
Liver Graft MicroRNAs Expression in Different Etiology of Acute Jaundice after Living Donor Liver Transplantation

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. Patient Characteristics
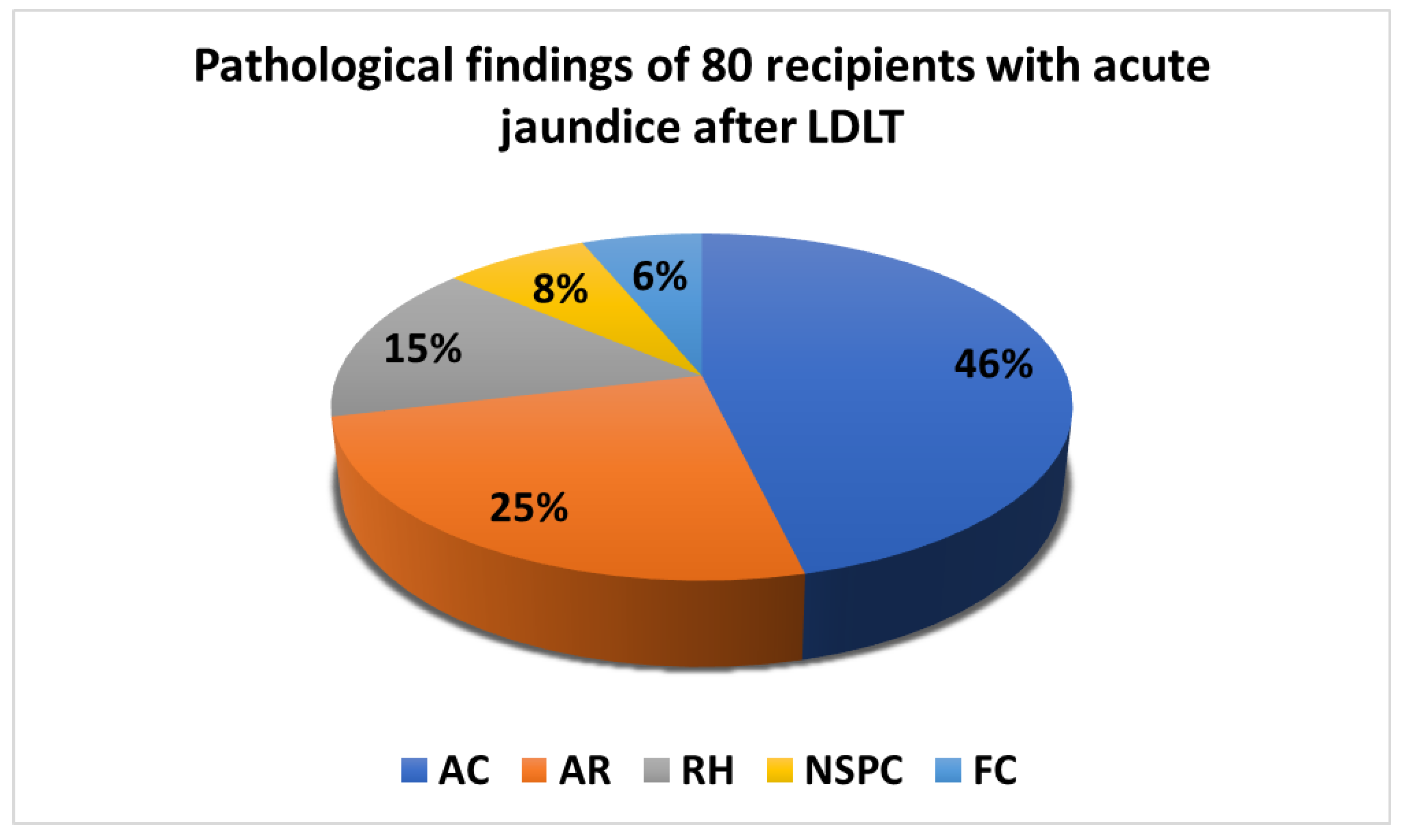
2.2. Clinical Biochemistry Data and Liver Graft Pathologies
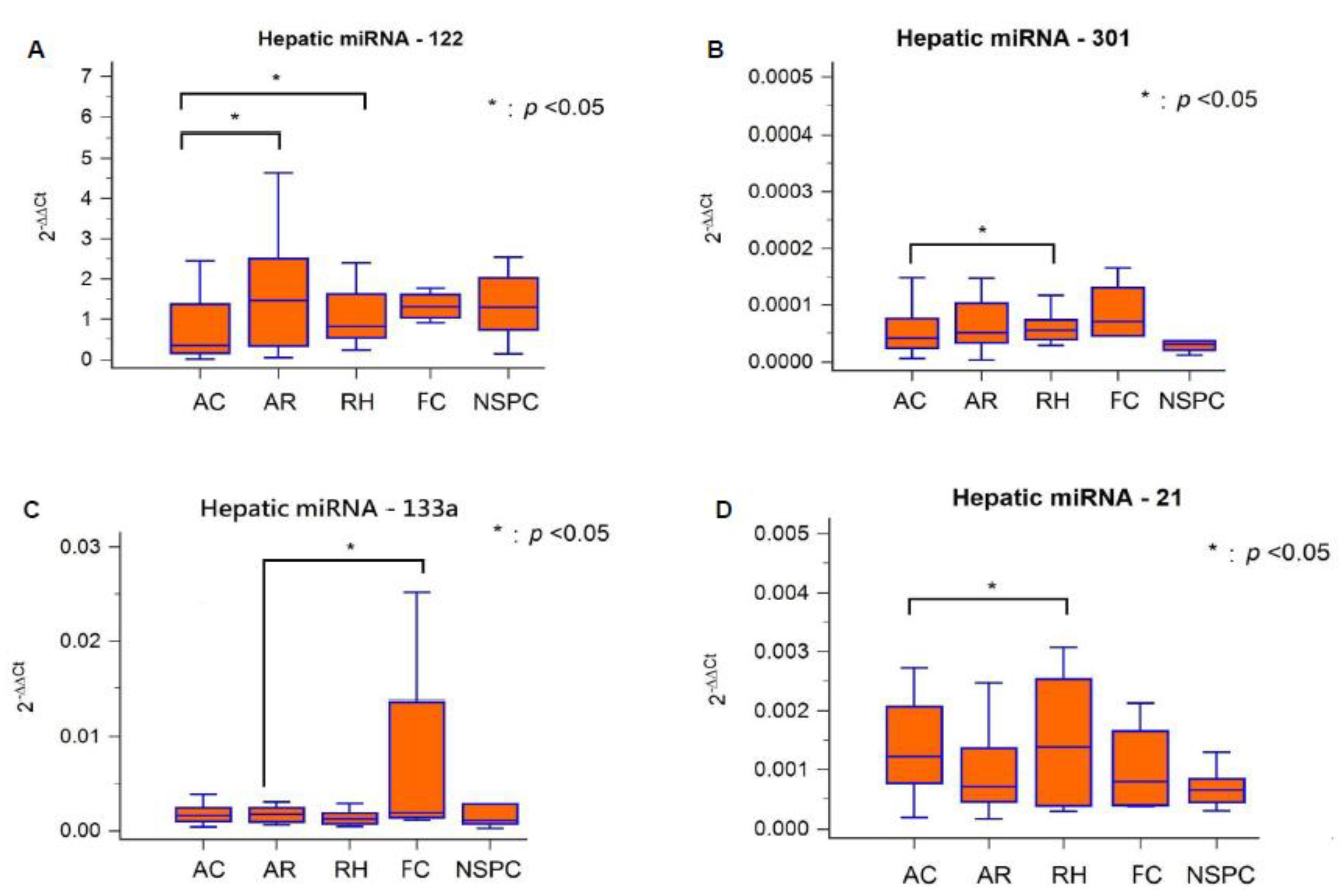
2.3. Hepatic miRNAs Expression Patterns in Different Graft Pathologies
3. Discussion
4. Methods
4.1. Study Population and Design
4.2. Real-Time Quantitative Reverse Transcription—Polymerase Chain Reaction for Hepatic miRNA Identification in Liver Graft
4.3. Ethical Statement
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Millson, C.; Considine, A.; Cramp, M.E.; Holt, A.; Hubscher, S.; Hutchinson, J.; Jones, K.; Leithead, J.; Masson, S.; Menon, K.; et al. Adult liver transplantation: UK clinical guideline-part 2: Surgery and post-operation. Frontline Gastroenterol. 2020, 11, 385–396. [Google Scholar] [CrossRef]
- Sood, S.; Testro, A.G. Immune monitoring post liver transplant. World J. Transplant. 2014, 4, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Chiu, K.W.; Eng, H.L.; Cheng, Y.F.; Jawan, B.; Chen, C.L. Application of percutaneous liver biopsy in living donor liver transplantation. Int. J. Clin. Exp. Med. 2018, 11, 10372–10379. [Google Scholar]
- Israeli, M.; Klein, T.; Brandhorst, G.; Oellerich, M. Confronting the challenge: Individualized immune monitoring after organ transplantation using the cellular immune function assay. Clin. Chim. Acta 2012, 413, 1374–1378. [Google Scholar] [CrossRef] [PubMed]
- Kohut, T.J.; Barandiaran, J.F.; Keating, B.J. Genomics and Liver Transplantation: Genomic Biomarkers for the Diagnosis of Acute Cellular Rejection. Liver Transplant. 2020, 26, 1337–1350. [Google Scholar] [CrossRef]
- Gui, J.; Tian, Y.; Wen, X.; Zhang, W.; Zhang, P.; Gao, J.; Run, W.; Tian, L.; Jia, X.; Gao, Y. Serum microRNA characterization identifies miR-885-5p as a potential marker for detecting liver pathologies. Clin. Sci. 2011, 120, 183–193. [Google Scholar] [CrossRef]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA profiling: Approaches and considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef]
- Ekine-Afolabi, B.A.; Njan, A.A.; Rotimi, S.O.; Ri, A.; Elbehi, A.M.; Cash, E.; Adeyeye, A. The Impact of Diet on the Involvement of Non-Coding RNAs, Extracellular Vesicles, and Gut Microbiome-Virome in Colorectal Cancer Initiation and Progression. Front. Oncol. 2020, 10, 583372. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Finch, M.L.; Marquardt, J.U.; Yeoh, G.C.; Callus, B.A. Regulation of microRNAs and their role in liver development, regeneration and disease. Int. J. Biochem. Cell Biol. 2014, 54, 288–303. [Google Scholar] [CrossRef]
- Zhou, M.; Hara, H.; Dai, Y.; Mou, L.; Cooper, D.K.; Wu, C.; Cai, Z. Circulating Organ-Specific MicroRNAs Serve as Biomarkers in Organ-Specific Diseases: Implications for Organ Allo- and Xeno-Transplantation. Int. J. Mol. Sci. 2016, 17, 1232. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, T.; Akat, K.M.; Yang, H.; Schwartz, J.E.; Li, C.; Bang, H.; Ben-Dov, I.Z.; Lee, J.R.; Ikle, D.; Demetris, A.J.; et al. Serum MicroRNA Transcriptomics and Acute Rejection or Recurrent Hepatitis C Virus in Human Liver Allograft Recipients: A Pilot Study. Transplantation 2021, 106, 806–820. [Google Scholar] [CrossRef] [PubMed]
- Wanga, C.C.; Huangc, K.T.; Chenc, K.D.; Hsuc, L.W.; Lina, C.C.; Chiub, K.W. Hepatic miR-122 expression correlated with IL-28B genetic polymorphisms in hepatocellular carcinoma patients with living donor liver transplantation. ScienceAsia 2021, 47, 578–584. [Google Scholar] [CrossRef]
- Wang, X.W.; Heegaard, N.H.; Orum, H. MicroRNAs in liver disease. Gastroenterology 2012, 142, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Chen, I.H.; Goto, S.; Lai, C.Y.; Tseng, H.P.; Hsu, L.W.; Chiu, K.W.; Lin, C.C.; Wang, C.C.; Cheng, Y.F. Hepatic miR-301a as a Liver Transplant Rejection Biomarker? And Its Role for Interleukin-6 Production in Hepatocytes. OMICS 2017, 21, 55–66. [Google Scholar] [CrossRef]
- Bessho, K.; Shanmukhappa, K.; Sheridan, R.; Shivakumar, P.; Mourya, R.; Walters, S.; Kaimal, V.; Dilbone, E.; Jegga, A.G.; Bezerra, J.A. Integrative genomics identifies candidate microRNAs for pathogenesis of experimental biliary atresia. BMC Syst. Biol. 2013, 7, 104. [Google Scholar] [CrossRef]
- Olaizola, P.; Lee-Law, P.Y.; Arbelaiz, A.; Lapitz, A.; Perugorria, M.J.; Bujanda, L.; Banales, J.M. MicroRNAs and extracellular vesicles in cholangiopathies. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1293–1307. [Google Scholar] [CrossRef]
- Kennedy, L.L.; Meng, F.; Venter, J.K.; Zhou, T.; Karstens, W.A.; Hargrove, L.A.; Wu, N.; Kyritsi, K.; Greene, J.; Invernizzi, P.; et al. Knockout of microRNA-21 reduces biliary hyperplasia and liver fibrosis in cholestatic bile duct ligated mice. Lab. Investig. 2016, 96, 1256–1267. [Google Scholar] [CrossRef]
- Hamdorf, M.; Kawakita, S.; Everly, M. The Potential of MicroRNAs as Novel Biomarkers for Transplant Rejection. J. Immunol. Res. 2017, 2017, 4072364. [Google Scholar] [CrossRef]
- Loureiro, D.; Tout, I.; Narguet, S.; Benazzouz, S.M.; Mansouri, A.; Asselah, T. miRNAs as Potential Biomarkers for Viral Hepatitis B and C. Viruses 2020, 12, 1440. [Google Scholar] [CrossRef]
- Di Stefano, A.B.; Pappalardo, M.; Moschella, F.; Cordova, A.; Toia, F. MicroRNAs in solid organ and vascularized composite allotransplantation: Potential biomarkers for diagnosis and therapeutic use. Transplant. Rev. 2020, 34, 100566. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. miRNA-based biomarkers, therapies, and resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, H.; Zhao, M.; Chang, C.; Lu, Q. Clinical significance of miRNAs in autoimmunity. J. Autoimmun. 2020, 109, 102438. [Google Scholar] [CrossRef]
- Kaul, V.; Krams, S. MicroRNAs as master regulators of immune responses in transplant recipients. Curr. Opin. Organ Transplant. 2015, 20, 29–36. [Google Scholar] [CrossRef]
- Baker, J.R.; Vuppusetty, C.; Colley, T.; Papaioannou, A.I.; Fenwick, P.; Donnelly, L.; Ito, K.; Barnes, P.J. Oxidative stress dependent microRNA-34a activation via PI3Kα reduces the expression of sirtuin-1 and sirtuin-6 in epithelial cells. Sci. Rep. 2016, 6, 35871. [Google Scholar] [CrossRef]
- Tavabie, O.D.; Karvellas, C.J.; Salehi, S.; Speiser, J.L.; Rose, C.F.; Menon, K.; Prachalias, A.; Heneghan, M.A.; Agarwal, K.; Lee, W.M.; et al. A novel microRNA-based prognostic model outperforms standard prognostic models in patients with acetaminophen-induced acute liver failure. J. Hepatol. 2021, 75, 424–434. [Google Scholar] [CrossRef]
- López Panqueva, R.D.P. Algoritmos útiles para el diagnóstico histopatológico de la enfermedad hepática con base en los patrones de daño hepático. Rev. Colomb. Gastroenterol. 2016, 31, 443–457. [Google Scholar] [CrossRef]
- Fiel, M.I. Pathology of chronic hepatitis B and chronic hepatitis C. Clin. Liver Dis. 2010, 14, 555–575. [Google Scholar] [CrossRef]
- Brown, G.T.; Kleiner, D.E. Histopathology of nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Metabolism 2016, 65, 1080–1086. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Category | Patients (n = 80) |
|---|---|
| Age, mean ± SD (range) (years) | 52.56 ± 13.26 (20–66) |
| Gender (male/female), n (%) | 58 (72.5%)/22 (27.5%) |
| Primary liver disease, n (%) | |
| HBV | 28 (35%) |
| HCV | 22 (27.5%) |
| HBV + HCV | 3 (3.75%) |
| Alcohol | 14 (17.5%) |
| Other (Cholangiocarcinoma, Biliary atresia, Primary biliary cirrhosis, Cryptogenic cirrhosis) | 13 (16.25%) |
| Laboratory data | |
| AFP (ng/dL), mean ± SD | 7.29 ± 2.33 |
| Albumin (g/dL) | 4.11 ± 0.54 |
| ALT (U/L) | 165.19 ± 133.23 |
| AST (U/L) | 232.70 ± 194.03 |
| Total bilirubin (mg/dL) | 3.25 ± 6.81 |
| Direct bilirubin (mg/dL) | 1.91 ± 4.64 |
| ALK-P (U/L) | 282.08 ± 335.13 |
| r-GT (U/L) | 183.17 ± 180.17 |
| WBC (103/μL) | 4.48 ± 2.26 |
| Hemoglobin (g/dL) | 11.80 ± 3.39 |
| Segment % | 62.01 ± 10.87 |
| Lymphocyte % | 26.24 ± 10.58 |
| N/L ratio | 3.39 ± 2.96 |
| Category | AC, n = 37 (46%) | AR, n = 20 (25%) | RH, n = 12 (15%) | FC, n = 5 (6%) | NSPC, n = 6 (8%) | p Value |
|---|---|---|---|---|---|---|
| Albumin | 4.16 ± 0.53 | 3.99 ± 0.63 | 4.03 ± 0.52 | 4.42 ± 0.45 | 4.17 ± 0.39 | 0.140 |
| ALT | 152.68 ± 96.75 | 212.40 ±174.52 | 152.33 ±187.89 | 142.00 ± 38.53 | 130.00 ± 77.21 | 0.051 |
| AST | 217.51 ± 131.87 | 269.30 ± 183.06 | 250.50 ± 373.12 | 167.00 ± 57.29 | 223.50 ±145.24 | 0.112 |
| Total Bilirubin | 5.71 ± 13.69 | 5.32 ± 10.40 | 0.80 ± 0.47 | 0.90 ± 0.41 | 0.87 ± 0.37 | 0.456 |
| Direct Bilirubin | 2.20 ± 4.22 | 10.36 ± 31.55 | 5.45 ± 17.81 | 0.24 ± 0.22 | 0.27 ± 0.12 | 0.062 |
| ALK-P | 375.51 ± 393.50 | 312.80 ± 339.23 | 107.50 ± 55.29 | 99.20 ±44.68 | 105.00 ± 38.16 | 0.275 |
| γ-GT | 247.81 ± 211.48 | 165.35 ± 134.16 | 89.03 ± 104.91 | 146.00 ±170.66 | 63.18 ± 53.23 | 0.060 |
| N/L ratio | 3.61 ± 3.76 | 3.01 ± 2.48 | 4.46 ± 2.44 | 1.77 ± 0.77 | 2.45 ± 0.59 | 0.227 |
| Category | miRNA-122 | miRNA-301 | miRNA-133a | miRNA-21 |
|---|---|---|---|---|
| AC, n = 37 (46%) | 1.06 ± 1.39 a,b | 0.000068 ± 0.000084 c | 0.0086 ± 0.0256 | 0.00139 ± 0.00101 e |
| AR, n = 20 (25%) | 3.20 ± 7.27 a’ | 0.000080 ± 0.000079 | 0.0036 ± 0.0054 d | 0.00137 ± 0.00151 |
| RH, n = 12 (15%) | 4.05 ± 10.37 b’ | 0.000189 ± 0.000430 c’ | 0.0159 ± 0.0465 | 0.00319 ± 0.00608 e’ |
| FC, n = 5 (6%) | 1.09 ± 0.60 | 0.000071 ± 0.000061 | 0.0402 ± 0.0738 d’ | 0.00087 ± 0.00079 |
| NSPC, n = 6 (8%) | 1.34 ± 0.78 | 0.000050 ± 0.000061 | 0.0039 ± 0.0067 | 0.00070 ± 0.00035 |
| Category | miRNA-122 | miRNA-301 | miRNA-133a | miRNA-21 |
|---|---|---|---|---|
| Acute cholangitis (AC) | ▽ | ▽ | ▽ | |
| Acute rejection (AR) | ▲ | ▽ | ||
| Recurrent hepatitis (RH) | ▲ | ▲ | ▲ | |
| Fatty change (FC) | ▲ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-H.; Wu, K.-T.; Wang, C.-C.; Huang, K.-T.; Chen, K.-D.; Hsu, L.-W.; Eng, H.-L.; Chiu, K.-W. Liver Graft MicroRNAs Expression in Different Etiology of Acute Jaundice after Living Donor Liver Transplantation. Biology 2022, 11, 1228. https://doi.org/10.3390/biology11081228
Lin S-H, Wu K-T, Wang C-C, Huang K-T, Chen K-D, Hsu L-W, Eng H-L, Chiu K-W. Liver Graft MicroRNAs Expression in Different Etiology of Acute Jaundice after Living Donor Liver Transplantation. Biology. 2022; 11(8):1228. https://doi.org/10.3390/biology11081228
Chicago/Turabian StyleLin, Shu-Hsien, Kun-Ta Wu, Chih-Chi Wang, Kuang-Tzu Huang, Kuang-Den Chen, Li-Wen Hsu, Hock-Liew Eng, and King-Wah Chiu. 2022. "Liver Graft MicroRNAs Expression in Different Etiology of Acute Jaundice after Living Donor Liver Transplantation" Biology 11, no. 8: 1228. https://doi.org/10.3390/biology11081228