelet aggregation, vasoconstriction, and angiogenesis through unique binding specificity for TXA2, PGF2alpha, and 15(S)-HETE. J. Biol. Chem. 2010, 285, 39001–39012. [Google Scholar] [CrossRef] [Green Version]
- Mans, B.J.; Ribeiro, J.M. A novel clade of cysteinyl leukotriene scavengers in soft ticks. Insect Biochem. Mol. Biol. 2008, 38, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Mans, B.J.; Ribeiro, J.M. Function, mechanism and evolution of the moubatin-clade of soft tick lipocalins. Insect Biochem. Mol. Biol. 2008, 38, 841–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu-Daude, F.; Claverie, A.; Plichart, C.; Boulanger, D.; Mphande, F.A.; Bossin, H.C. Specific human antibody responses to Aedes aegypti and Aedes polynesiensis saliva: A new epidemiological tool to assess human exposure to disease vectors in the Pacific. PLoS Negl. Trop. Dis. 2018, 12, e0006660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oseno, B.; Marura, F.; Ogwang, R.; Muturi, M.; Njunge, J.; Nkumama, I.; Mwakesi, R.; Mwai, K.; Rono, M.K.; Mwakubambanya, R.; et al. Characterization of Anopheles gambiae D7 salivary proteins as markers of human-mosquito bite contact. Parasites Vectors 2022, 15, 11. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
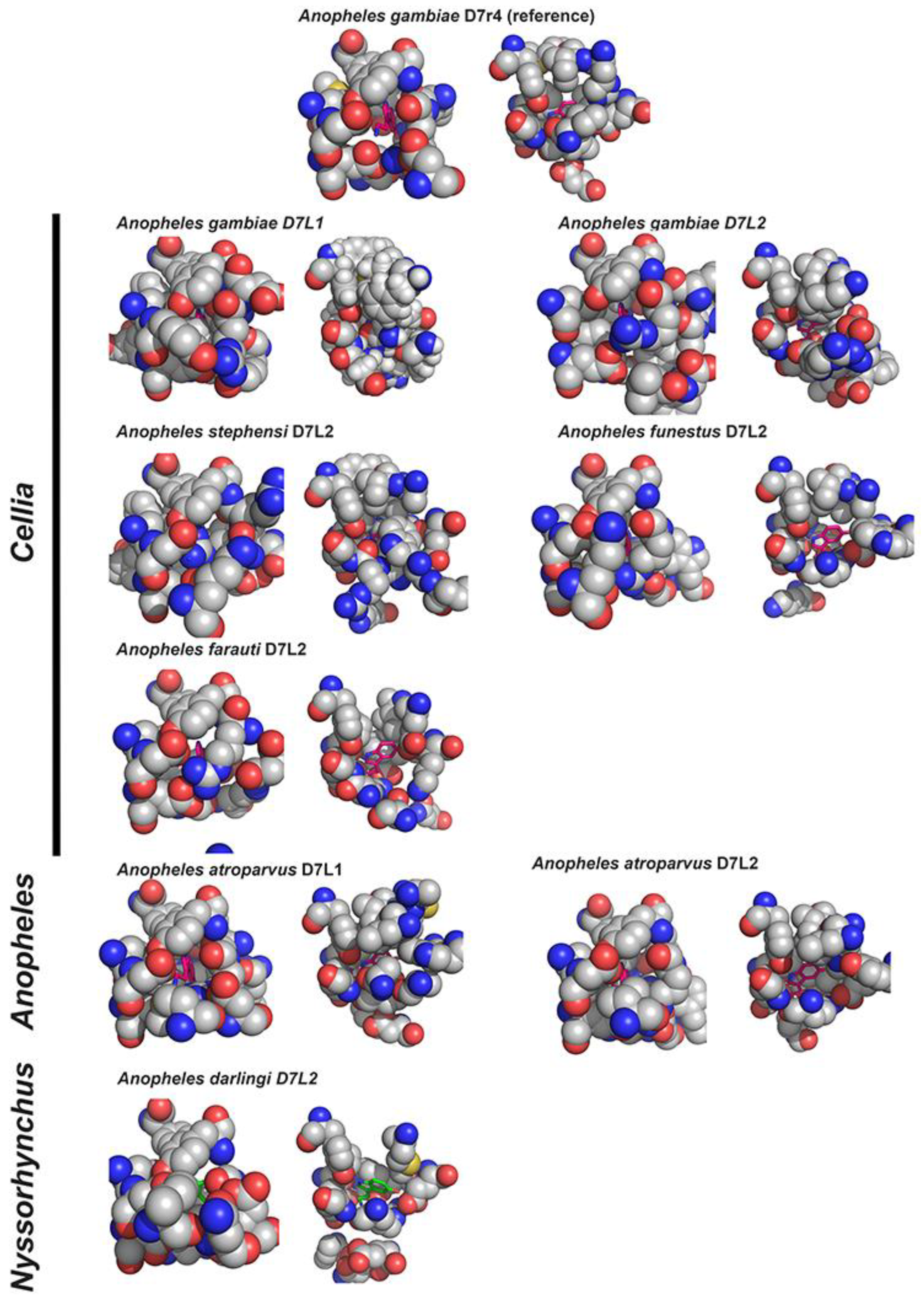{kind=link}
| Protein (Accession Number) | Ligands (KD in nM, When Available) | PDB | References |
|---|---|---|---|
| An. gambiae D7r1 (AGAP008284) | Serotonin (1.99) | [34] | |
| Histamine (103) | |||
| An. gambiae D7r2 (AGAP008282) | Serotonin (2.93) | [34] | |
| Norepinephrine (2.84) | |||
| Histamine (90) | |||
| Epinephrine (64) | |||
| An. gambiae D7r3 (AGAP008283) | Serotonin (0.16) | [34] | |
| Norepinephrine (3.19) | |||
| Histamine (41.0) | |||
| Epinephrine (312.0) | |||
| An. gambiae D7r4 (AGAP008281) | ** | 2QEV | [28,34] |
| Serotonin (0.93) | 2QEH | ||
| Norepinephrine (645) | 2QEO | ||
| Histamine (111) | 2QEB | ||
| Tryptamine | 2PQL | ||
| An. gambiae D7r5 (AGAP008280) | No binding to any biogenic amine tested | [34] | |
| An. stephensi D7r1 (hamadarin) (ASTE016512) | Factor XII | [35] | |
| High Molecular Weight Kininogen | |||
| Aedes D7S1 (AAEL006406) | No binding to any biogenic amine tested | 7TVC | [29] |
| Cu. quinquefasciatus D7S (D7CQ1) (AAR18437.1) | No binding to any biogenic amine tested | 7TVY | [29] |
| An. gambiae D7L3 (AGAP028120) | Serotonin (22.2) | [29] | |
| Histamine (1536.1) | |||
| Norepinephrine (2531.6) | |||
| No binding to tryptamine, octopamine, dopamine or epinephrine | |||
| No binding to any eicosanoid tested or ADP | |||
| An. stephensi D7L2 (previously D7L1) (AF420266.1) | ** | 3NGV | [30] |
| LTC4 (3.7) | 3NHI | ||
| LTD4 (5.0) | |||
| LTE4 (6.1) | |||
| U46619 (98.0) | 3NHT | ||
| Carboxyclic thromboxane A2 (38.3) | |||
| PGD2 (1500.0) | |||
| PGE2 (1900.00) | |||
| PGF2a (671.0) | |||
| U51605 (775.0) | |||
| No binding to any biogenic amine tested, LTB4 or ADP. | |||
| An. atroparvus D7L1 (AATE004070) | Serotonin (2.9) | [29] | |
| Tryptamine (<2) | |||
| Histamine (548.8) | |||
| LTC4 (257.0) | |||
| LTD4 (460.8) | |||
| LTE4 (344.8) | |||
| No binding to dopamine, octopamine, norepinephrine or epinephrine | |||
| No binding to U46619 or any other eicosanoid tested | |||
| An. darlingi D7L2 (EU934268.1) | ** | 7U1N | [29] |
| Serotonin (17.5) | 7TX8 | ||
| Tryptamine (518.1) | |||
| Dopamine (833.3) | |||
| Octopamine (1113.5) | |||
| Norepinephrine (3257.3) | |||
| LTC4 (13.8) | |||
| LTD4 (26.1) | |||
| LTE4 (243.9) | |||
| U46619 (91.7) | 7TX8 | ||
| No binding to histamine, epinephrine or LTB4 | |||
| Ae. aegypti D7L 1 (AAEL006424) | ** | 3DXL | [34,36] |
| Serotonin (0.39) | |||
| Norepinephrine (0.119) | 3DYE | ||
| Histamine (140) | |||
| Epinephrine (102) | |||
| LTC4 (57.4) | |||
| LTD4 (54.3) | |||
| LTE4 (60.2) | 3DZT | ||
| LTB4 (140) | |||
| No binding to U46619 | |||
| Ae. aegypti D7L2 (AAEL006417) | Serotonin (1.68) | [37] | |
| Norepinephrine (110) | |||
| Histamine (1130) | |||
| LTC4 (5270) | |||
| LTD4 (597) | |||
| LTE4 (1930) | |||
| LTB4 (769) | |||
| U46619 (69.4) | |||
| No binding to epinephrine | |||
| Ae albopictus D7L1 (AALF024478) | Serotonin (4.51) | [38] | |
| Norepinephrine (3.67) | |||
| Histamine (278) | |||
| Epinephrine (4110) | |||
| Dopamine (11) | |||
| Tryptamine (570) | |||
| LTC4 (67.7) | |||
| LTD4 (332) | |||
| LTE4 (567) | |||
| LTB4 (342) | |||
| U46619 (946) | |||
| Cu. quinquefasciatus D7L1 (CPIJ014549) | 5’-ATP (30.77) | [39] | |
| 5’-ADP (32.68) | 6V4C | ||
| 5’-AMP (77.52) | |||
| Adenosine (312.50) | |||
| Adenine (1760.56) | |||
| No binding to 5’-GTP, 5’-TTP, 3’-AMP, cAMP or PolyP | |||
| No binding to any biogenic amine or eicosanoid tested | |||
| Cu. quinquefasciatus D7L2 (CPIJ014551) | Serotonin (7.46) | [39] | |
| Histamine (383.14) | |||
| Epinephrine (226.24) | |||
| LTC4 (151.75) | |||
| LTD4 (156.49) | |||
| LTE4 (158.73) | |||
| U46619 (934.58) | |||
| Arachidonic acid (1083.42) | |||
| No binding to U46619, 5’ATP or 5’-ADP | |||
| P. papatasi D7L (AGE83092) | LTC4 (5.88) | [40] | |
| LTD4 (1.75) | |||
| LTE4 (15.87) | |||
| U46619 (751.8) | |||
| No binding to any biogenic amine tested | |||
| P. duboscqi D7L (ABI15936) | ** | 6MTF, 6MT7 | [40] |
| LTC4 (3.2) | |||
| LTD4 (4.16) | |||
| LTE4 (29.4) | |||
| U46619 (1282.0) | |||
| No binding to any biogenic amine tested | |||
| P. duboscqi Salivary protein 15 a and b PDSP15a (ABI15933) PDSP15b (ABI15943) | ** | 4OZD | [41] |
| Polyphosphate | |||
| Dextran sulfate | |||
| Heparin | |||
| No binding to FXII, FXIIa, FXI, prekallikrein or kallikrein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarenga, P.H.; Andersen, J.F. An Overview of D7 Protein Structure and Physiological Roles in Blood-Feeding Nematocera. Biology 2023, 12, 39. https://doi.org/10.3390/biology12010039
Alvarenga PH, Andersen JF. An Overview of D7 Protein Structure and Physiological Roles in Blood-Feeding Nematocera. Biology. 2023; 12(1):39. https://doi.org/10.3390/biology12010039
Chicago/Turabian StyleAlvarenga, Patricia H., and John F. Andersen. 2023. "An Overview of D7 Protein Structure and Physiological Roles in Blood-Feeding Nematocera" Biology 12, no. 1: 39. https://doi.org/10.3390/biology12010039