
Effects of Temperature on Transparent Exopolymer Particle Production and Organic Carbon Allocation of Four Marine Phytoplankton Species


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture Condition
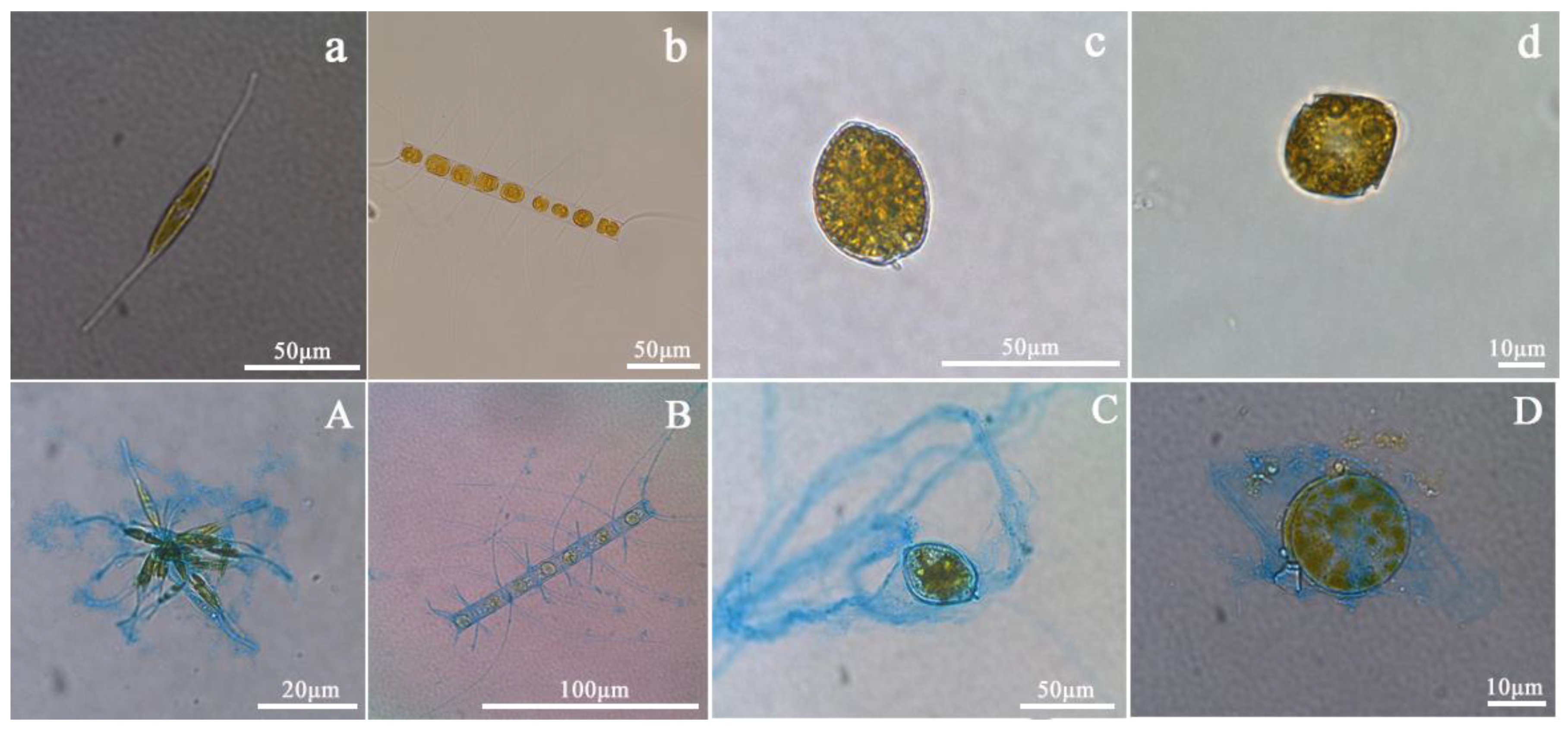
2.2. Measurement of Phytoplankton Abundance and Biomass
2.3. Determination of Chlorophyll a (Chl a) Concentrations
2.4. Colorimetric Determination of TEP
2.5. Calculation of TEP Carbon Content
2.6. Determination of the POC and DOC Content
2.7. Statistical Data Analysis
3. Results
3.1. Effects of Temperature on Phytoplankton Growth
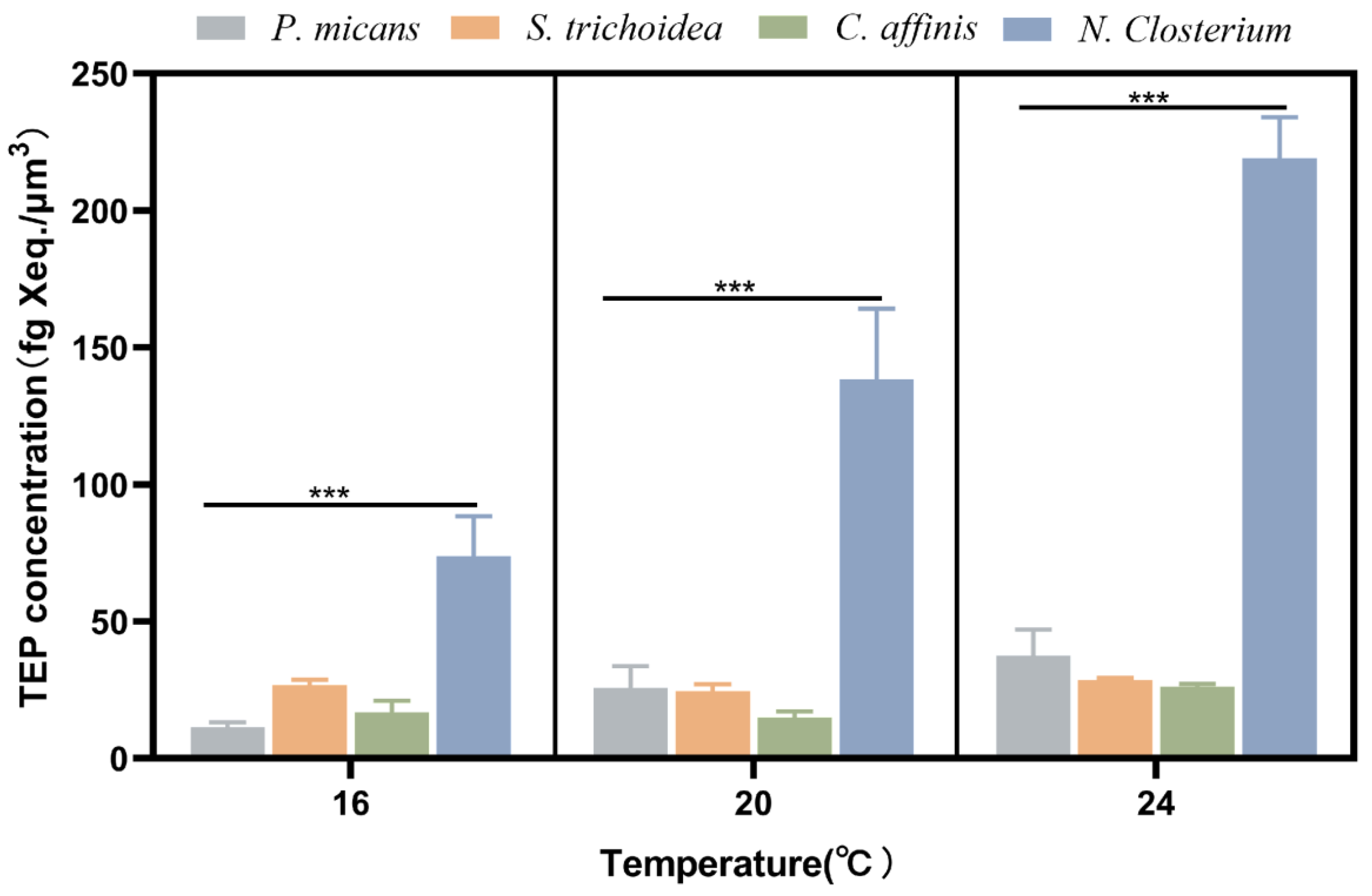
3.2. Effect of Temperature on TEP Production
3.3. Effects of Temperature on the Allocation of Organic Carbon Content in Phytoplankton
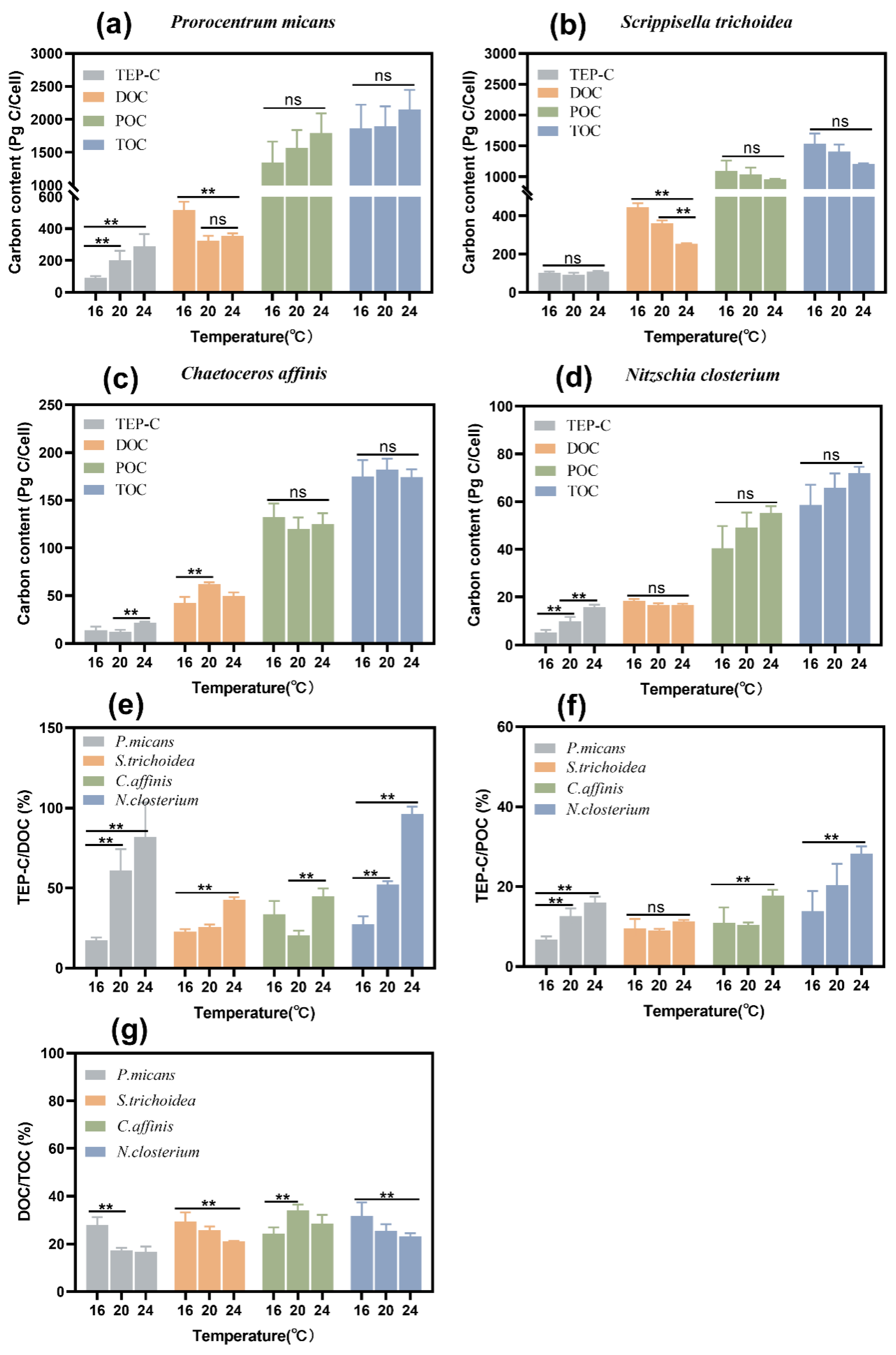
3.3.1. Effects of Temperature on DOC, POC, and TOC Contents
3.3.2. Effect of Temperature on TEP-C, TEP-C/DOC, and TEP-C/POC
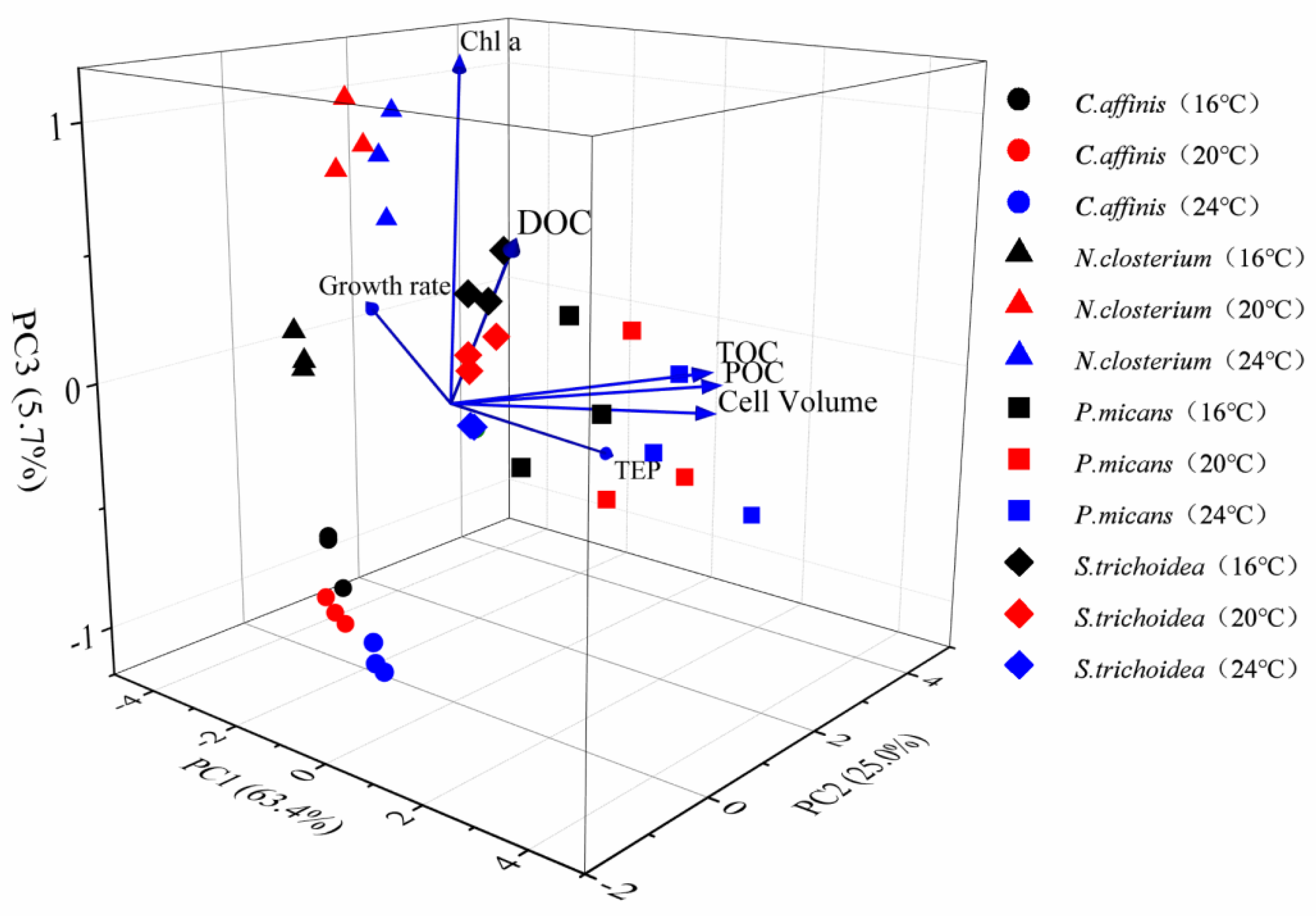
3.3.3. PCA Analysis
4. Discussion
4.1. Effects of Temperature on Phytoplankton Growth
4.2. Effect of Temperature on TEP Production of Phytoplankton
4.3. Relationship between TEP Production and Phytoplankton Physiologies
4.4. Effects of Temperature on TEP-C and Allocation of Organic Carbon Content
4.5. PCA Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharp, J.H. Excretion of organic matter by marine phytoplankton: Do healthy cells do it. Limnol. Oceanogr. 1977, 22, 381–399. [Google Scholar] [CrossRef]
- Fogg, G.E. The extracelluar products of algae. Oceanogr. Mar. Biol. Annu. Rev. 1966, 4, 195–212. [Google Scholar]
- Fogg, G.E. The ecological significance of extracellular products of phytoplankton photosynthesis. Bot. Mar. 1983, 26, 3–14. [Google Scholar] [CrossRef]
- Biddanda, B.; Benner, R. Carbon, nitrogen, and carbohydrate fluxes during the production of particulate and dissolved organic matter by marine phytoplankton. Limnol. Oceanogr. 1997, 42, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Alldredge, A.L.; Passow, U.; Logan, B.E. The abundance and significance of a class of large, transparent organic particles in the ocean. Deep.-Sea Res. Part I Oceanogr. Res. Pap. 1993, 40, 1131–1140. [Google Scholar] [CrossRef]
- Passow, U. Production of transparent exopolymer particles (TEP) by phyto-and bacterioplankton. Mar. Ecol. Prog. Ser. 2002, 236, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thornton, D.C.O. Diatom aggregation in the sea: Mechanisms and ecological implications. Eur. J. Phycol. 2002, 37, 149–161. [Google Scholar] [CrossRef]
- Burns, W.G.; Marchetti, A.; Kai, Z. Enhanced formation of transparent exopolymer particles (TEP) under turbulence during phytoplankton growth. J. Plankton Res. 2019, 41, 349–361. [Google Scholar] [CrossRef]
- Verdugo, P.; Alldredge, A.L.; Azam, F.; Kirchman, D.L.; Passow, U.; Santschi, P.H. The oceanic gel phase: A bridge in the DOM–POM continuum. Mar. Chem. 2004, 92, 67–85. [Google Scholar] [CrossRef]
- Engel, A.; Passow, U. Carbon and nitrogen content of transparent exopolymer particles (TEP) in relation to their Alcian Blue adsorption. Mar. Ecol. Prog. Ser. 2001, 219, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Lee, W.C.; Kim, H.C.; Jo, N.; Jang, H.K.; Kang, J.J.; Lee, D.; Kim, K.; Lee, S.H. Transparent exopolymer particle (TEPs) dynamics and contribution to particulate organic carbon (POC) in Jaran bay, Korea. Water 2020, 12, 1057. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.; Li, Q.P.; Yang, W.; Liu, X.; Wu, Z. Transparent exopolymer particle dynamics along a shelf-to-sea gradient and impacts on the regional carbon cycle. Sci. Total Environ. 2022, 808, 152117. [Google Scholar] [CrossRef] [PubMed]
- Precali, R.; Giani, M.; Marini, M.; Grilli, F.; Ferrari, C.R.; Pečar, O.; Paschini, E. Mucilaginous aggregates in the northern Adriatic in the period 1999–2002: Typology and distribution. Sci. Total Environ. 2005, 353, 10–23. [Google Scholar] [CrossRef]
- Novis, P.; Schallenberg, M.; Saulnier-Talbot, É.; Kilroy, C. The diatom Lindvia intermedia identified as the producer of nuisance pelagic mucilage in lakes. New Zeal. J. Bot. 2017, 55, 1–17. [Google Scholar] [CrossRef]
- Yi, B.; Chen, K.B.; Zhou, J.J.; Lv, Y.H. Characteristics of red tide in coastal region of South China from 2009 to 2016. Trans. Oceanol. Limnol. 2018, 2, 23–31. [Google Scholar]
- Wang, S.F.; Tang, D.L. Analysis of harmful algal blooms associated with ocean environments in the south China Sea. Mar. Sci. Bull. 2010, 29, 577–583. [Google Scholar]
- Zhang, C.; Dang, H.; Azam, F.; Benner, R.; Legendre, L.; Passow, U.; Polimene, L.; Robinson, C.; Suttle, C.A.; Jiao, N. Evolving paradigms in biological carbon cycling in the ocean. Natl. Sci. Rev. 2018, 4, 481–499. [Google Scholar] [CrossRef] [Green Version]
- Fukao, T.; Kimoto, K.; Kotani, Y. Production of transparent exopolymer particles by four diatom species. Fish. Sci. 2010, 76, 755–760. [Google Scholar] [CrossRef]
- Sancetta, C. Identifying marine diatoms and dinoflagellates. Mar. Geol. 1997, 136, 320–321. [Google Scholar] [CrossRef]
- Davison, I.R. Environmental effects on algal photosynthesis: Temperature. J. Phycol. 1991, 27, 2–8. [Google Scholar] [CrossRef]
- Claquin, P.; Probert, I.; Lefebvre, S.; Veron, B. Effects of temperature on photosynthetic parameters and TEP production in eight species of marine microalgae. Phycologia 2008, 44, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, A.P.; Hann, B.J. Warm spring and summer water temperatures in small eutrophic lakes of the Canadian prairies: Potential implications for phytoplankton and zooplankton. J. Plankton Res. 2009, 31, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Goldman, J.C.; Mann, R. Temperature-influenced variations in speciation and chemical composition of marine phytoplankton in outdoor mass cultures. J. Exp. Mar. Biol. Ecol. 1980, 46, 29–39. [Google Scholar] [CrossRef]
- Montagnes, D.J.S.; Franklin, M. Effect of temperature on diatom volume, growth rate, and carbon and nitrogen content: Reconsidering some paradigms. Limnol. Oceanogr. 2001, 46, 2008–2018. [Google Scholar] [CrossRef] [Green Version]
- Sunagawa, S.; Acinas, S.G.; Bork, P.; Bowler, C.; Babin, M.; Boss, E.; Cochrane, G.; de Vargas, C.; Follows, M.; Gorsky, G.; et al. Tara Oceans: Towards global ocean ecosystems biology. Nat. Rev. Microbiol. 2020, 18, 428–445. [Google Scholar] [CrossRef]
- Zlotnik, I.; Dubinsky, Z. The effect of light and temperature on DOC excretion by phytoplankton. Limnol. Oceanogr. 1989, 34, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.J.; Zhao, Y.F.; Zhu, M.L.; Liang, J.H.; Du, J.; Sun, X.X. Spatio-temporal Variation of Transparent Exopolymer Particles (TEP) and Their Sinking Flux in a Temperate Bay: Jiaozhou Bay, China. Estuar. Coast. Shelf Sci. 2020, 246, 107051. [Google Scholar] [CrossRef]
- Chen, J.; Guo, K.; Thornton, D.C.O.; Wu, Y. Effect of Temperature on the Release of Transparent Exopolymer Particles (TEP) and Aggregation by Marine Diatoms (Thalassiosira weissflogii and Skeletonema marinoi). J. Ocean. Univ. China 2021, 20, 56–66. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Division Rates. In Handbook of Phycological Methods: Culture Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University Press: Cambridge, UK, 1973; pp. 289–311. [Google Scholar]
- Passow, U.; Alldredge, A.L. A dye-binding assay for the spectrophotometric measurement of transparent exopolymer particles (TEP). Limnol. Oceanogr. 1995, 40, 1326–1335. [Google Scholar] [CrossRef]
- Passow, U. Transparent exopolymer particles (TEP) in aquatic environments. Prog. Oceanogr. 2002, 55, 287–333. [Google Scholar] [CrossRef] [Green Version]
- Deng, G.; Li, Y.-G.; Hu, H.-J.; Qi, Y.-Z.; Geng, Y.-H.; Li, Z.-K. Effects of temperature, light and pH on photosynthesis, and of light-dark cycle on growth rate and biomass of Scrippsiella trochoidea and Alexandrium tamarense. J. Wuhan Bot. Res. 2004, 22, 129–135. [Google Scholar]
- Wang, Z.F.; Zhang, Q.; Lv, H.Y. Effects of temperature, salinity, light and pH on the growth of red tide organisms Prorocentrum micans. Oceanol. Limnol. Sin. 2001, 1, 15–18. [Google Scholar]
- Engel, A.; Meyerhöfer, M.; Bröckel, K. Chemical and biological composition of suspended particles and aggregates in the Baltic Sea in summer (1999). Estuar. Coast. Shelf Sci. 2002, 55, 729–741. [Google Scholar] [CrossRef] [Green Version]
- Keys, M.; Tilstone, G.; Findlay, H.S.; Widdicombe, C.E.; Lawson, T. Effects of elevated CO2 and temperature on phytoplankton community biomass, species composition and photosynthesis during an experimentally induced autumn bloom in the western English Channel. Biogeosciences 2018, 15, 3203–3222. [Google Scholar] [CrossRef] [Green Version]
- Underwood, G.J.C.; Smith, D.J. Predicting epipelic diatom exopolymer concentrations in intertidal sediments from sediment Chlorophyll a. Microb. Ecol. 1998, 35, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Engel, A. Distribution of transparent exopolymer particles (TEP) in the northeast Atlantic Ocean and their potential significance for aggregation processes. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2004, 51, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Pedrotti, M.; Peters, F.; Beauvais, S.; Vidal, M.; Egge, J.; Jacobsen, A.; Marrase, C. Effects of nutrients and turbulence on the production of transparent exopolymer particles: A mesocosm study. Mar. Ecol. Prog. 2010, 419, 57–69. [Google Scholar] [CrossRef]
- Wu, C.Y.; Jiao, N.Z. Method of analysis of size spectra of marine Plankton by flow imaging techniques. Chin. High Technol. Lett. 2005, 15, 71–74. [Google Scholar]
- Lind, J.L.; Heimann, K.; Miller, E.A.; Vliet, C.; Hoogenraad, N.J.; Wetherbee, R. Substratum adhesion and gliding in a diatom are mediated by extracellular proteoglycans. Planta 1997, 203, 213–221. [Google Scholar] [CrossRef]
- Hoagland, K.D.; Rosowski, J.R.; Gretz, M.R.; Roemer, S.C. Diatom extracellular polymeric substances: Function, fine structure, chemistry, and physiology. J. Phycol. 1993, 29, 537–566. [Google Scholar] [CrossRef]
- Higgins, M.; Molino, P.; Mulvaney, P.; Wetherbee, R. The structure and nanomechanical properties of the adhesive mucilage that mediates diatom-substratum adhesion and motility. J. Phycol. 2003, 39, 1181–1193. [Google Scholar] [CrossRef]
- Holland, R.; Dugdale, T.M.; Wetherbee, R.; Brennan, A.B.; Finlay, J.A.; Callow, J.A.; Callow, M.E. Adhesion and motility of fouling diatoms on a silicone elastomer. Biofouling 2004, 20, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zheng, J.Y.; Yang, J.Y.; Lin, C.G.; Song, H.Q. Studies on the chemical compositions of extracellular polymeric substances from marine benthic diatoms. Mar. Environ. Sci. 2016, 35, 641–646. [Google Scholar]
- Higgins, M.J.; Crawford, S.A.; Mulvaney, P.; Wetherbee, R. The topography of soft, adhesive diatom “trails” as observed by atomic force microscopy. Biofouling 2000, 16, 133–139. [Google Scholar] [CrossRef]
- Higgins, M.; Crawford, S.A.; Mulvaney, P.; Wetherbee, R. Characterization of the adhesive mucilages secreted by live diatom cells using atomic force microscopy. Protist 2002, 153, 25–38. [Google Scholar] [CrossRef]
- Staats, N.; Stal, L.J.; Mur, L.R. Exopolysaccharide production by the epipelic diatom Cylindrotheca closterium: Effects of nutrient conditions. J. Exp. Mar. Biol. Ecol. 2000, 249, 13–27. [Google Scholar] [CrossRef]
- Goldman, J.C. Temperature effects on phytoplankton growth in continuous culture. Limnol. Oceanogr. 1977, 22, 932–936. [Google Scholar] [CrossRef]
- Harris, G. Phytoplankton Ecology: Structure, Function and Fluctuation; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Thompson, P.A.; Guo, M.; Harrison, P.J. Effects of variation in temperature. I. On the biochemical composition of eight species of marine phytoplankton. J. Phycol. 1992, 28, 481–488. [Google Scholar] [CrossRef]
- Baines, S.B.; Pace, M.L. The production of dissolved organic matter by phytoplankton and its importance to bacteria: Patterns across marine and freshwater systems. Limnol. Oceanogr. 1991, 36, 1078–1090. [Google Scholar] [CrossRef] [Green Version]
- Marañón, E.; Cermeño, P.; Pérez, V. Continuity in the photosynthetic production of dissolved organic carbon from eutrophic to oligotrophic waters. Mar. Ecol. Prog. Ser. 2005, 299, 7–17. [Google Scholar] [CrossRef]
- Castillo, C.R.; Sarmento, H.; Alvarez-Salgado, X.A.; Gasol, J.M.; Marraséa, C. Production of chromophoric dissolved organic matter by marine phytoplankton. Limnol. Oceanogr. 2010, 55, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Thornton, D.C.O. Primary Production in the Ocean. In Advances in Photosynthesis- Fundamental Aspects; InTech: Rijeka, Croatia, 2012; pp. 468–470. [Google Scholar]
- Seebah, S.; Fairfield, C.; Ullrich, M.S.; Passow, U. Aggregation and sedimentation of Thalassiosira weissflogii (diatom) in a warmer and more acidified future ocean. PLoS ONE 2014, 9, e112379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mari, X.; Rochelle-Newall, E.; Torréton, J.-P.; Pringault, O.; Jouon, A.; Migon, C. Water Residence Time: A Regulatory Factor of the DOM to POM Transfer Efficiency. Limnol. Oceanogr. 2007, 52, 808–819. [Google Scholar] [CrossRef] [Green Version]
- Chen, J. Factors Affecting Carbohydrate Production and the Formation of Transparent Exopolymer Particles (TEP) by Diatoms; Texas A&M University: College Station, TX, USA, 2014; pp. 1–269. [Google Scholar]
- Nagata, T. Production mechanisms of dissolved organic matter. In Microbial Ecology of the Oceans; Liss/Wiley: New York, NY, USA, 2000; pp. 121–152. [Google Scholar]
- Mari, X.; Passow, U.; Migon, C.; Burd, A.B.; Legendre, L. Transparent exopolymer particles: Effects on carbon cycling in the ocean. Prog. Oceanogr. 2017, 151, 13–37. [Google Scholar] [CrossRef] [Green Version]
- Zamanillo, M.; Ortega-Retuerta, E.; Nunes, S.; Rodríguez-Ros, P.; Dall’Osto, M.; Estrada, M.; Sala, M.M.; Simó, R. Main drivers of transparent exopolymer particle distribution across the surface Atlantic Ocean. Biogeosciences 2019, 16, 733–749. [Google Scholar] [CrossRef] [Green Version]
- Passow, U.; De La Rocha, C.L.; Fairfield, C.; Schmidt, K. Aggregation as a function of and mineral particles. Limnol. Oceanogr. 2014, 59, 532–547. [Google Scholar] [CrossRef]
- Facchinelli, A.; Sacchi, E.; Mallen, L. Multivariate statistical and GIS-based approach to identify heavy metal sources in soils. Environ. Pollut. 2001, 114, 313–324. [Google Scholar] [CrossRef]
- Xiao, W.; Liu, X.; Irwin, A.J.; Laws, E.A.; Wang, L.; Chen, B.; Zeng, Y.; Huang, B. Warming and eutrophication combine to restructure diatoms and dinoflagellates. Water Res. 2017, 128, 206–216. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Equivalent Spherical/Cone Diameter (μm) | Cell Volume (×103 μm3) | Initial Cell Density (Cells·mL−1) | Collection Site |
|---|---|---|---|---|
| Prorocentrum micans | 37.30 ± 3.29 | 15.19 ± 5.24 | 200 | Daya Bay |
| Nitzschia closterium | 22.44 ± 3.38 | 0.10 ± 0.02 | 3000 | Pearl River Estuary |
| Chaetoceros affinis | 15.29 ± 1.83 | 1.12 ± 0.41 | 1000 | Hong Kong Tsing Yi |
| Scrippisella trichoidea | 26.65 ± 4.90 | 7.45 ± 0.29 | 1000 | Pearl River Estuary |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, K.; Chen, J.; Yuan, J.; Wang, X.; Xu, S.; Hou, S.; Wang, Y. Effects of Temperature on Transparent Exopolymer Particle Production and Organic Carbon Allocation of Four Marine Phytoplankton Species. Biology 2022, 11, 1056. https://doi.org/10.3390/biology11071056
Guo K, Chen J, Yuan J, Wang X, Xu S, Hou S, Wang Y. Effects of Temperature on Transparent Exopolymer Particle Production and Organic Carbon Allocation of Four Marine Phytoplankton Species. Biology. 2022; 11(7):1056. https://doi.org/10.3390/biology11071056
Chicago/Turabian StyleGuo, Kangli, Jie Chen, Jian Yuan, Xiaodong Wang, Shuaishuai Xu, Shengwei Hou, and Yan Wang. 2022. "Effects of Temperature on Transparent Exopolymer Particle Production and Organic Carbon Allocation of Four Marine Phytoplankton Species" Biology 11, no. 7: 1056. https://doi.org/10.3390/biology11071056