
Probiotic Adhesion to Skin Keratinocytes and Underlying Mechanisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
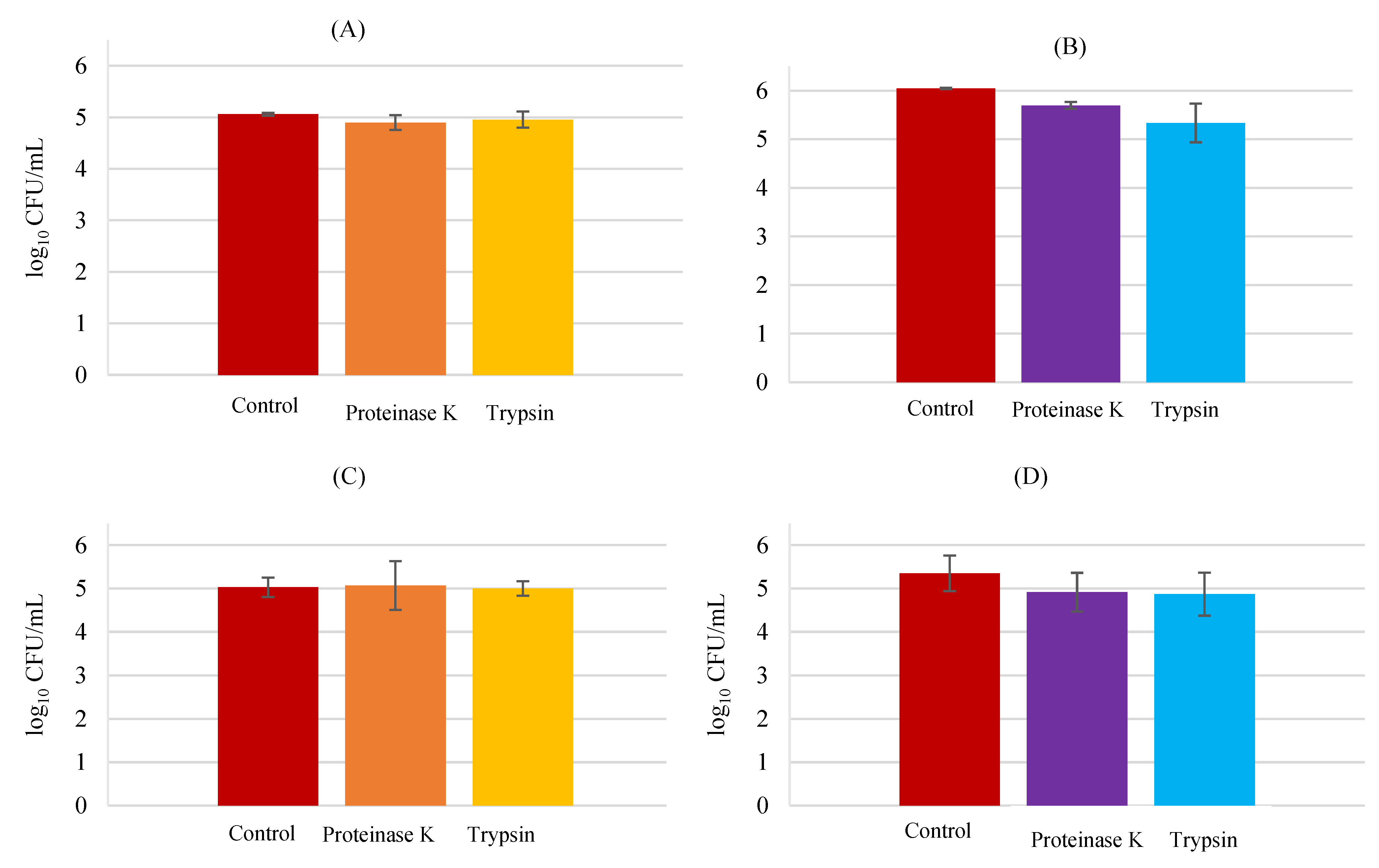{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Cell Culture Assays (HaCaT)
2.3. Invasion Assays
2.4. Adhesion Assays
2.5. Carbohydrate Analysis
2.6. Protein Analysis
2.7. Human Skin Equivalent Assays
2.8. Statistical Analysis
3. Results and Discussion
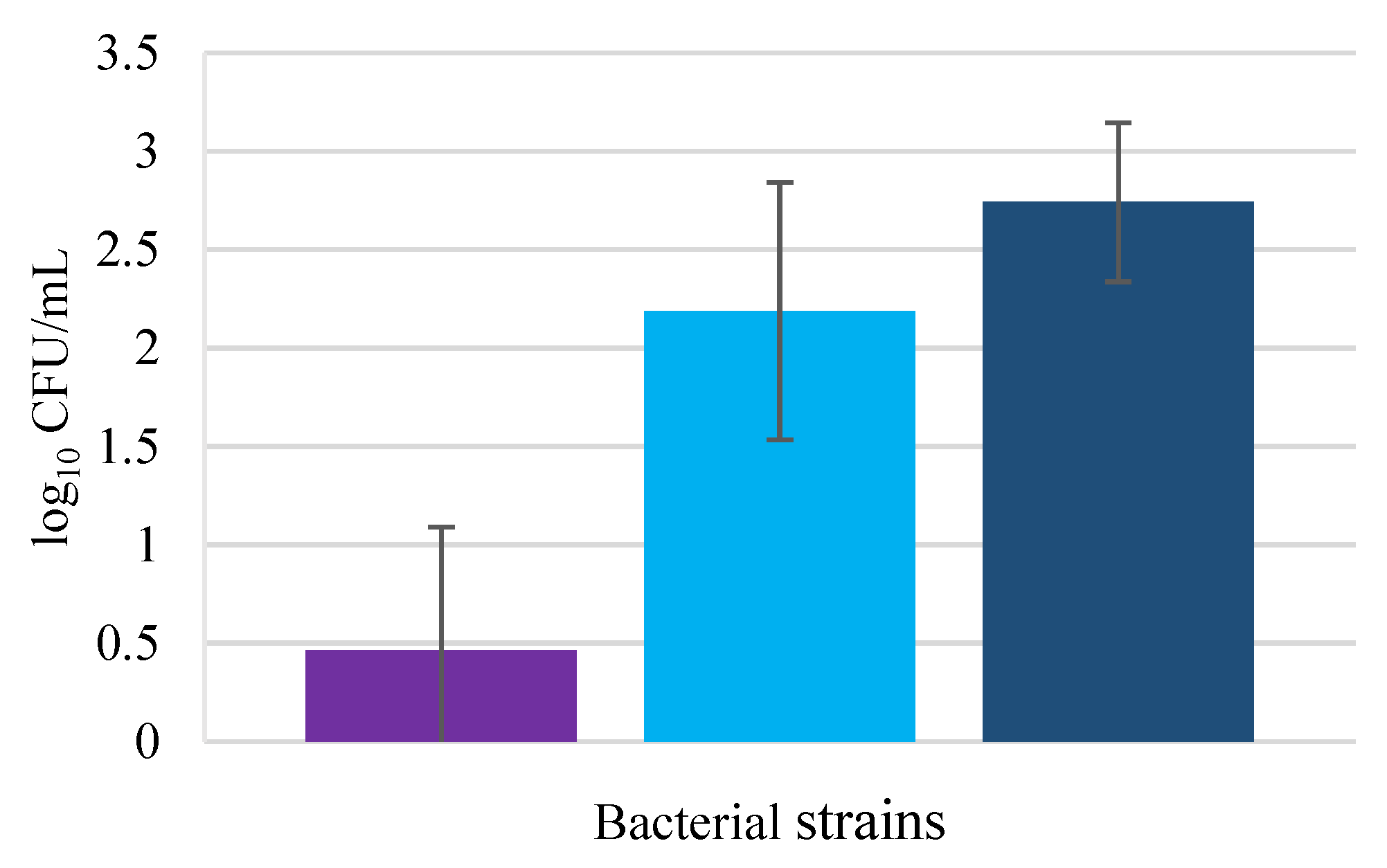
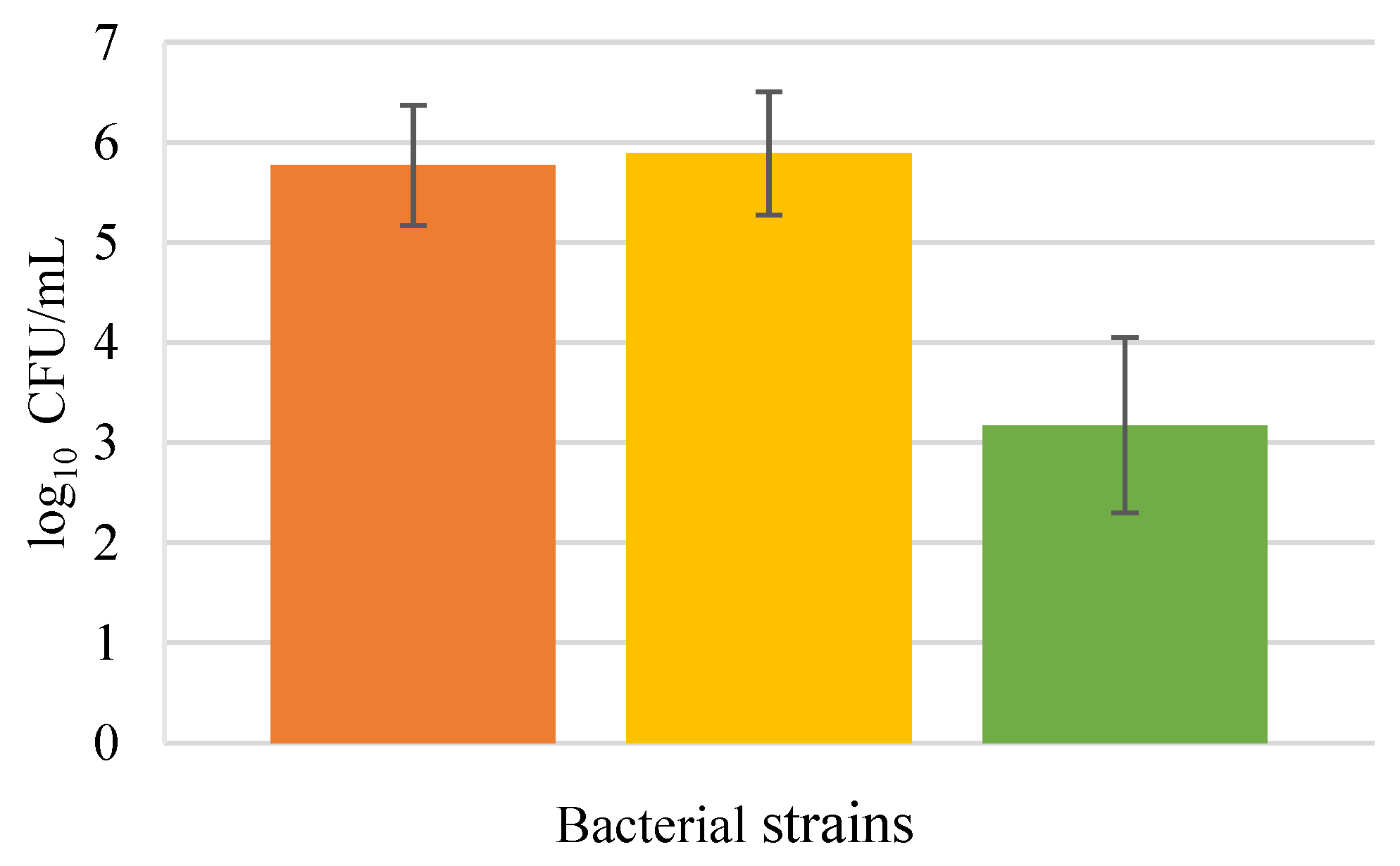
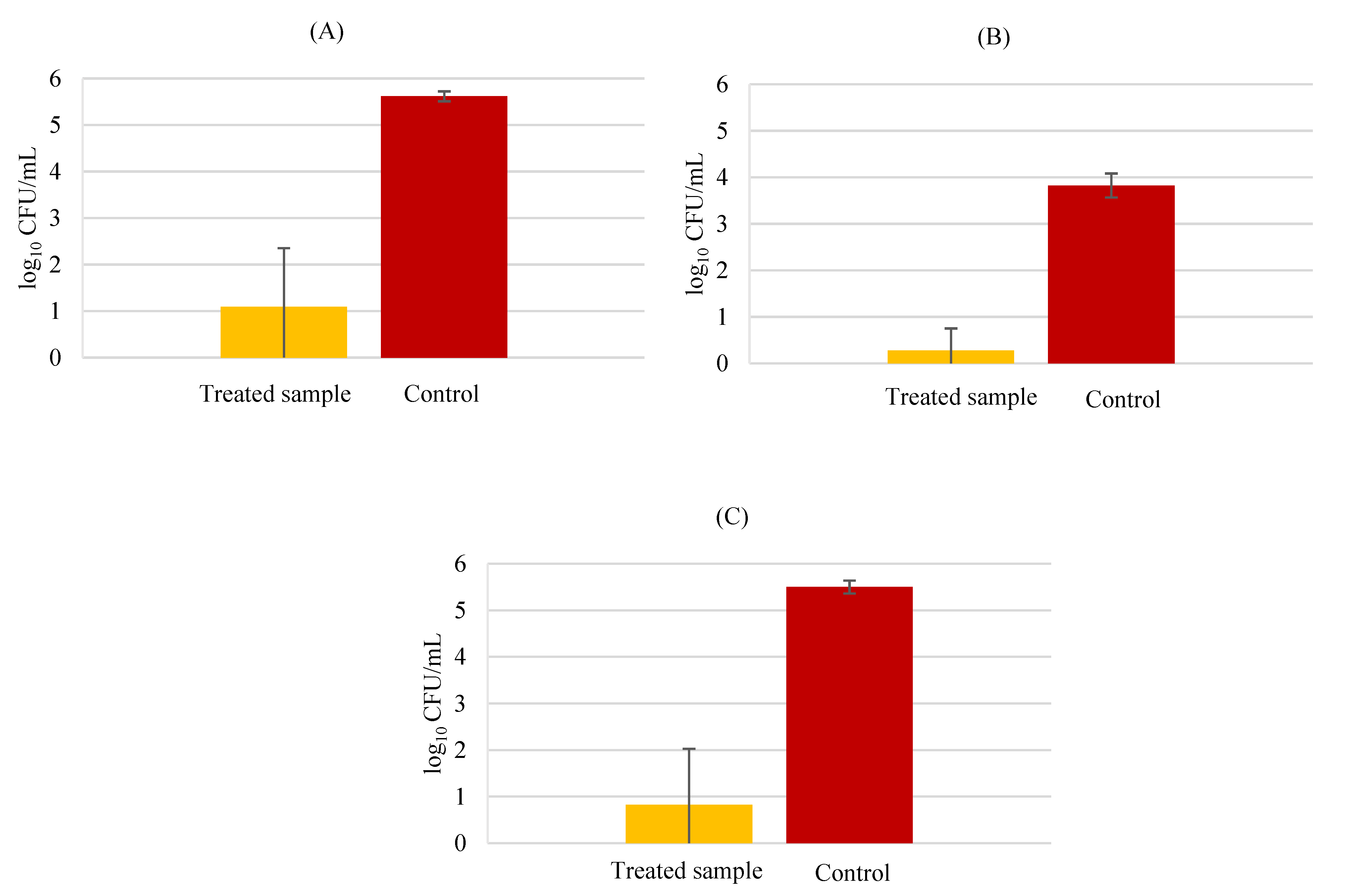
3.1. Invasion Assays
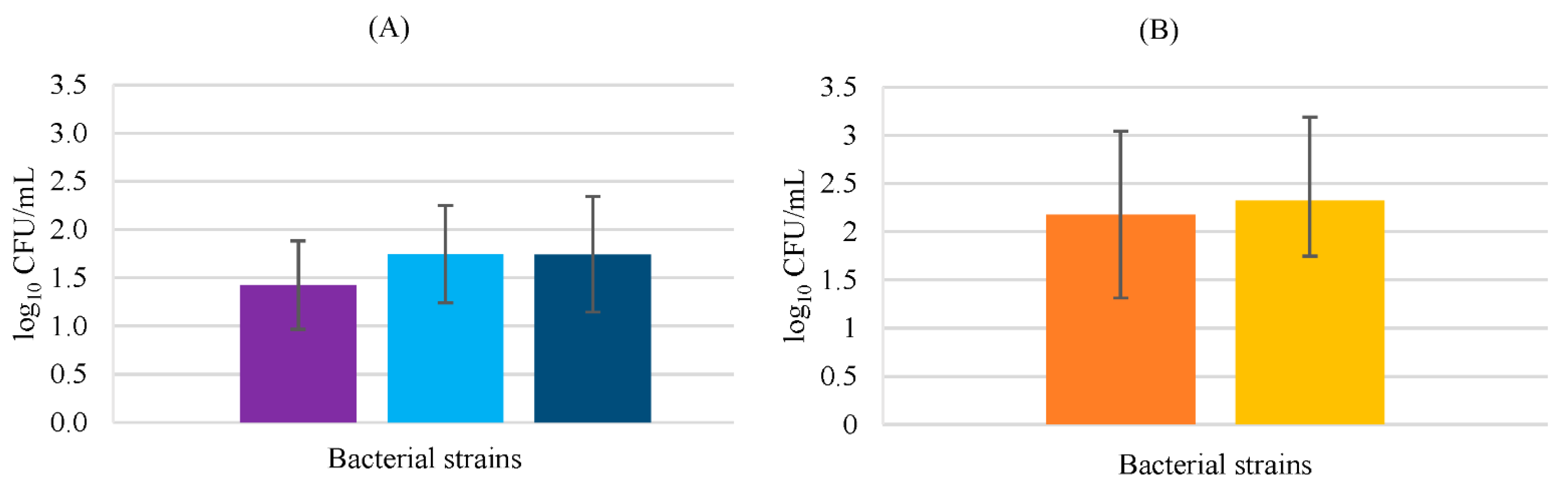
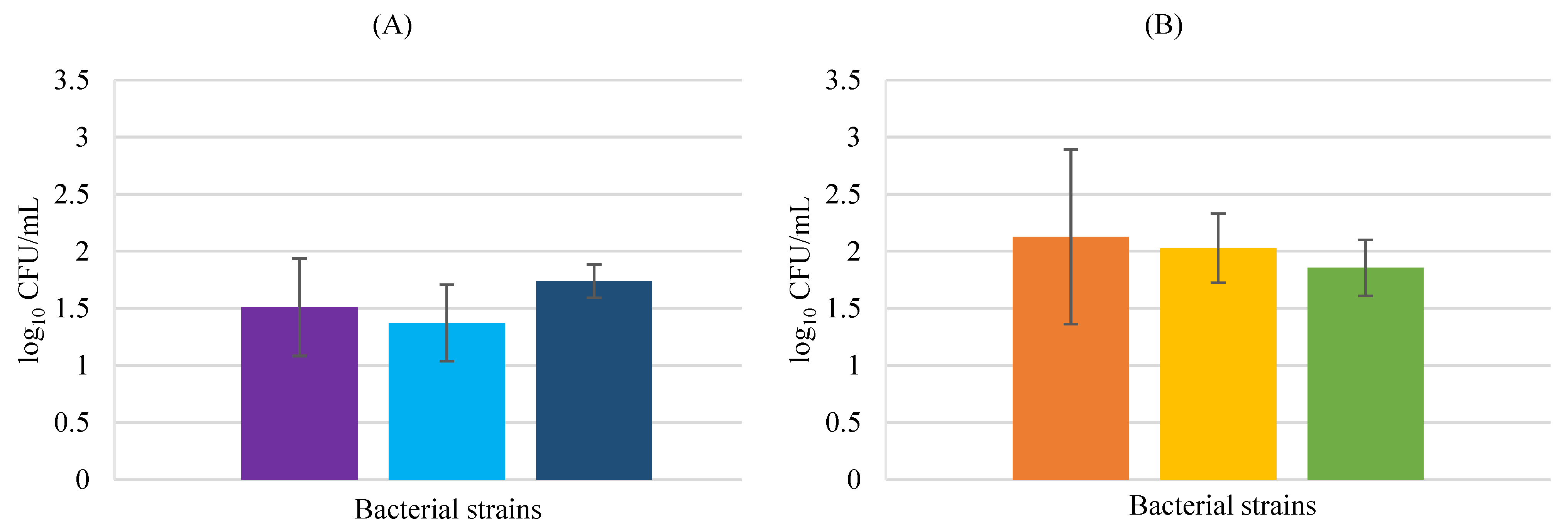
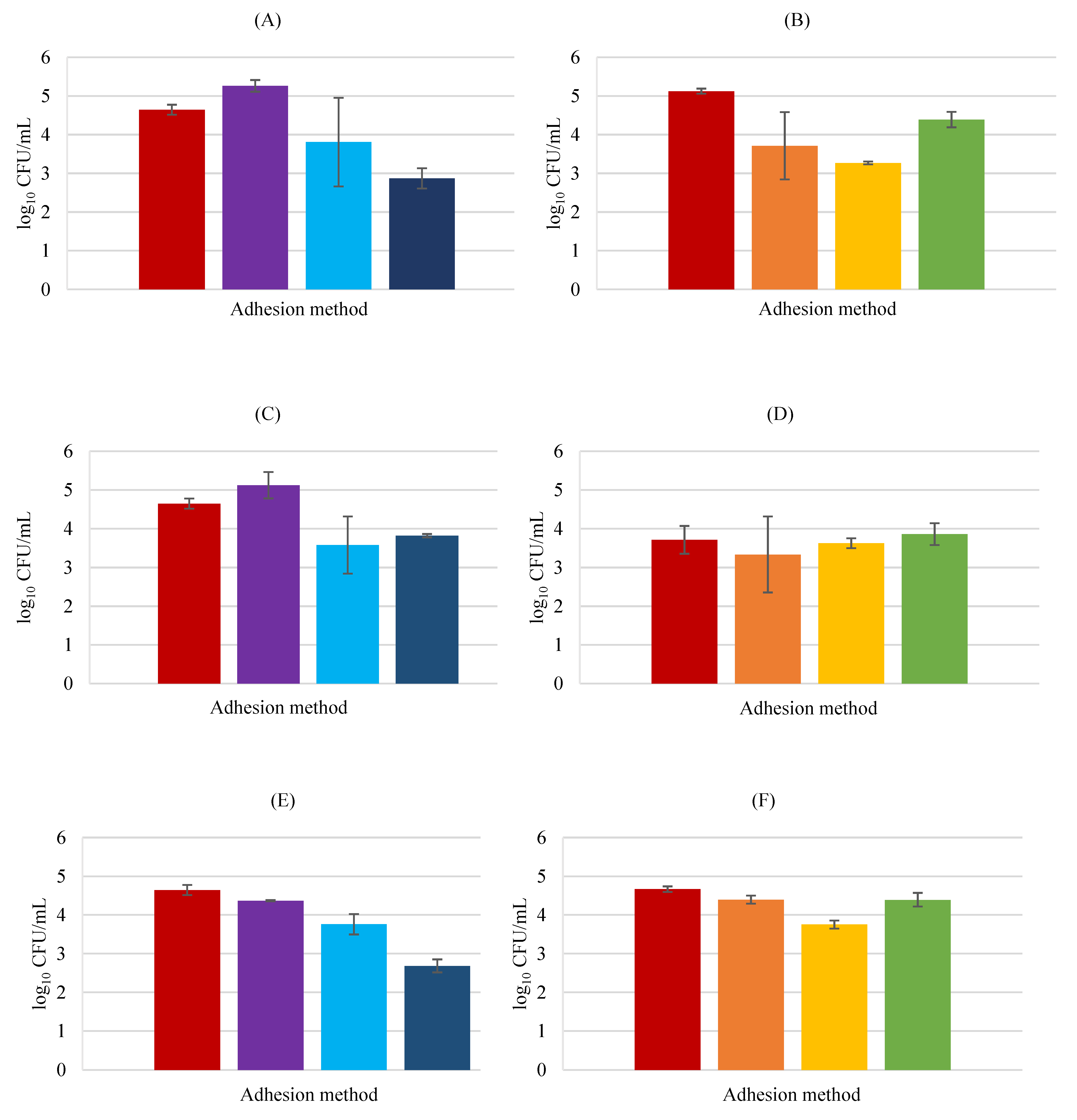
3.2. Adhesion Assays
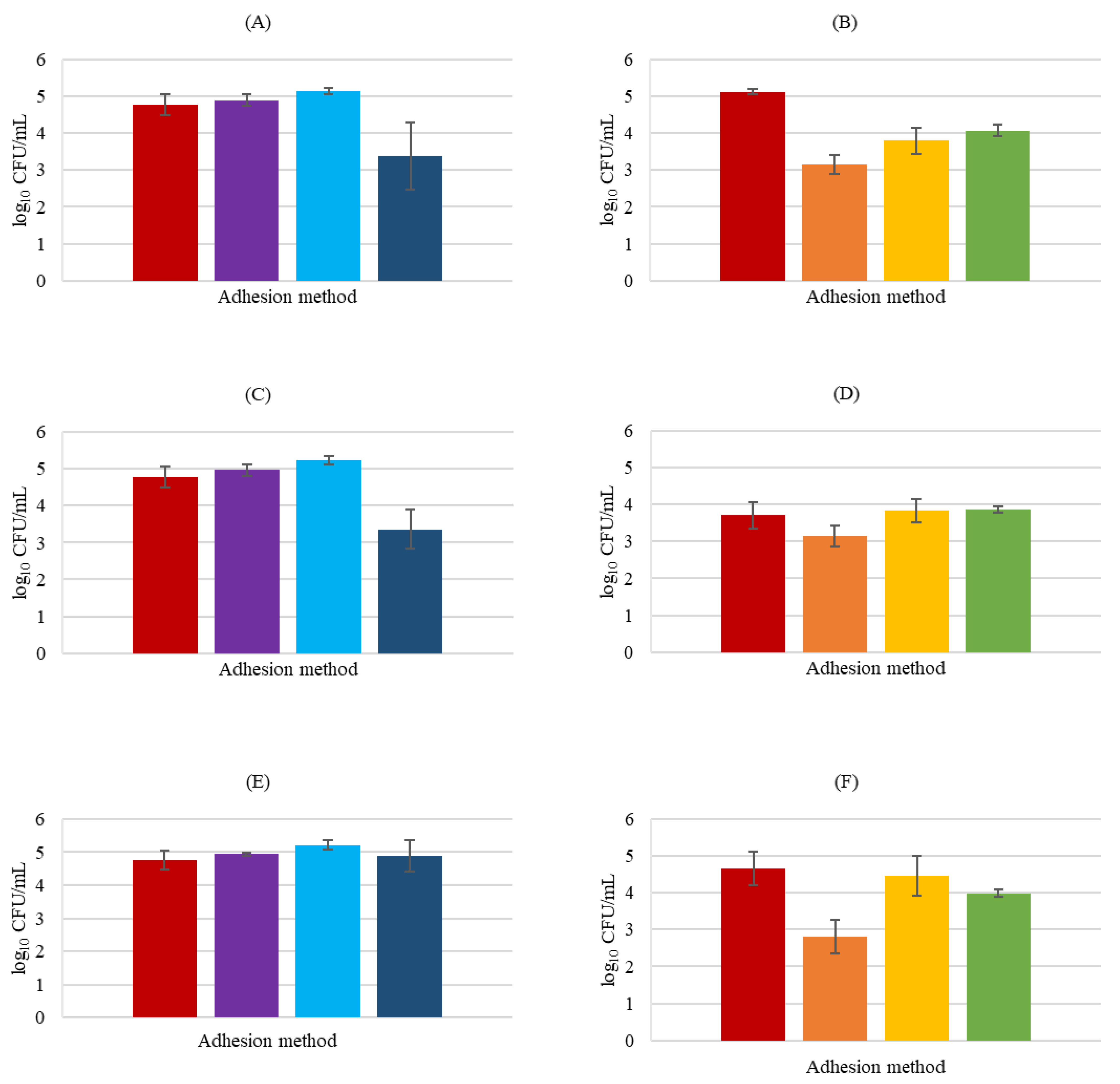
3.3. Adhesion Mechanisms
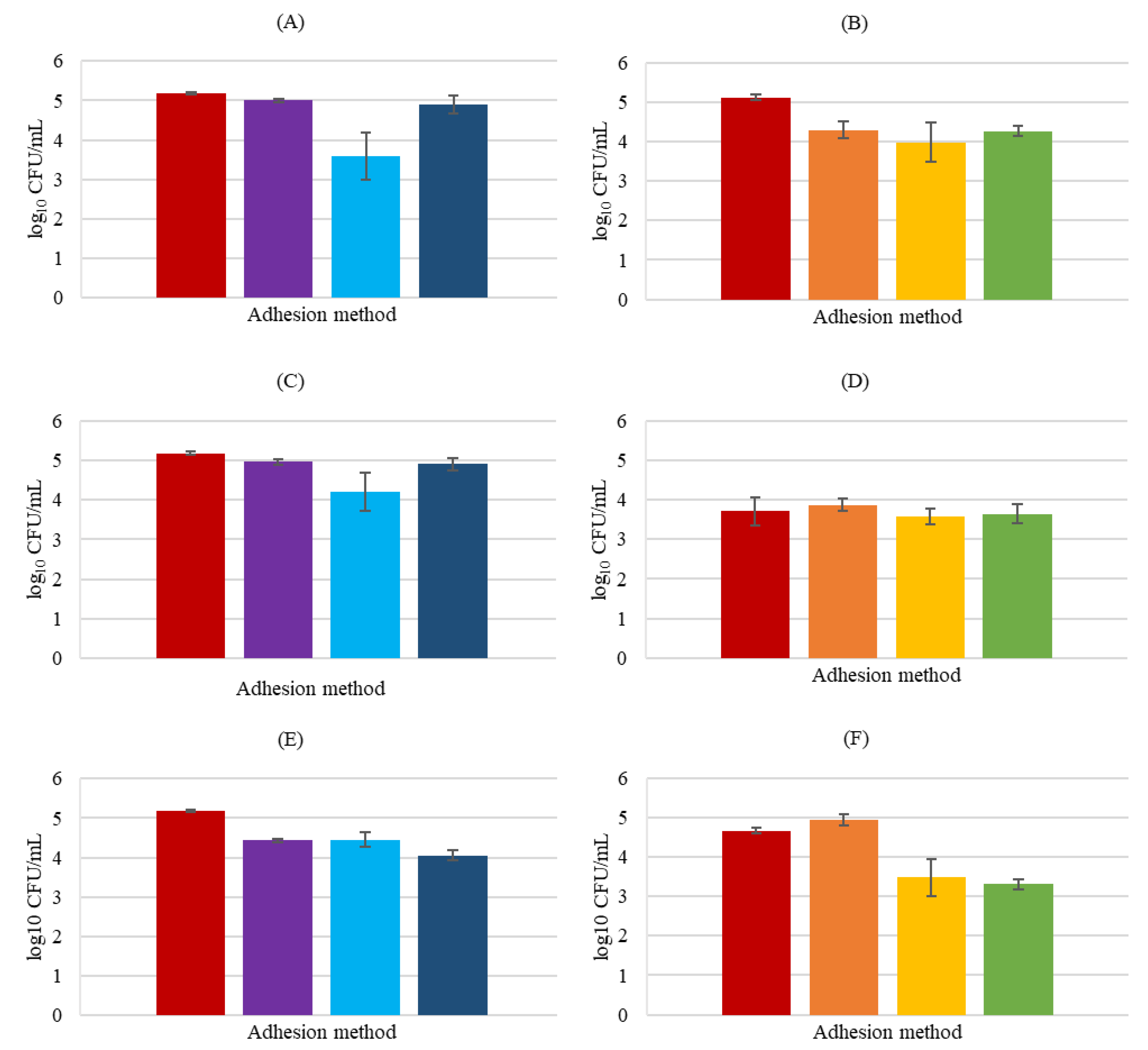
3.4. Carbohydrate Analysis
3.5. Protein Analysis
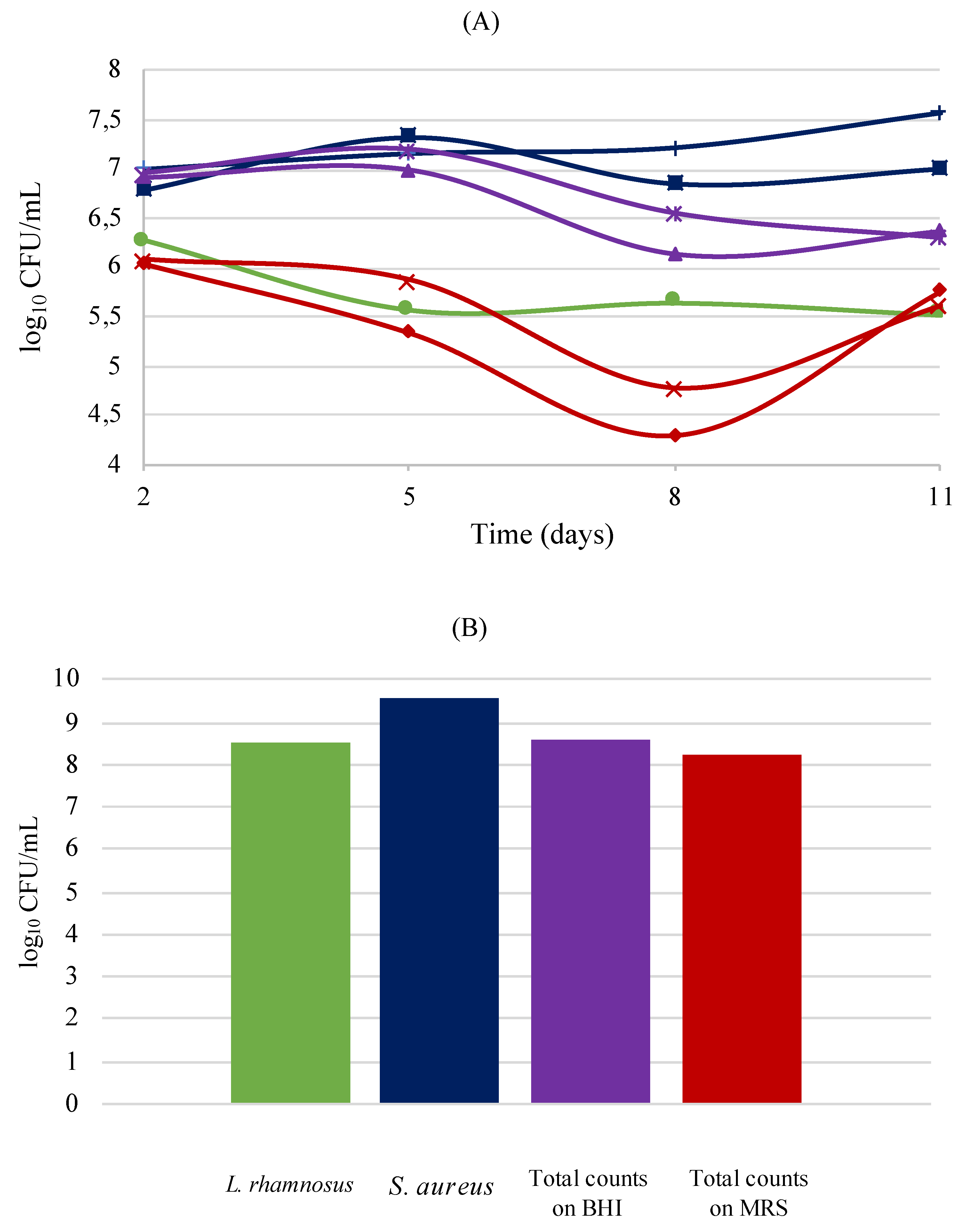
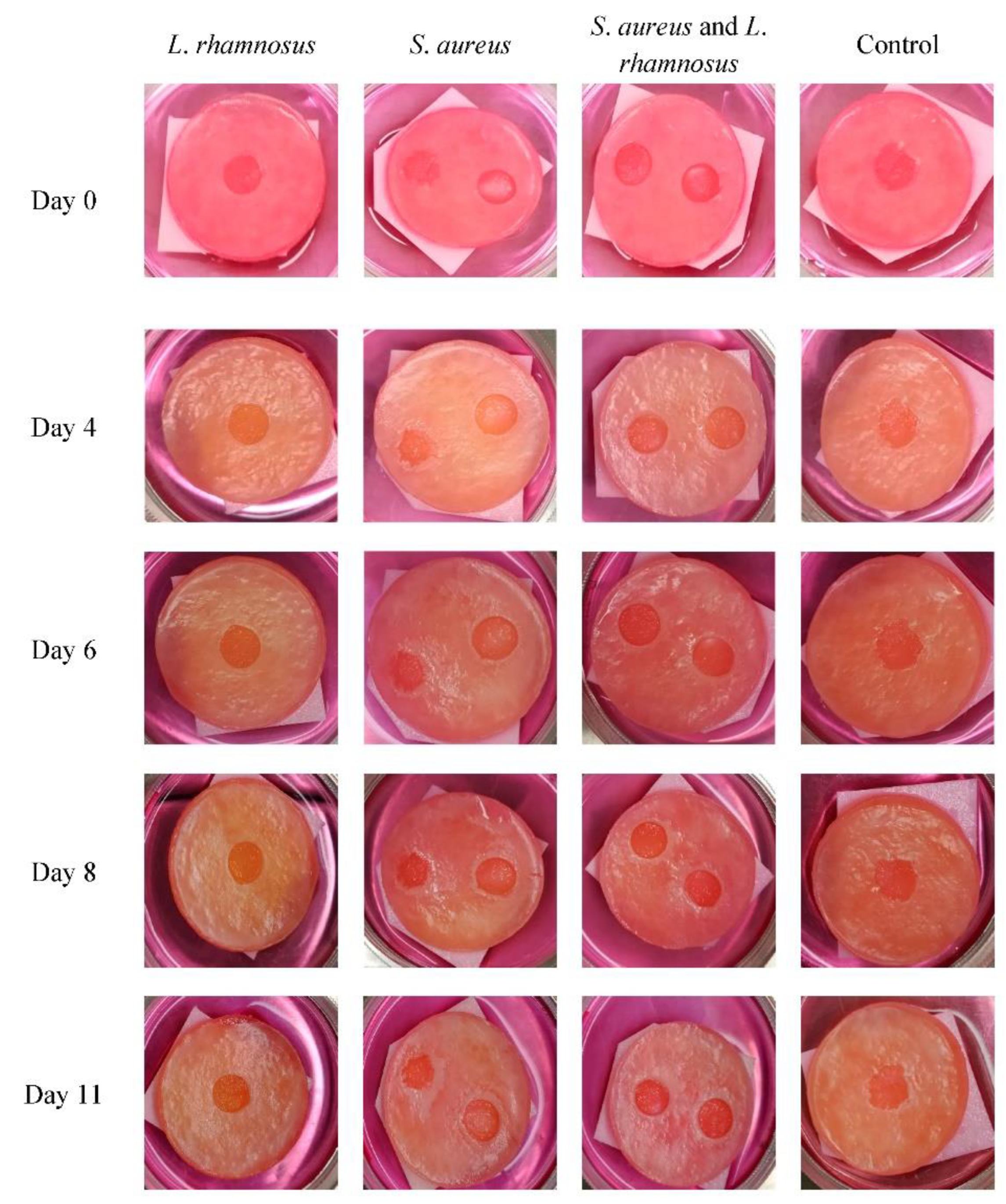
3.6. Human Skin Models
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fijan, S.; Frauwallner, A.; Langerholc, T.; Krebs, B.; Haar, J.; Heschl, A.; Mičetić Turk, D.; Rogelj, I. Efficacy of Using Probiotics with Antagonistic Activity against Pathogens of Wound Infections: An Integrative Review of Literature. BioMed Res. Int. 2019, 2019, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Egert, M.; Simmering, R. The microbiota of the human skin. Adv. Exp. Med. Biol. 2016, 902, 61–81. [Google Scholar] [CrossRef]
- Grogan, M.D.; Bartow-McKenney, C.; Flowers, L.; Knight, S.A.B.; Uberoi, A.; Grice, E.A. Research Techniques Made Simple: Profiling the Skin Microbiota. J. Investig. Dermatol. 2019, 139, 747–752.e1. [Google Scholar] [CrossRef] [PubMed]
- Nickoloff, B.J.; Turkat, L.A. Keratinocytes: Key Immunocytes of the Integument. Am. J. Pathol. 1993, 143, 325–331. [Google Scholar] [PubMed]
- Wilson, V.G. Growth and Differentiation of HaCaT Keratinocytes. Mehods Mol. Biol. 2013, 1341, 257–284. [Google Scholar] [CrossRef]
- Edwards, A.M.; Potter, U.; Meenan, N.A.; Potts, J.R.; Massey, R.C. Staphylococcus aureus keratinocyte invasion is dependent upon multiple high-affinity fibronectin-binding repeats within FnBPA. PLoS ONE 2011, 6, e18899. [Google Scholar] [CrossRef]
- Kaliyeva, S.S.; Lavrinenko, A.V.; Tishkambayev, Y.; Zhussupova, G.; Issabekova, A.; Begesheva, D.; Simokhina, N. Microbial Landscape and Antibiotic Susceptibility Dynamics of Skin and Soft Tissue Infections in Kazakhstan 2018. Antibiotics 2022, 11, 659. [Google Scholar] [CrossRef]
- Wu, W.; Jin, Y.; Bai, F. Chapter 41: Pseudomonas aeruginosa. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 2, pp. 753–767. [Google Scholar] [CrossRef]
- LaBauve, A.E.; Wargo, M.J. Growth and laboratory maintenance of Pseudomonas aeruginosa. Curr. Protoc. Microbiol. 2012, 6, Unit 6E.1. [Google Scholar] [CrossRef]
- Blicharz, L.; Rudnicka, L.; Samochocki, Z. Staphylococcus aureus: An underestimated factor in the pathogenesis of atopic dermatitis? Adv. Dermatol. Allergol. 2019, 36, 11–17. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Yoon, T.Y.; Jung, S.K.; Chang, S.H. Cellulitis due to Escherichia coli in three immunocompromised subjects. Br. J. Dermatol. 1998, 139, 885–888. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.W.; Alt-Holland, A.; Egles, C.; Garlick, J.A. Three-dimensional tissue models of normal and diseased skin. Curr. Protoc. Cell Biol. 2008, 19, Unit 19.9. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Tsuchiya, A.; Ogawa, M.; Ono, M.; Suda, N.; Sekimoto, K.; Takeo, M.; Tsuji, T. Tissue-scale tensional homeostasis in skin regulates structure and physiological function. Commun. Biol. Nat. Res. 2020, 3, 637. [Google Scholar] [CrossRef]
- Zhang, Z.; Michniak-Kohn, B.B. Tissue engineered human skin equivalents. Pharmaceutics 2012, 4, 26–41. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays 2016, 38, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.R.; Boland, C.R.; Patel, M.; Thrash, B.; Menter, A. Cutaneous manifestations of gastrointestinal disease: Part I. J. Am. Acad. Dermatol. 2013, 68, 189.e1-21. [Google Scholar] [CrossRef] [PubMed]
- Cinque, B.; la Torre, C.; Melchiorre, E.; Marchesani, G.; Zoccali, G.; Palumbo, P.; di Marzio, L.; Masci, A.; Mosca, L.; Mastromarino, P.; et al. Use of Probiotics for Dermal Applications. In Probiotics; Liong, M.T., Ed.; Springer: Berlin, Heidelberg, Germany, 2011; pp. 221–241. [Google Scholar]
- Prince, T. Evaluation of the Utility of Probiotics for the Prevention of Infections in a Model of the Skin. Ph.D. Thesis, University of Manchester, Manchester, UK, 2012. [Google Scholar]
- Gläser, R.; Harder, J.; Lange, H.; Bartels, J.; Christophers, E.; Schröder, J.M. Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 2005, 6, 57–64. [Google Scholar] [CrossRef]
- Rangaraj, A.; Ye, L.; Sanders, A.J.; Price, P.E.; Harding, K.G.; Jiang, W.G. Molecular and cellular impact of Psoriasin (S100A7) on the healing of human wounds. Exp. Ther. Med. 2017, 13, 2151–2160. [Google Scholar] [CrossRef]
- Prasad, J.; McJarrow, P.; Gopal, P. Heat and osmotic stress responses of probiotic Lactobacillus rhamnosus HN001 (DR20) in relation to viability after drying. Appl. Environ. Microbiol. 2003, 69, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Hosseinidoust, Z.; van de Ven, T.G.M.; Tufenkji, N. Evolution of Pseudomonas aeruginosa Virulence as a Result of Phage Predation. Appl. Environ. Microbiol. 2013, 79, 6110–6116. [Google Scholar] [CrossRef]
- Prasad, A.S.B. Pseudomonas aeruginosa virulence proteins pseudolysin and protease IV impede cutaneous wound healing. Lab. Investig. 2020, 100, 1532–1550. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Morello, E.; Garnier, J.; Barrault, C.; Garnier, M.; Burucoa, C.; Lecron, J.C.; Si-Tahar, M.; Bernard, F.X.; Bodet, C. Pseudomonas aeruginosa flagellum is critical for invasion, cutaneous persistence and induction of inflammatory response of skin epidermis. Virulence 2018, 9, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Mempel, M.; Schnopp, C.; Hojka, M.; Fesq, H.; Weidinger, S.; Schaller, M.; Korting, H.C.; Ring, J.; Abeck, D. Invasion of human keratinocytes by Staphylococcus aureus and intracellular bacterial persistence represent haemolysin-independent virulence mechanisms that are followed by features of necrotic and apoptotic keratinocyte cell death. Br. J. Dermatol. 2002, 146, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Prince, T.; McBain, A.J.; O’Neill, C.A. Lactobacillus reuteri protects epidermal keratinocytes from Staphylococcus aureus-induced cell death by competitive exclusion. Appl. Environ. Microbiol. 2012, 78, 5119–5126. [Google Scholar] [CrossRef]
- Mohammedsaeed, W.; McBain, A.J.; Cruickshank, S.M.; O’Neill, C.A. Lactobacillus rhamnosus GG inhibits the toxic effects of Staphylococcus aureus on epidermal keratinocytes. Appl. Environ. Microbiol. 2014, 80, 5773–5781. [Google Scholar] [CrossRef]
- Abedi, D.; Feizizadeh, S.; Akbari, V.; Jafarian-Dehkordi, A. In vitro anti-bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp. bulgaricus on Escherichia coli. Res. Pharm. Sci. 2013, 8, 260–268. [Google Scholar]
- Al-Malkey, M.K.; Ismeeal, M.C.; Abo Al-Hur, F.J.; Mohammed, S.W.; Nayyef, H.J. Antimicrobial effect of probiotic Lactobacillus spp. on Pseudomonas aeruginosa. J. Contemp. Med. Sci. 2017, 3, 218–223. [Google Scholar] [CrossRef]
- Lopes, E.G.; Moreira, D.A.; Gullón, P.; Gullón, B.; Cardelle-Cobas, A.; Tavaria, F.K. Topical application of probiotics in skin: Adhesion, antimicrobial and antibiofilm in vitro assays. J. Appl. Microbiol. 2017, 122, 450–461. [Google Scholar] [CrossRef]
- Coppola, R.; Succi, M.; Tremonte, P.; Reale, A.; Salzano, G. Antibiotic susceptibility of Lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. Le Lait 2005, 85, 193–204. [Google Scholar] [CrossRef]
- Hor, Y.Y.; Liong, M.T. Use of extracellular extracts of lactic acid bacteria and bifidobacteria for the inhibition of dermatological pathogen Staphylococcus aureus. Dermatol. Sin. 2014, 32, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Saenger, W. Proteinase K. In Handbook of Proteolytic Enzymes; Rawlings, N.D., Salvesen, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 3240–3242. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, J.; Song, J.; Yang, J.; Xu, T.; Pan, C.; Zhang, L. One-step synthesis of an antibacterial and pro-healing wound dressing that can treat wound infections. J. Mat. Chem. B 2017, 5, 8451–8458. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, J.; Douglas, I.; Rimmer, S.; Swanson, L.; MacNeil, S. Development of three-dimensional tissue-engineered models of bacterial infected human skin wounds. Tissue Eng. Part. C Methods 2009, 15, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Beldon, P. Basic science of wound healing. Surgery 2010, 28, 409–412. [Google Scholar] [CrossRef]
- Jung, Y.O.; Jeong, H.; Cho, Y.; Lee, E.O.; Jang, H.W.; Kim, J.; Nam, K.; Lim, K.M. Lysates of a probiotic, Lactobacillus rhamnosus, can improve skin barrier function in a reconstructed human epidermis model. Int. J. Mol. Sci. 2019, 20, 4289. [Google Scholar] [CrossRef]
- Ortines, R.V.; Liu, H.; Cheng, L.I.; Cohen, T.S.; Lawlor, H.; Gami, A.; Wang, Y.; Dillen, C.A.; Archer, N.K.; Miller, R.J.; et al. Neutralizing alpha-toxin accelerates healing of Staphylococcus aureus-infected wounds in nondiabetic and diabetic mice. Antimicrob. Agents Chemother. 2018, 62, e02288-17. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lizardo, M.; Magalhães, R.M.; Tavaria, F.K. Probiotic Adhesion to Skin Keratinocytes and Underlying Mechanisms. Biology 2022, 11, 1372. https://doi.org/10.3390/biology11091372
Lizardo M, Magalhães RM, Tavaria FK. Probiotic Adhesion to Skin Keratinocytes and Underlying Mechanisms. Biology. 2022; 11(9):1372. https://doi.org/10.3390/biology11091372
Chicago/Turabian StyleLizardo, Mariana, Rui Miguel Magalhães, and Freni Kekhasharú Tavaria. 2022. "Probiotic Adhesion to Skin Keratinocytes and Underlying Mechanisms" Biology 11, no. 9: 1372. https://doi.org/10.3390/biology11091372