
Diversity and Assemblage of Harmful Algae in Homestead Fish Ponds in a Tropical Coastal Area
 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
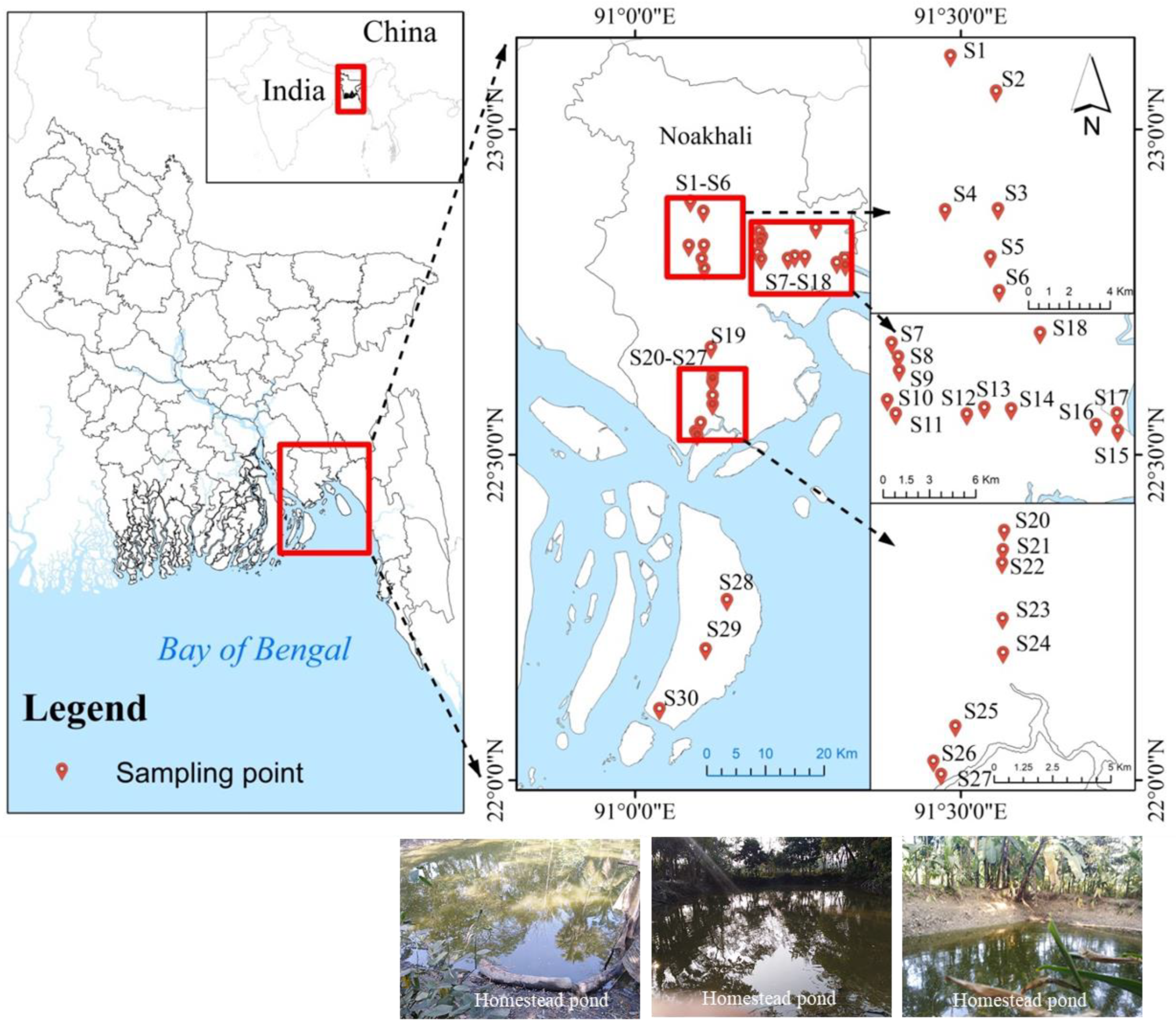
2.1. Study Site and Sampling Design
2.2. Water Sample Collection, Identification and Enumeration of Algae
2.3. Diversity Indices
2.4. Physico-Chemical Parameters
2.5. Statistical Analysis of Collected Data
2.6. Ethical Statement
3. Results and Discussion
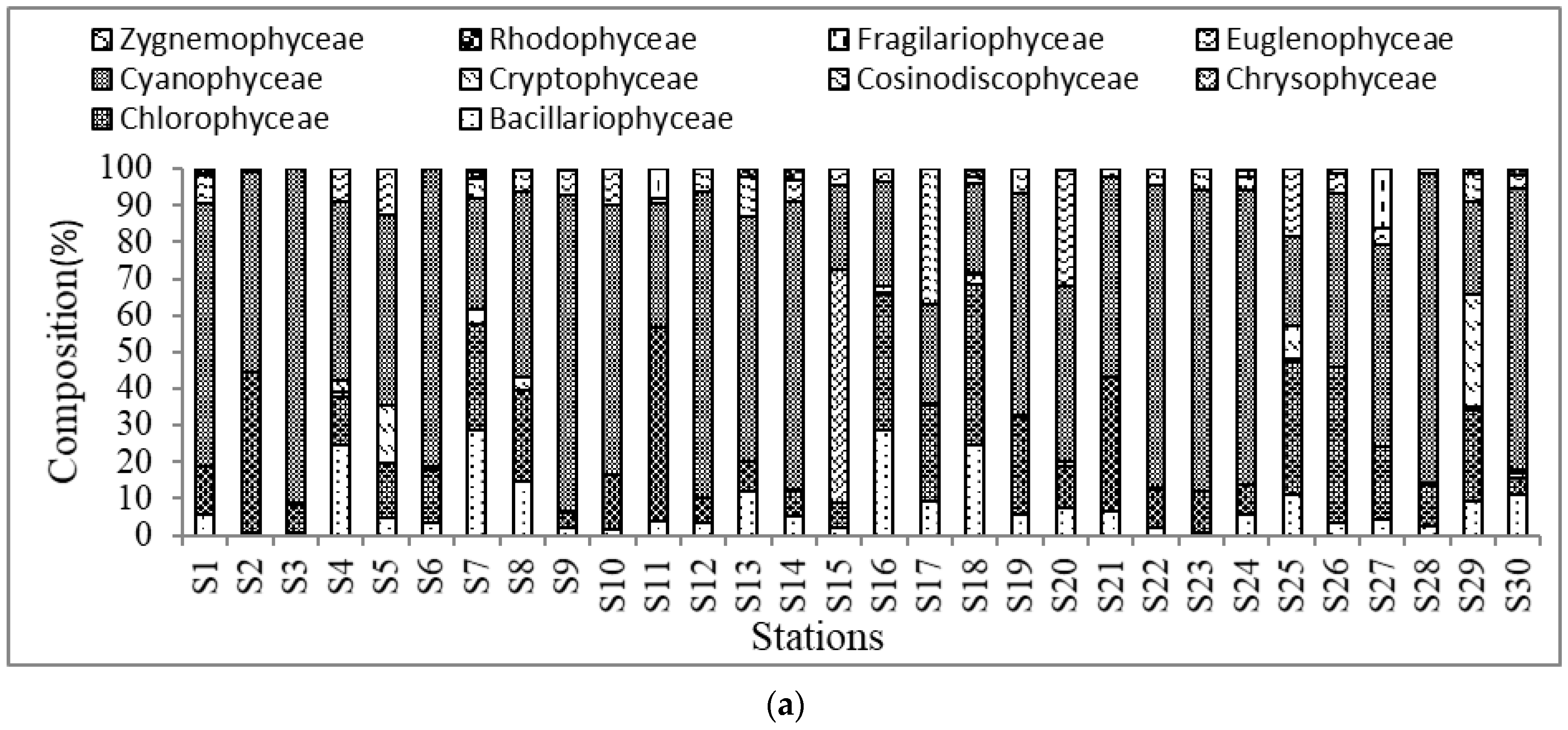
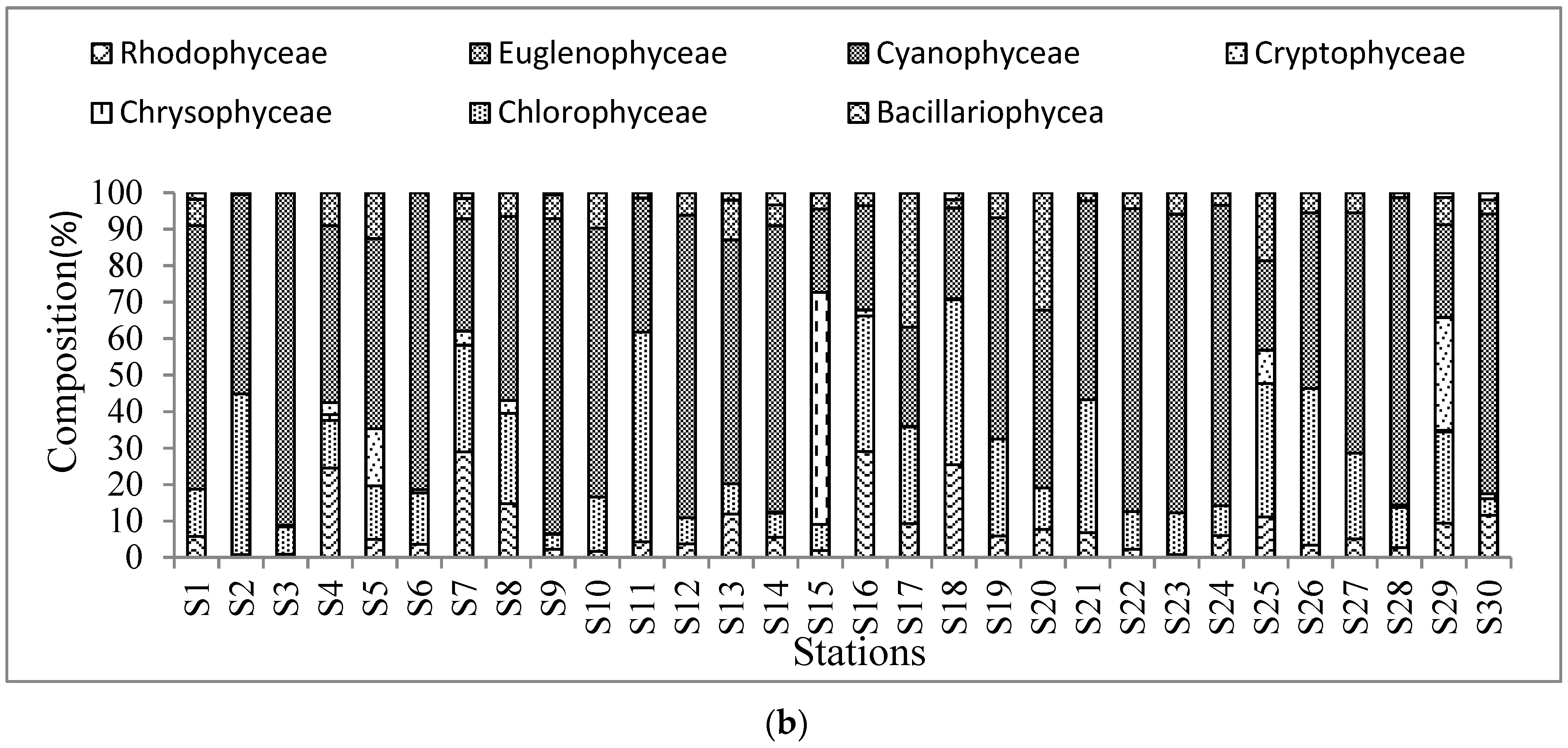
3.1. Algae Community and Composition
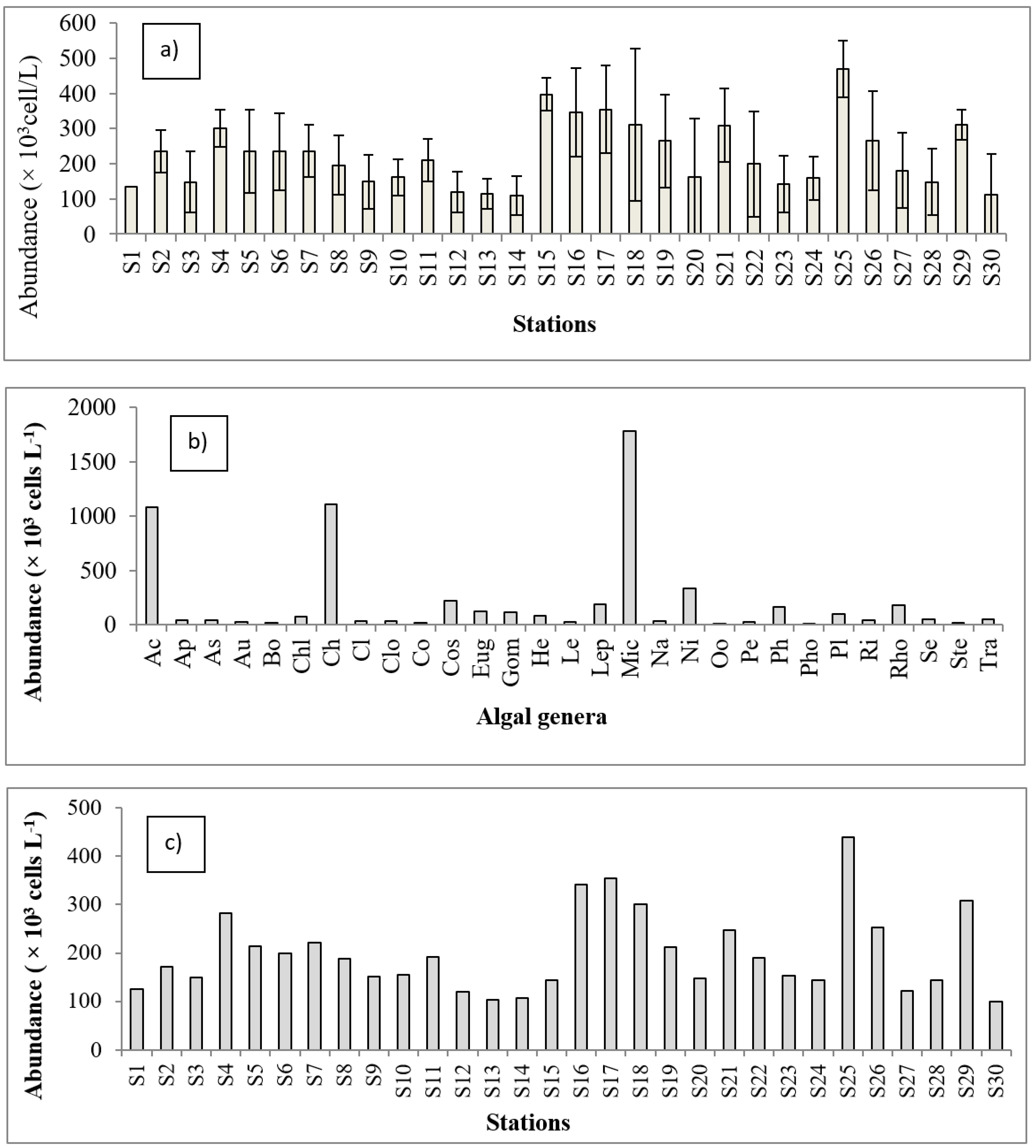
3.2. Algal Community Abundance and Diversity Indices
3.3. Assemblage of Algae
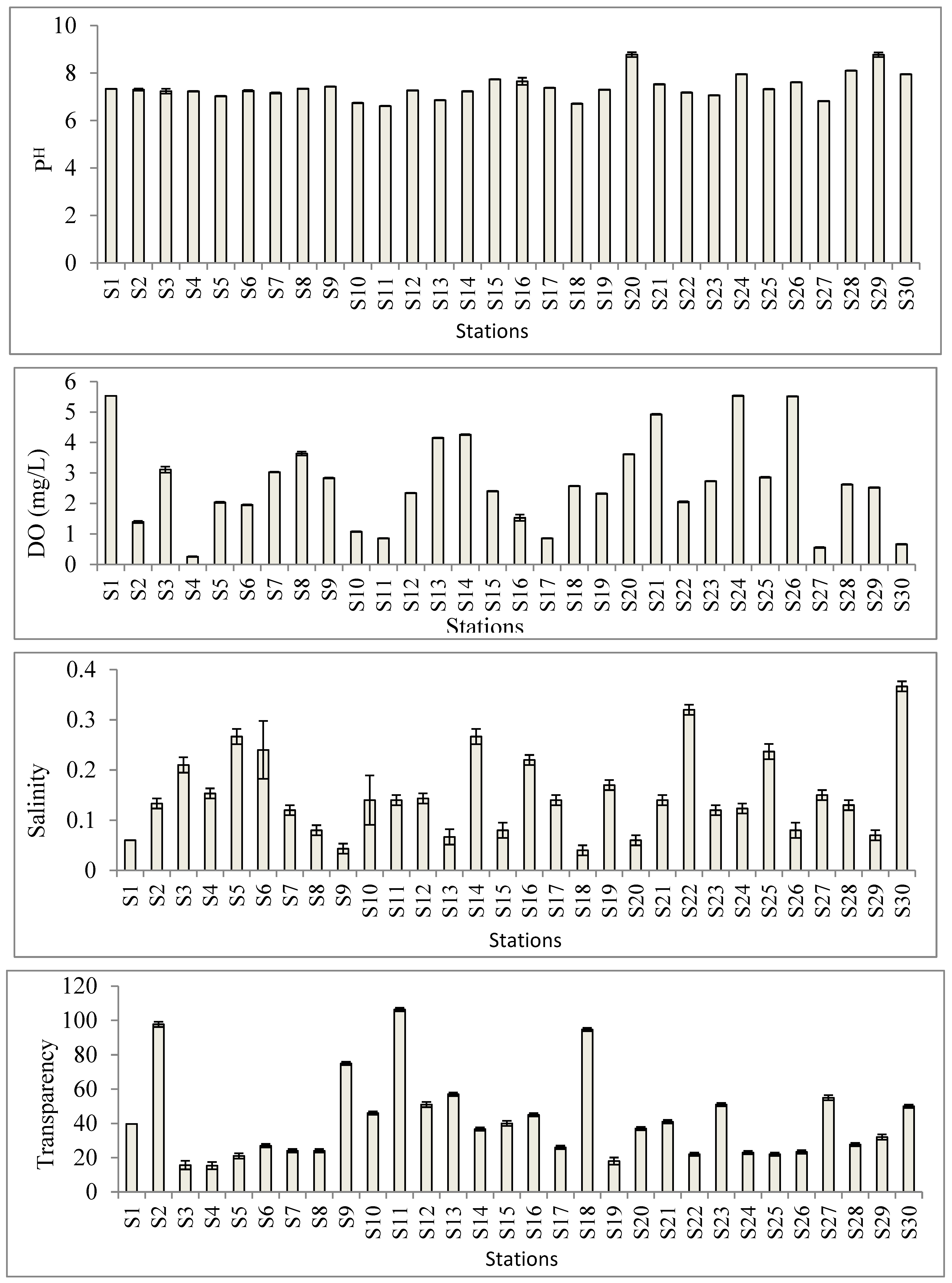
3.4. Physico-Chemical Parameters
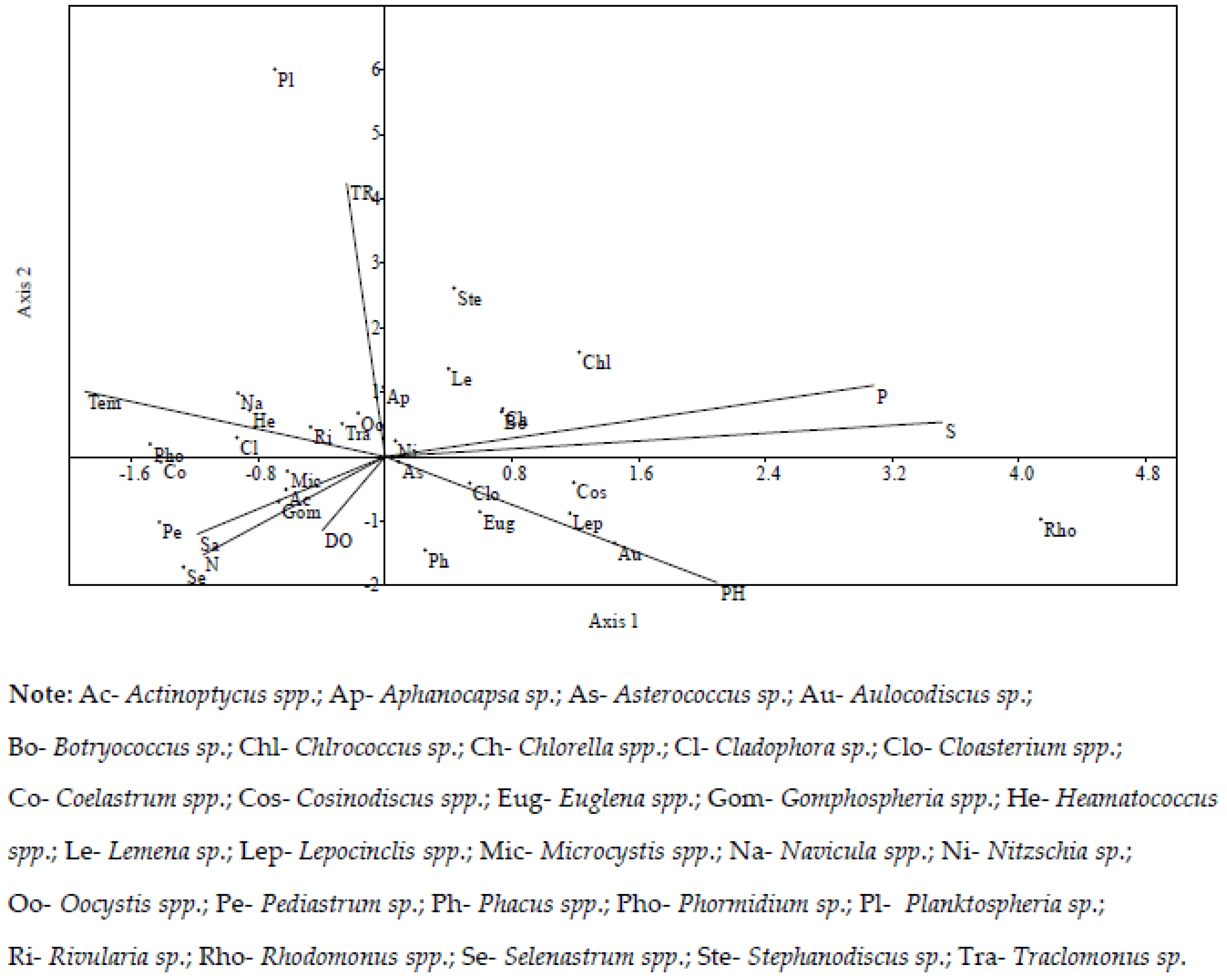
3.5. Relationship between Physico-Chemical Parameters and Biological Variables
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wickstead, J.H. Introduction to the Study of Tropical Plankton; Hutchinson: Paris, France, 1965. [Google Scholar]
- Suseela, M.R. Conservation and Diversity of Fresh Water Algae. In Biology and Biodiversity of Microalgae; Anand, N., Ed.; Centre for Advanced Studies in Botany, University of Madras: Chennai, India, 2009; pp. 1–41. [Google Scholar]
- Telesh, I.V. Plankton of the Baltic estuarine ecosystem with emphasis on Neva estuary: A review of present knowledge and research perspectives. Mar. Pollut. Bull. 2004, 49, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, A.F. A comparative study of phytoplankton communities of some rivers, creeks and burrow pits in the Niger Delta area. J. Appl. Sci. Environ. Manag. 2000, 4, 41–46. [Google Scholar]
- Nuccio, C.; Melillo, C.; Massi, L.; Innamorati, M. Phytoplankton abundance, community structure and diversity in the eutrophicated Orbetello lagoon (Tuscany) from 1995 to 2001. Oceanol. Acta 2003, 26, 15–25. [Google Scholar] [CrossRef]
- Song, X.; Huang, L.; Zhang, J.; Huang, X.; Zhang, J.; Yin, J.; Tan, Y.; Liu, S. Variation of phytoplankton biomass and primary production in Daya Bay during spring and summer. Mar. Pollut. Bull. 2004, 49, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Bahaar, S.W.N.; Bhat, G.A. Aquatic bio diversity in the paddy fields of Kashmir valley (J and K), India. Asian J. Agric. Res. 2011, 5, 26–29. [Google Scholar]
- Margalef, R. Life forms of phytoplankton as survival alternatives in an unstable environment. Oceanol. Acta 1978, 1, 493–509. [Google Scholar]
- Al, M.A.; Akhtar, A.; Hassan, M.L.; Rahman, M.F.; Warren, A. An approach to analyzing environmental drivers of phytoplankton community patterns in coastal waters in the northern Bay of Bengal, Bangladesh. Reg. Stud. Mar. Sci. 2019, 29, 100642. [Google Scholar]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1971; Volume 3, p. 5. [Google Scholar]
- Krebs, C.H.J. Ecological Methodology; Harper Collins Publisher: New York, NY, USA, 1989; p. 645. [Google Scholar]
- Margalef, R. Perspectives in Ecological Theory; The University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Reynolds, C.S. The responsible of phytoplankton communities to changing lake environments. Swiss J. Hydrol. 1987, 49, 220–236. [Google Scholar] [CrossRef]
- Chaturvedi, R.K.; Sharma, K.P.; Sharma, K.; Bhardwaj, S.M.; Sharma, S. Plankton community of polluted waters around Sanganer, Jaipur. J. Environ. Pollut. 1999, 6, 77–84. [Google Scholar]
- Saravanakumar, A.; Rajkumar, M.; Thivakaran, G.A.; Serebiah, J.S. Abundance and seasonal variations of phytoplankton in the creek waters of western mangrove of Kachchh-Gujarat. J. Environ. Biol. 2008, 29, 271. [Google Scholar]
- Hossain, M.Y.; Begum, M.; Ahmed, Z.F.; Hoque, M.A.; Karim, M.A.; Wahab, M.A. A study on the effects of iso- phosphorus fertilizers on plankton production in fish ponds. South Pac. Stud. 2006, 26, 101–110. [Google Scholar]
- Belton, B.; Azad, A. The characteristics and status of pond aquaculture in Bangladesh. Aquaculture 2012, 358, 196–204. [Google Scholar] [CrossRef]
- E-Jahan, K.M.; Ahmed, M.; Belton, B. The impacts of aquaculture development on food security: Lessons from Bangladesh. Aquac. Res. 2010, 41, 481–495. [Google Scholar] [CrossRef]
- Akter, S.; Rahman, M.M.; Faruk, A.; Bhuiyan, M.N.M.; Hossain, A.; Asif, A.A. Qualitative and quantitative analysis of phytoplankton in culture pond of Noakhali district, Bangladesh. Int. J. Fish. Aquat. Stud. 2018, 6, 371–375. [Google Scholar]
- Sarker, M.M.; Hossain, M.B.; Islam, M.M.; Kamal, A.H.M.; Idris, M.H. Unravelling the diversity and assemblage of phytoplankton in homestead ponds of central coastal belt, Bangladesh. Aquac. Res. 2020, 52, 167–184. [Google Scholar] [CrossRef]
- Khan, N.S.; Bari, J.B.A. The effects of physico-chemical parameters on plankton distribution in poultry manure and artificial formulated feed treated fish ponds, Noakhali, Bangladesh. Int. J. Fish. Aquat. Stud. 2019, 7, 1–7. [Google Scholar]
- Khan, N.S.; Uddin, A.; Bari, J.B.A.; Tisha, N.A. Evaluation the potentiality of ancient ponds by palmer’s Algal pollution index, Noakhali, Bangladesh. Int. J. Fish. Aquat. Res. 2019, 4, 28–31. [Google Scholar]
- Ahmed, S.; Rahman, A.F.M.A.; Hossain, M.B. Phytoplankton biodiversity in seasonal waterlogged paddy fields, Bangladesh. Ecologia 2013, 1, 1–8. [Google Scholar] [CrossRef]
- Hossain, M.Y.; Jasmine, S.; Ibrahim, A.H.M.; Ahmed, Z.F.; Ohtomi, J.; Fulanda, B.; Wahab, M.A. A preliminary observation on water quality and plankton of an earthen fish pond in Bangladesh: Recommendations for future studies. Pak. J. Biol. Sci. 2007, 10, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Begum, M.; Hossain, M.Y.; Wahab, M.A.; Ahmed, Z.F.; Alam, M.J.; Shah, M.M.R.; Jasmine, S. Effects of iso- nutrient fertilization on plankton production in earthen ponds of Bangladesh. Pak. J. Biol. Sci. 2007, 10, 1221–1228. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, A.H.; Al Mamun, A. Physio-chemical conditions and plankton population of two fishponds in Khulna. Univ. J. Zool. Rajshahi Univ. 2006, 25, 41–44. [Google Scholar] [CrossRef]
- Affan, A.; Jewel, A.S.; Haque, M.; Khan, S.; Lee, J.B. Seasonal cycle of phytoplankton in aquaculture ponds in Bangladesh. ALGAE 2005, 20, 43–52. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. A Key to the More Frequently Occurring Freshwater Algae. In Freshwater Algae: Identification, Enumeration and Use as Bioindicators; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 141–248. [Google Scholar]
- Belcher, H.; Swale, E. A Beginner’s Guide to Freshwater Algae; HMSO: London, UK, 1976.
- Davis, C.C. The Marine and Fresh-Water Plankton; Michigan State University: Ann Arbor, MI, USA, 1955. [Google Scholar]
- Manickam, N.; Bhavan, P.S.; Vijayan, P.; Sumathi, G. Phytoplankton species diversity in the parambikulam-aliyar irrigational canals (Tamil Nadu, India). Int. J. Pharma Bio Sci. 2012, 3, 289–300. [Google Scholar]
- Manickam, N.; Bhavan, P.S.; Santhanam, P.; Muralisankar, T.; Kumar, S.D.; Balakrishnan, S.; Ananth, S.; Devi, A.S. Phytoplankton bio-diversity in the two perennial lakes of Coimbatore, Tamil Nadu, India. Acta Ecol. Sin. 2020, 40, 81–89. [Google Scholar] [CrossRef]
- Pitchaikani, J.S.; Lipton, A.P. Nutrients and phytoplankton dynamics in the fishing grounds off Tiruchendur coastal waters, Gulf of Mannar, India. SpringerPlus 2016, 5, 1405. [Google Scholar] [CrossRef]
- Pielou, E.C. Species-diversity and pattern-diversity in the study of ecological succession. J. Theor. Biol. 1966, 10, 370–583. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 125. [Google Scholar]
- APHA. Standard Methods for Examination of Water and Waste Water, 18th ed.; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics Software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Sharma, R.C.; Tiwari, V. Phytoplankton diversity in relation to physico-chemical environmental variables of Nachiketa Tal, Garhwal Himalaya. Biodivers. Int. J. 2018, 2, 102–110. [Google Scholar]
- Shah, M.M.R.; Hossain, M.Y.; Begum, M.; Ahmed, Z.F.; Ohtomi, J.; Rahman, M.M.; Alam, M.J.; Islam, M.A.; Fulanda, B. Seasonal variations of phytoplankton community structure and production in relation to environmental factors of the southwest coastal waters of Bangladesh. J. Fish. Aquat. Sci. 2008, 3, 102–113. [Google Scholar]
- Gogoi, P.; Sinha, A.; Sarkar, S.D.; Chanu, T.N.; Yadav, A.K.; Koushlesh, S.K.; Borah, S.; Das, S.K.; Das, B.K. Seanoal influence of physicochemical parameters on phytoplankton diversity and assemblage pattern in Kailash Khal, a tropical wetland, Sundarbans, India. Appl. Water Sci. 2019, 9, 156. [Google Scholar] [CrossRef]
- Iqbal, M.M.; Billah, M.M.; Haider, M.N.; Islam, M.S.; Payel, H.R.; Bhuiyan, M.K.A.; Dawood, M.A.O. Seasonal distribution of phytoplankton community in a subtrophical estuary of the south-eastern coast Bangladesh. Zool. Ecol. 2017, 27, 304–310. [Google Scholar] [CrossRef]
- Hossain, M.I.; Alam, M.M.; Kamal, B.M.M.; Galib, S.M. Investigation of phytoplankton and physico-chemical parameters in nursery, Growout and Broodstock Ponds. J. Sci. Res. 2013, 5, 555–571. [Google Scholar] [CrossRef]
- Chowdhury, M.M.R.; Mondol, M.R.K.; Sarker, C. Seasonal variation of plankton population of Borobila beel in Rangpur district. Univ. J. Zool. Rajshahi Univ. 2007, 26, 49–54. [Google Scholar] [CrossRef]
- Effendi, H.; Kawaroe, M.; Lestari, D.F.; Mursalin; Permadi, T. Distribution of phytoplankton diversity and abundance in Mahakam Delta, East Kalimantan. Procedia Environ. Sci. 2016, 33, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.R.A.; Pramanik, M.M.; Hasan, M.M. Diversity indices of plankton communities in the River Meghna of Bangladesh. Int. J. Fish. Aquat. Stud. 2017, 5, 330–334. [Google Scholar]
- Ahsan, D.A.; Kabir, A.N.; Rahman, M.M.; Mahabub, S.; Yesmin, R.; Faruque, M.H.; Naser, M.N. Plankton composition, abundance and diversity in hilsa (Tenualosa ilisha) migratory rivers of Bangladesh during spawning season. Dhaka Univ. J. Biol. Sci. 2012, 21, 177–189. [Google Scholar] [CrossRef]
- Sidik, M.J.; Rashed-Un-Nabi, M.; Hoque, M.A. Distribution of phytoplankton community in relation to environmental parameters in cage culture area of Sepanggar Bay, Sabah, Malaysia. Estuar. Coast. Shelf Sci. 2008, 80, 251–260. [Google Scholar] [CrossRef]
- Gao, X.L.; Song, J.M. Phytoplankton distribution and their relationship with the environment in the Changjiang Estuary, China. Mar. Pollut. Bull. 2005, 50, 327–335. [Google Scholar] [CrossRef]
- Huang, L.; Jian, W.; Song, X.; Huang, X.; Liu, S.; Qian, P.; Yin, K.; Wu, M. Species diversity and distribution for phytoplankton of the Pearl River estuary during rainy and dry seasons. Mar. Pollut. Bull. 2004, 49, 588–596. [Google Scholar] [CrossRef]
- Aktan, Y.; Tufekci, V.; Tufekci, H.; Aykulu, G. Distribution patterns, biomass estimates and diversity of phytoplankton in Izmit Bay (Turkey). Estuar. Coast. Shelf Sci. 2005, 64, 372–384. [Google Scholar] [CrossRef]
- Haider, M.A.; Shahriar, S.I.M.; Hosen, M.H.A.; Chhanda, M.S.; Khatun, M.M. A study on water quality parameters and benthos abundance in freshwater homestead ponds of Dinajpur, Bangladesh. Int. J. Fish. Aquat. Stud. 2017, 5, 27–32. [Google Scholar]
- Munni, M.A.; Fardus, Z.; Mia, M.Y.; Afrin, R. Assessment of pond water quality for fish culture: A case study of Santosh region in Tangail, Bangladesh. J. Environ. Sci. Nat. Resour. 2013, 6, 157–162. [Google Scholar] [CrossRef]
- Sharma, R.C.; Singh, N.; Chauhan, A. The influence of physico-chemical parameters on phytoplankton distribution in a head water stream of Garhwal Himalayas: A case study. Egypt. J. Aquat. Res. 2016, 42, 11–21. [Google Scholar] [CrossRef]
- Ahmed, K.K.U.; Ahamed, S.U.; Hossain, M.R.A.; Ahmed, T.; Barman, S. Quantitative and qualitative assessment of plankton: Some ecological aspect and water quality parameters of river Meghna, Bangladesh. Bangladesh J. Fish. Res. 2003, 7, 131–140. [Google Scholar]
- Hossain, M.A.R. An overview of fisheries sector of Bangladesh. Res. Agric. Livest. Fish. 2014, 1, 109–126. [Google Scholar] [CrossRef]
- Thillai, R.K.; Rajkumar, M.; Sun, J.; Ashok, P.V.; Perumal, P. Seasonal variations of phytoplankton diversity in the Coleroon coastal waters, southeast coast of India. Acta Oceanol. Sin. 2010, 29, 97–108. [Google Scholar] [CrossRef]
- Sultana, T.; Saifullah, A.S.M.; Moniruzzaman, M.; Rpy, S.; Al Mamun, S. Assessment of surface water quality: A case study of Tangail Municiple area, Bangladesh. Bangladesh J. Sci. Res. 2012, 25, 155–163. [Google Scholar]
- Gambhir, R.S.; Kapoor, V.; Nirola, A.; Sohi, R.; Bansal, V. Water pollution: Impact of pollutants and new promising techniques in purification process. J. Hum. Ecol. 2012, 37, 103–109. [Google Scholar] [CrossRef]
- Hossain, M.S.; Uddin, M.J.; Fakhruddin, A.N.M. Impacts of shrimp farming on the coastal environment of Bangladesh and approach for management. Rev. Environ. Sci. Bio/Technol. 2013, 12, 313–332. [Google Scholar] [CrossRef]
- Zebek, E. Response of planktonic cyanobacteria and periphyton assemblages to physicochemical properties of stormwater in a shallow urban lake. J. Elem. 2015, 20. [Google Scholar] [CrossRef]
- Macedo, M.F.; Duarte, P.; Mendes, P.; Ferreira, J.G. Annual variation of environmental variables, phytoplankton species composition and photosynthetic parameters in a coastal lagoon. J. Plankton Res. 2001, 23, 719–732. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae Class | Name of Algae Genera |
|---|---|
| * Bacillariophyceae (20) | Chaetoceros sp., Cocconies sp., Cosinodiscus spp., Cosmarium sp., Cyclotella sp., Diatom spp., Ellerbackia sp., Epithemia sp., Fragilaria sp., Gomphonema sp., Gyrosigma sp., Hemidiscus sp., Hyalodiscus sp., Meridion sp., Navicula spp., Nitzschia sp., Peronia sp., Tabellaria sp., Triceratium sp., Rhoicosphenia sp. |
| * Chlorophyceae (32) | Actinastrum sp., Ankristodesmus sp., Asterococcus sp., Botryococcus sp., Bulbocheate sp., Chlamydomonus sp., Chorella spp., Cladophora sp., Closterium spp., Coelestrum spp., Crucigenia sp., Draparnaldia sp., Enteromorpha sp., Eudorina sp., Gonium sp., Heamatococcus spp., Hydrodictyon sp., Mesotaenium sp., Oocystis spp., Pediastrum sp., Planktosphaera sp., Pleurococcus sp., Pleurotaenium sp., Scenedesmus spp., Selenastrum spp., Spirogyra sp., Staurastrum sp., Tetradron sp., Tetraspora sp., Volvox sp., Xanthidium sp., Zygnema sp. |
| * Chrysophyceae (2) | Mallomonas sp., Uroglena sp. |
| Cosinodiscophyceae (2) | Cerataulina sp., Stephanodiscus sp. |
| * Cryptophyceae (1) | Rhodomonus sp. |
| * Cyanophyceae (21) | Actinoptychus spp., Amphiprora sp., Anabaena sp, Aphanizomenon sp., Aphanocapsa sp., Aphanothece sp., Arthrospira sp., Aulacodiscus sp., Chlrococcus sp., Coelosphaerium spp., Gloecapsa spp., Gloeotrichia sp., Gomphosphaeria spp., Microcystis spp., Nostoc sp., Oscillatoria sp., Phormidium sp., Pseudanabena sp., Rivularia spp., Stigonema sp., Tolypothrix sp. |
| * Euglenophyceae (4) | Euglena spp., Lepocinclis spp., Phacus spp., Tracehlomonas sp. |
| Fragilariophyceae (2) | Asterionella sp., Thalassiothrix sp. |
| * Rhodophyceae (1) | Lemanea sp. |
| Zygnemophyceae (4) | Actinotaenium sp., Desmidium sp., Micrasterias sp., Mougeotia sp. |
| 0 | Tem | PH | DO | Sa | TR | N | P | S | BAC | CHL | CRY | CYA | EUG | RH | TA | H’ | J’ | d | D |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tem | 0.00 | ||||||||||||||||||
| PH | −0.38 | 0.00 | |||||||||||||||||
| DO | 0.33 | 0.24 | 0.00 | ||||||||||||||||
| Sa | −0.22 | −0.10 | −0.35 | 0.00 | |||||||||||||||
| TR | 0.17 | −0.34 | −0.23 | −0.29 | 0.00 | ||||||||||||||
| N | −0.02 | −0.08 | 0.14 | −0.03 | −0.06 | 0.00 | |||||||||||||
| P | 0.01 | −0.04 | −0.05 | −0.17 | 0.12 | −0.03 | 0.00 | ||||||||||||
| S | −0.24 | 0.02 | −0.17 | −0.10 | 0.05 | 0.17 | 0.75 | 0.00 | |||||||||||
| BAC | −0.04 | −0.04 | −0.22 | −0.04 | −0.05 | 0.20 | 0.35 | 0.47 | 0.00 | ||||||||||
| CHL | 0.07 | −0.10 | −0.05 | −0.04 | 0.19 | −0.15 * | 0.37 | 0.41 | 0.56 | 0.00 | |||||||||
| CRY | −0.38 | 0.45 | −0.04 | −0.07 | −0.17 | −0.24 | 0.42 | 0.46 | 0.25 | 0.28 | 0.00 | ||||||||
| CYA | 0.22 | −0.03 | −0.03 | 0.31 | −0.37 | −0.20 | −0.27 | −0.31 | −0.26 | −0.16 | −0.26 | 0.00 | |||||||
| EUG | −0.15 | 0.08 | −0.09 | 0.08 | −0.35 | 0.15 | 0.35 | 0.47 | 0.35 | 0.43 | 0.43 | −0.19 | 0.00 | ||||||
| RH | −0.10 | −0.05 | 0.22 | −0.29 | 0.22 | 0.15 | 0.24 | 0.22 | 0.35 | 0.10 | 0.35 | −0.51 | −0.07 | 0.00 | |||||
| TA | −0.05 | 0.06 | −0.18 | −0.02 | −0.14 | −0.12 | 0.41 | 0.52 | 0.60 | 0.78 | 0.40 | −0.05 | 0.64 | 0.00 | 0.00 | ||||
| H’ | −0.21 | 0.04 | −0.08 | 0.25 | −0.32 | −0.22 | −0.41 | −0.43 | −0.03 | 0.06 | −0.04 | 0.17 | −0.10 | −0.05 | 0.04 | 0.00 | |||
| J’ | 0.01 | −0.09 | −0.17 | 0.06 | −0.02 | −0.21 | −0.10 | −0.03 | 0.19 | 0.36 | 0.04 | 0.20 | 0.29 | 0.00 | 0.20 | 0.23 | 0.00 | ||
| d | −0.36 | 0.21 | 0.02 | 0.07 | 0.09 | 0.30 | −0.02 | 0.11 | 0.22 | 0.16 | 0.28 | −0.41 | 0.04 | 0.36 | 0.09 | 0.12 | −0.35 | 0.00 | |
| D | 0.19 | −0.04 | 0.19 | −0.12 | −0.03 | 0.03 | 0.00 | −0.13 | −0.47 | −0.62 | −0.31 | 0.06 | −0.31 | −0.18 | −0.36 | −0.34 | −0.66 | −0.38 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akter, L.; Ullah, M.A.; Hossain, M.B.; Karmaker, A.R.; Hossain, M.S.; Albeshr, M.F.; Arai, T. Diversity and Assemblage of Harmful Algae in Homestead Fish Ponds in a Tropical Coastal Area. Biology 2022, 11, 1335. https://doi.org/10.3390/biology11091335
Akter L, Ullah MA, Hossain MB, Karmaker AR, Hossain MS, Albeshr MF, Arai T. Diversity and Assemblage of Harmful Algae in Homestead Fish Ponds in a Tropical Coastal Area. Biology. 2022; 11(9):1335. https://doi.org/10.3390/biology11091335
Chicago/Turabian StyleAkter, Liza, Md. Akram Ullah, Mohammad Belal Hossain, Anu Rani Karmaker, Md. Solaiman Hossain, Mohammed Fahad Albeshr, and Takaomi Arai. 2022. "Diversity and Assemblage of Harmful Algae in Homestead Fish Ponds in a Tropical Coastal Area" Biology 11, no. 9: 1335. https://doi.org/10.3390/biology11091335