
Sex-Related Pain Behavioral Differences following Unilateral NGF Injections in a Rat Model of Low Back Pain

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. NGF-LBP Model
2.2. Cutaneous Hindpaw Mechanical Behavioral Assay
2.3. Cutaneous Trunk Mechanical Behavioral Assay
2.4. Algometer Deep Trunk Behavioral Assay
3. Statistical Analyses
4. Results
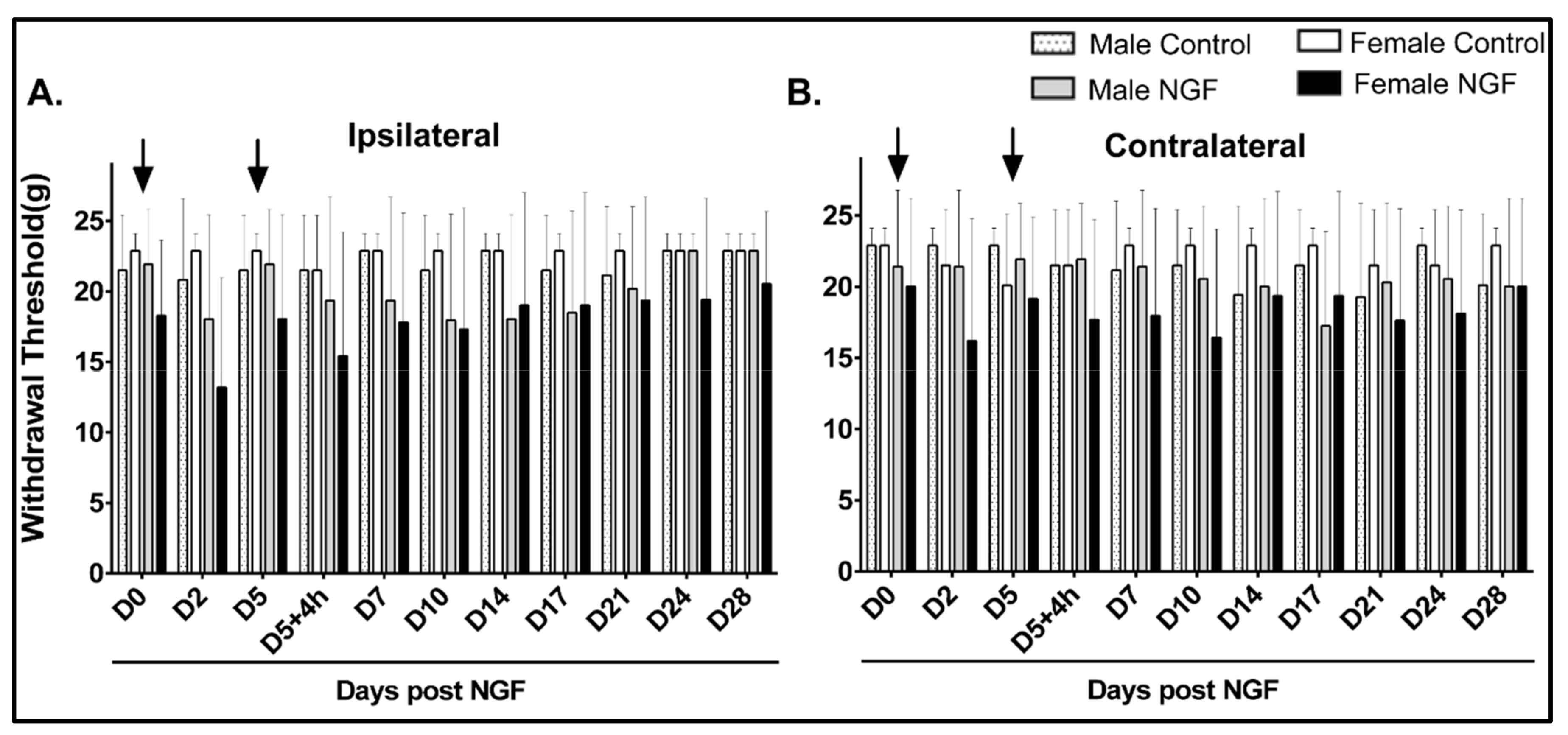
4.1. Cutaneous Hindpaw Mechanical Hypersensitivity
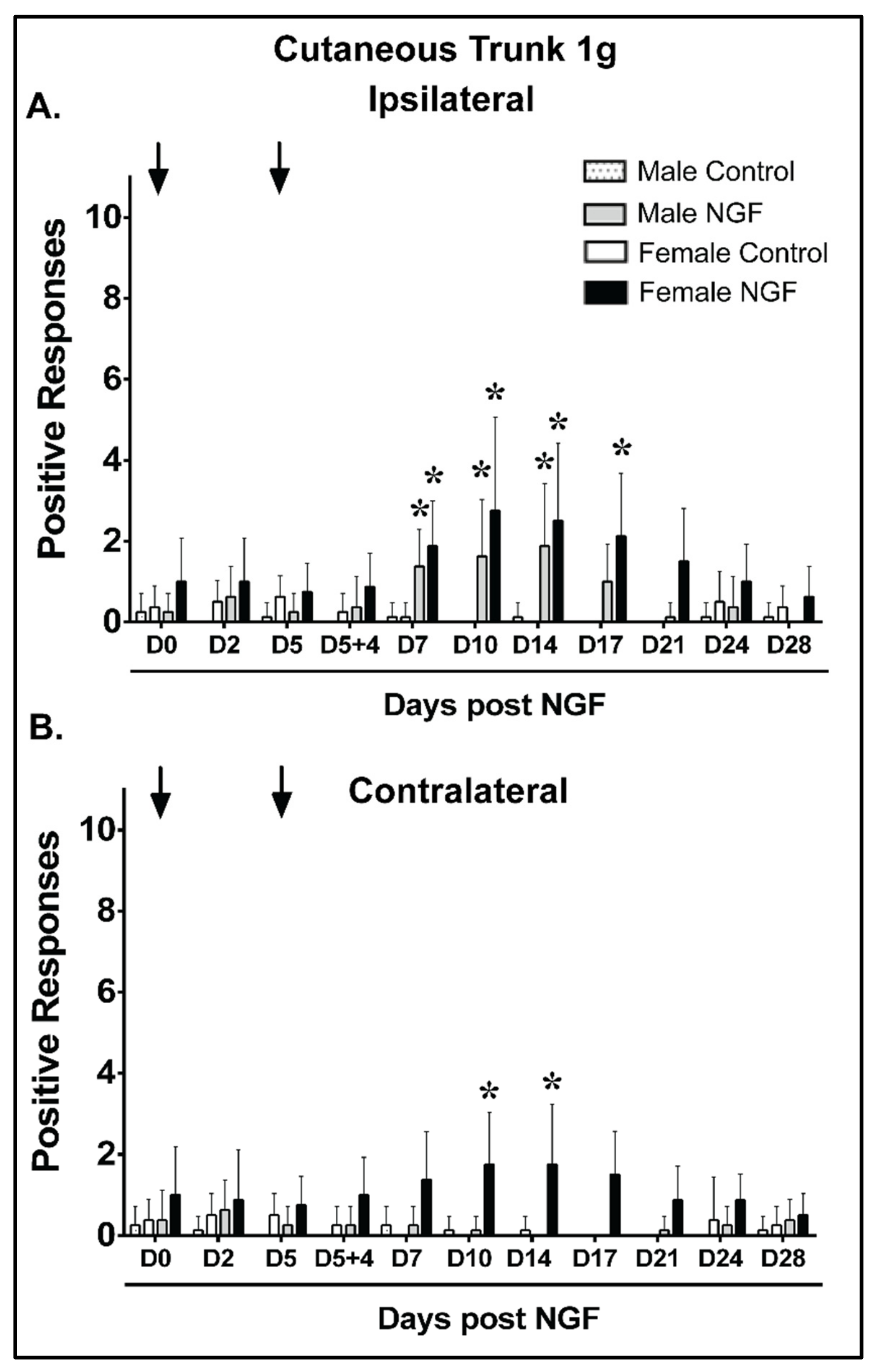
4.2. Cutaneous Trunk Mechanical Hypersensitivity (1 g)
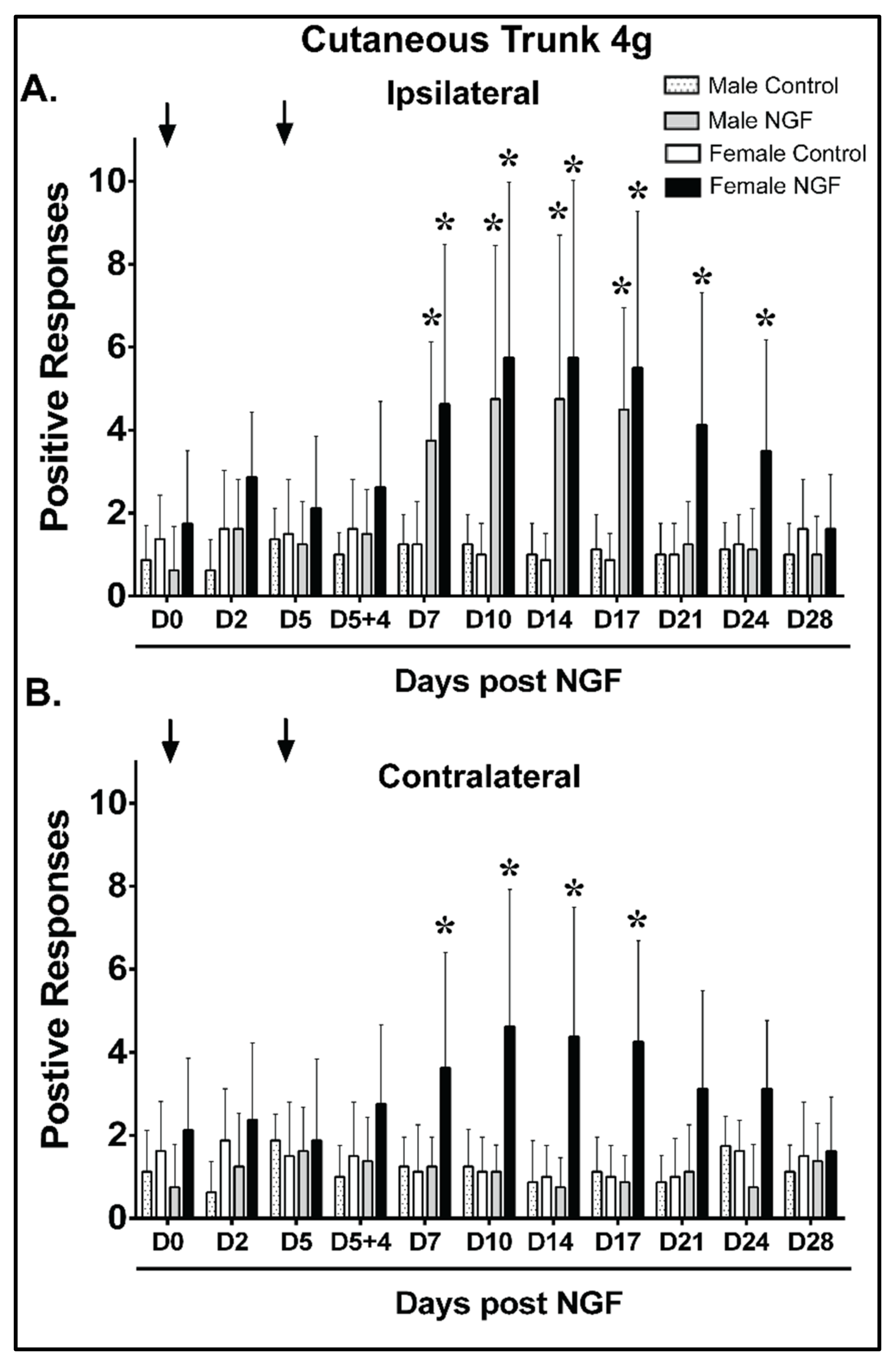
4.3. Cutaneous Trunk Mechanical Hypersensitivity (4 g)
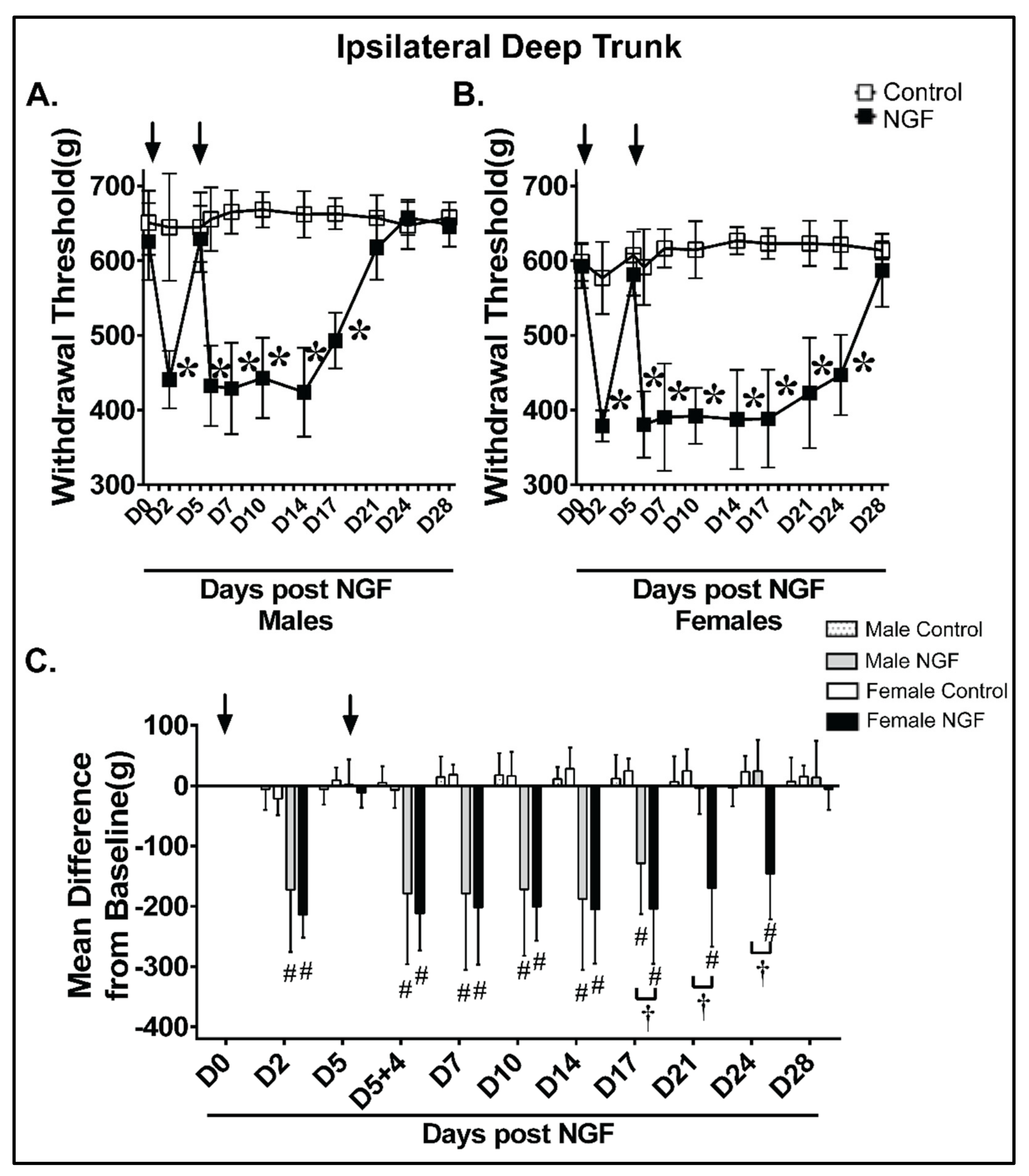
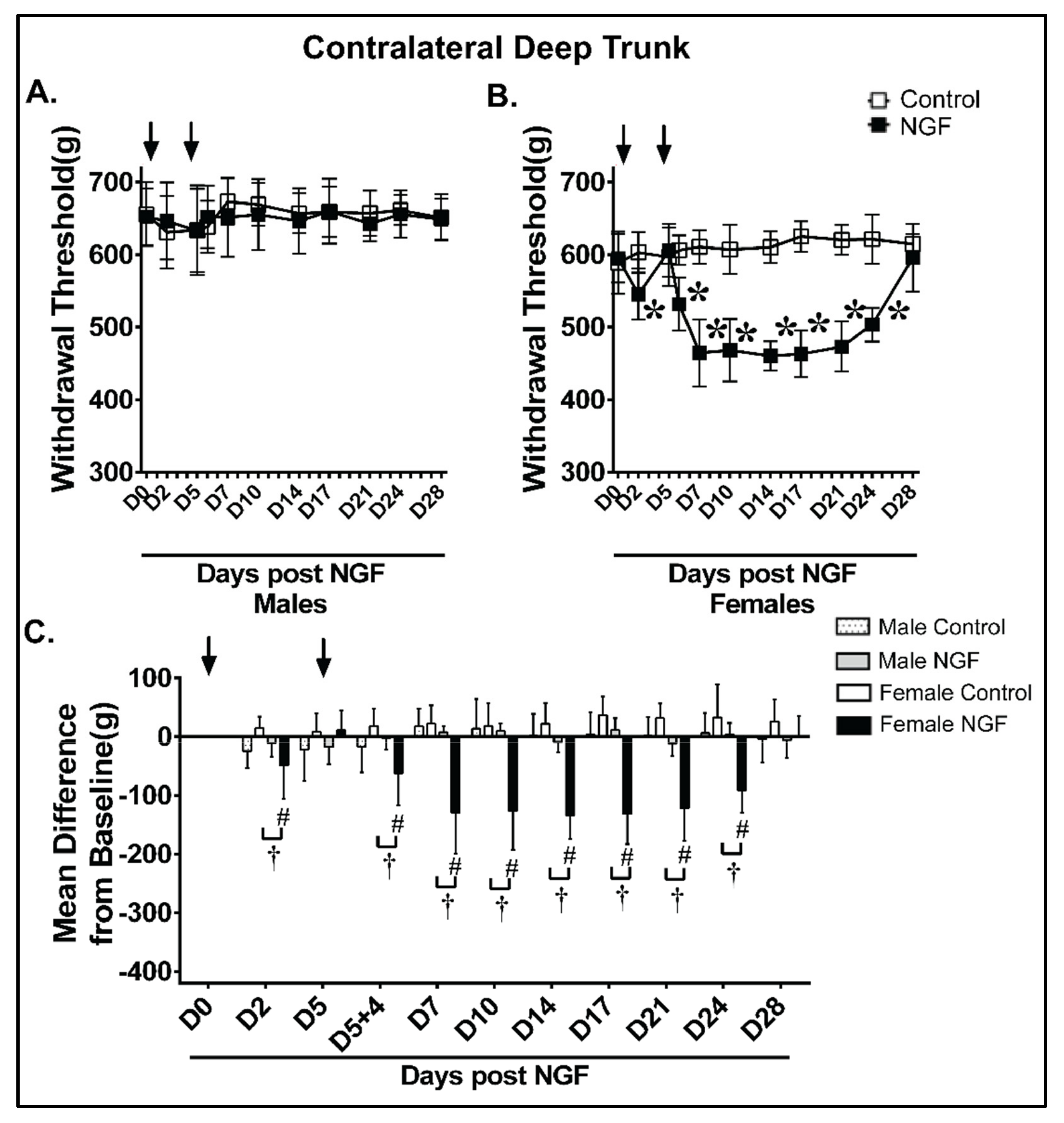
4.4. Deep Trunk Mechanical Hyperalgesia
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoy, D.; March, L.; Brooks, P.; Blyth, F.; Woolf, A.; Bain, C.; Williams, G.; Smith, E.; Vos, T.; Barendregt, J.; et al. The global burden of low back pain: Estimates from the Global Burden of Disease 2010 study. Ann. Rheum. Dis. 2014, 73, 968–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Gong, Y.; Liu, J.; Guo, Y.; Tang, H.; Qin, S.; Zhao, Y.; Wang, S.; Xu, Z.; Chen, B. Peripheral and Central Pathological Mechanisms of Chronic Low Back Pain: A Narrative Review. J. Pain Res. 2021, 14, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Allegri, M.; Montella, S.; Salici, F.; Valente, A.; Marchesini, M.; Compagnone, C.; Baciarello, M.; Manferdini, M.E.; Fanelli, G. Mechanisms of low back pain: A guide for diagnosis and therapy. F1000Research 2016, 5, 1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, J.; Bruce-Low, S.; Smith, D. A reappraisal of the deconditioning hypothesis in low back pain: Review of evidence from a triumvirate of research methods on specific lumbar extensor deconditioning. Curr. Med. Res. Opin 2014, 30, 865–911. [Google Scholar] [CrossRef] [PubMed]
- Deyo, R.A.; Weinstein, J.N. Low back pain. N. Engl. J. Med. 2001, 344, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Queme, L.F.; Jankowski, M.P. Sex differences and mechanisms of muscle pain. Curr. Opin. Physiol. 2019, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mogil, J.S. Qualitative sex differences in pain processing: Emerging evidence of a biased literature. Nat. Rev. Neurosci. 2020, 21, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Alhilou, A.M.; Shimada, A.; Svensson, C.I.; Svensson, P.; Ernberg, M.; Cairns, B.E.; Christidis, N. Sex-related differences in response to masseteric injections of glutamate and nerve growth factor in healthy human participants. Sci. Rep. 2021, 11, 13873. [Google Scholar] [CrossRef]
- Mogil, J.S. Perspective: Equality need not be painful. Nature 2016, 535, S7. [Google Scholar] [CrossRef] [Green Version]
- Mogil, J.S. Sex differences in pain and pain inhibition: Multiple explanations of a controversial phenomenon. Nat. Rev. Neurosci. 2012, 13, 859–866. [Google Scholar] [CrossRef]
- White, K.P.; Speechley, M.; Harth, M.; Ostbye, T. The London Fibromyalgia Epidemiology Study: The prevalence of fibromyalgia syndrome in London, Ontario. J. Rheumatol. 1999, 26, 1570–1576. [Google Scholar]
- Bergman, S.; Herrstrom, P.; Hogstrom, K.; Petersson, I.F.; Svensson, B.; Jacobsson, L.T. Chronic musculoskeletal pain, prevalence rates, and sociodemographic associations in a Swedish population study. J. Rheumatol 2001, 28, 1369–1377. [Google Scholar]
- Gerdle, B.; Bjork, J.; Coster, L.; Henriksson, K.; Henriksson, C.; Bengtsson, A. Prevalence of widespread pain and associations with work status: A population study. BMC Musculoskelet. Disord. 2008, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Fillingim, R.B.; King, C.D.; Ribeiro-Dasilva, M.C.; Rahim-Williams, B.; Riley, J.L., 3rd. Sex, gender, and pain: A review of recent clinical and experimental findings. J. Pain 2009, 10, 447–485. [Google Scholar] [CrossRef] [Green Version]
- Roussel, N.A.; Nijs, J.; Meeus, M.; Mylius, V.; Fayt, C.; Oostendorp, R. Central sensitization and altered central pain processing in chronic low back pain: Fact or myth? Clin. J. Pain 2013, 29, 625–638. [Google Scholar] [CrossRef] [Green Version]
- Strong, J.A.; Xie, W.; Bataille, F.J.; Zhang, J.M. Preclinical studies of low back pain. Mol. Pain 2013, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Qiu, S.; Riester, S.M.; Das, V.; Zhu, B.; Wallace, A.A.; van Wijnen, A.J.; Mwale, F.; Iatridis, J.C.; Sakai, D.; et al. Animal models for studying the etiology and treatment of low back pain. J. Orthop. Res. 2018, 36, 1305–1312. [Google Scholar] [CrossRef]
- Taguchi, T.; John, V.; Hoheisel, U.; Mense, S. Neuroanatomical pathway of nociception originating in a low back muscle (multifidus) in the rat. Neurosci. Lett. 2007, 427, 22–27. [Google Scholar] [CrossRef]
- Markman, J.D.; Bolash, R.B.; McAlindon, T.E.; Kivitz, A.J.; Pombo-Suarez, M.; Ohtori, S.; Roemer, F.W.; Li, D.J.; Viktrup, L.; Bramson, C.; et al. Tanezumab for chronic low back pain: A randomized, double-blind, placebo- and active-controlled, phase 3 study of efficacy and safety. Pain 2020, 161, 2068–2078. [Google Scholar] [CrossRef]
- Yang, S.; Huang, Y.; Ye, Z.; Li, L.; Zhang, Y. The Efficacy of Nerve Growth Factor Antibody for the Treatment of Osteoarthritis Pain and Chronic Low-Back Pain: A Meta-Analysis. Front. Pharmacol. 2020, 11, 817. [Google Scholar] [CrossRef]
- Cao, Z.; Li, Q.; Guo, J.; Li, Y.; Wu, J. Is Targeting Nerve Growth Factor Antagonist a New Option for Pharmacologic Treatment of Low Back Pain? A Supplemental Network Meta-Analysis of the American College of Physicians Guidelines. Front. Pharmacol. 2021, 12, 727771. [Google Scholar] [CrossRef]
- Ward, S.R.; Tomiya, A.; Regev, G.J.; Thacker, B.E.; Benzl, R.C.; Kim, C.W.; Lieber, R.L. Passive mechanical properties of the lumbar multifidus muscle support its role as a stabilizer. J. Biomech. 2009, 42, 1384–1389. [Google Scholar] [CrossRef] [Green Version]
- Zwambag, D.P.; Hurtig, M.B.; Vernon, H.; Brown, S.H.M. Investigation of the passive mechanical properties of spine muscles following disruption of the thoracolumbar fascia and erector spinae aponeurosis, as well as facet injury in a rat. Spine J. 2018, 18, 682–690. [Google Scholar] [CrossRef]
- Zwambag, D.P.; Ricketts, T.A.; Brown, S.H. Sarcomere length organization as a design for cooperative function amongst all lumbar spine muscles. J. Biomech. 2014, 47, 3087–3093. [Google Scholar] [CrossRef]
- Bang, W.S.; Lee, D.H.; Kim, K.T.; Cho, D.C.; Sung, J.K.; Han, I.B.; Kim, D.H.; Kwon, B.K.; Kim, C.H.; Park, K.S.; et al. Relationships between vitamin D and paraspinal muscle: Human data and experimental rat model analysis. Spine J. 2018, 18, 1053–1061. [Google Scholar] [CrossRef]
- Lieber, R.L.; Fridén, J. Functional and clinical significance of skeletal muscle architecture. Muscle Nerve 2000, 23, 1647–1666. [Google Scholar] [CrossRef]
- Barker, P.A.; Mantyh, P.; Arendt-Nielsen, L.; Viktrup, L.; Tive, L. Nerve Growth Factor Signaling and Its Contribution to Pain. J. Pain Res. 2020, 13, 1223–1241. [Google Scholar] [CrossRef]
- Hoheisel, U.; Reuter, R.; de Freitas, M.F.; Treede, R.D.; Mense, S. Injection of nerve growth factor into a low back muscle induces long-lasting latent hypersensitivity in rat dorsal horn neurons. Pain 2013, 154, 1953–1960. [Google Scholar] [CrossRef]
- Zhang, J.; Mense, S.; Treede, R.D.; Hoheisel, U. Prevention and reversal of latent sensitization of dorsal horn neurons by glial blockers in a model of low back pain in male rats. J. Neurophysiol. 2017, 118, 2059–2069. [Google Scholar] [CrossRef] [Green Version]
- De Azambuja, G.; Hortscht, U.; Hoheisel, U.; Oliveira Fusaro, M.C.; Mense, S.; Treede, R.D. Short-term swimming exercise attenuates the sensitization of dorsal horn neurons in rats with NGF-induced low back pain. Eur. J. Pain 2018, 22, 1409–1418. [Google Scholar] [CrossRef]
- Reed, N.R.; Reed, W.R.; Syrett, M.; Richey, M.L.; Frolov, A.; Little, J.W. Somatosensory behavioral alterations in a NGF-induced persistent low back pain model. Behav. Brain Res. 2022, 418, 113617. [Google Scholar] [CrossRef] [PubMed]
- Reed, W.R.; Little, J.W.; Lima, C.R.; Sorge, R.E.; Yarar-Fisher, C.; Eraslan, M.; Hurt, C.P.; Ness, T.J.; Gu, J.G.; Martins, D.F.; et al. Spinal Mobilization Prevents NGF-Induced Trunk Mechanical Hyperalgesia and Attenuates Expression of CGRP. Front. Neurosci. 2020, 14, 385. [Google Scholar] [CrossRef] [PubMed]
- Hirth, M.; Rukwied, R.; Gromann, A.; Turnquist, B.; Weinkauf, B.; Francke, K.; Albrecht, P.; Rice, F.; Hägglöf, B.; Ringkamp, M.; et al. Nerve growth factor induces sensitization of nociceptors without evidence for increased intraepidermal nerve fiber density. Pain 2013, 154, 2500–2511. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.; Kang, I.; Dong, X.D.; Christidis, N.; Ernberg, M.; Svensson, P.; Cairns, B.E. NGF-induced mechanical sensitization of the masseter muscle is mediated through peripheral NMDA receptors. Neuroscience 2014, 269, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Starkweather, A.R.; Lyon, D.E.; Kinser, P.; Heineman, A.; Sturgill, J.L.; Deng, X.; Siangphoe, U.; Elswick, R.K.; Greenspan, J.; Dorsey, S.G. Comparison of Low Back Pain Recovery and Persistence: A Descriptive Study of Characteristics at Pain Onset. Biol. Res. Nurs. 2016, 18, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Starkweather, A.R.; Ramesh, D.; Lyon, D.E.; Siangphoe, U.; Deng, X.; Sturgill, J.; Heineman, A.; Elswick, R.K., Jr.; Dorsey, S.G.; Greenspan, J. Acute Low Back Pain: Differential Somatosensory Function and Gene Expression Compared With Healthy No-Pain Controls. Clin. J. Pain 2016, 32, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Starkweather, A.R.; Heineman, A.; Storey, S.; Rubia, G.; Lyon, D.E.; Greenspan, J.; Dorsey, S.G. Methods to measure peripheral and central sensitization using quantitative sensory testing: A focus on individuals with low back pain. Appl. Nurs. Res. 2016, 29, 237–241. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Mills, C.; Leblond, D.; Joshi, S.; Zhu, C.; Hsieh, G.; Jacobson, P.; Meyer, M.; Decker, M. Estimating efficacy and drug ED50’s using von Frey thresholds: Impact of weber’s law and log transformation. J. Pain 2012, 13, 519–523. [Google Scholar] [CrossRef]
- Corey, S.M.; Vizzard, M.A.; Bouffard, N.A.; Badger, G.J.; Langevin, H.M. Stretching of the back improves gait, mechanical sensitivity and connective tissue inflammation in a rodent model. PLoS ONE 2012, 7, e29831. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Ahmadinia, K.; Li, X.; Hamilton, J.L.; Andrews, S.; Haralampus, C.A.; Xiao, G.; Sohn, H.M.; You, J.W.; Seo, Y.S.; et al. Development of an Experimental Animal Model for Lower Back Pain by Percutaneous Injury-Induced Lumbar Facet Joint Osteoarthritis. J. Cell Physiol. 2015, 230, 2837–2847. [Google Scholar] [CrossRef] [Green Version]
- Coggon, D.; Ntani, G.; Walker-Bone, K.; Palmer, K.T.; Felli, V.E.; Harari, R.; Barrero, L.H.; Felknor, S.A.; Gimeno, D.; Cattrell, A.; et al. Epidemiological Differences Between Localized and Nonlocalized Low Back Pain. Spine 2017, 42, 740–747. [Google Scholar] [CrossRef]
- Chenot, J.F.; Becker, A.; Leonhardt, C.; Keller, S.; Donner-Banzhoff, N.; Hildebrandt, J.; Basler, H.D.; Baum, E.; Kochen, M.M.; Pfingsten, M. Sex differences in presentation, course, and management of low back pain in primary care. Clin. J. Pain 2008, 24, 578–584. [Google Scholar] [CrossRef] [Green Version]
- La Porta, C.; Tappe-Theodor, A. Differential impact of psychological and psychophysical stress on low back pain in mice. Pain 2020, 161, 1442–1458. [Google Scholar] [CrossRef]
- Kuzawinska, O.; Lis, K.; Cudna, A.; Balkowiec-Iskra, E. Gender differences in the neurochemical response of trigeminal ganglion neurons to peripheral inflammation in mice. Acta. Neurobiol. Exp. 2014, 74, 227–232. [Google Scholar]
- Nazarian, A.; Tenayuca, J.M.; Almasarweh, F.; Armendariz, A.; Are, D. Sex differences in formalin-evoked primary afferent release of substance P. Eur. J. Pain 2014, 18, 39–46. [Google Scholar] [CrossRef]
- Vacca, V.; Marinelli, S.; Pieroni, L.; Urbani, A.; Luvisetto, S.; Pavone, F. Higher pain perception and lack of recovery from neuropathic pain in females: A behavioural, immunohistochemical, and proteomic investigation on sex-related differences in mice. Pain 2014, 155, 388–402. [Google Scholar] [CrossRef]
- Obeidat, A.M.; Donner, A.; Miller, R.E. An update on targets for treating osteoarthritis pain: NGF and TRPV1. Curr. Treat. Options Rheumatol. 2020, 6, 129–145. [Google Scholar] [CrossRef]
- Mai, L.; Huang, F.; Zhu, X.; He, H.; Fan, W. Role of Nerve Growth Factor in Orofacial Pain. J. Pain Res. 2020, 13, 1875–1882. [Google Scholar] [CrossRef]
- Malfait, A.M.; Miller, R.E.; Block, J.A. Targeting neurotrophic factors: Novel approaches to musculoskeletal pain. Pharmacol. Ther. 2020, 211, 107553. [Google Scholar] [CrossRef]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef] [Green Version]
- Lambiase, A.; Bracci-Laudiero, L.; Bonini, S.; Bonini, S.; Starace, G.; D’Elios, M.M.; De Carli, M.; Aloe, L. Human CD4+ T cell clones produce and release nerve growth factor and express high-affinity nerve growth factor receptors. J. Allergy Clin. Immunol. 1997, 100, 408–414. [Google Scholar] [CrossRef]
- Rost, B.; Hanf, G.; Ohnemus, U.; Otto-Knapp, R.; Groneberg, D.A.; Kunkel, G.; Noga, O. Monocytes of allergics and non-allergics produce, store and release the neurotrophins NGF, BDNF and NT-3. Regul. Pept. 2005, 124, 19–25. [Google Scholar] [CrossRef]
- Sessler, K.; Blechschmidt, V.; Hoheisel, U.; Mense, S.; Schirmer, L.; Treede, R.D. Spinal cord fractalkine (CX3CL1) signaling is critical for neuronal sensitization in experimental nonspecific, myofascial low back pain. J. Neurophysiol. 2021, 125, 1598–1611. [Google Scholar] [CrossRef]
- Sohrabji, F.; Miranda, R.C.; Toran-Allerand, C.D. Estrogen differentially regulates estrogen and nerve growth factor receptor mRNAs in adult sensory neurons. J. Neurosci. 1994, 14, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Berkley, K.J. Sex differences in pain. Behav. Brain Sci. 1997, 20, 371–380. [Google Scholar] [CrossRef]
- Dong, X.D.; Mann, M.K.; Kumar, U.; Svensson, P.; Arendt-Nielsen, L.; Hu, J.W.; Sessle, B.J.; Cairns, B.E. Sex-related differences in NMDA-evoked rat masseter muscle afferent discharge result from estrogen-mediated modulation of peripheral NMDA receptor activity. Neuroscience 2007, 146, 822–832. [Google Scholar] [CrossRef] [Green Version]
- Dina, O.A.; Aley, K.O.; Isenberg, W.; Messing, R.O.; Levine, J.D. Sex hormones regulate the contribution of PKC epsilon and PKA signalling in inflammatory pain in the rat. Eur. J. Neurosci. 2001, 13, 2227–2233. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Jin, L.; Zhao, Y.; Sui, M.Y.; Huang, L.; Tan, Y.X.; Chen, Y.K.; You, H.J. Sex-related differences in descending norepinephrine and serotonin controls of spinal withdrawal reflex during intramuscular saline induced muscle nociception in rats. Exp. Neurol. 2011, 228, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Loyd, D.R.; Murphy, A.Z. The role of the periaqueductal gray in the modulation of pain in males and females: Are the anatomy and physiology really that different? J. Neurosci. 2009, 2009, 462879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, J.; Holmes, M.; Coughlin, M. Endogenous NGF and nerve impulses regulate the collateral sprouting of sensory axons in the skin of the adult rat. J. Neurosci. 1992, 12, 1454–1466. [Google Scholar] [CrossRef]
- Xie, W.R.; Deng, H.; Li, H.; Bowen, T.L.; Strong, J.A.; Zhang, J.M. Robust increase of cutaneous sensitivity, cytokine production and sympathetic sprouting in rats with localized inflammatory irritation of the spinal ganglia. Neuroscience 2006, 142, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Diamond, J.; Coughlin, M.; Macintyre, L.; Holmes, M.; Visheau, B. Evidence that endogenous beta nerve growth factor is responsible for the collateral sprouting, but not the regeneration, of nociceptive axons in adult rats. Proc. Natl. Acad. Sci. USA 1987, 84, 6596–6600. [Google Scholar] [CrossRef] [Green Version]
- Krock, E.; Rosenzweig, D.H.; Chabot-Dore, A.J.; Jarzem, P.; Weber, M.H.; Ouellet, J.A.; Stone, L.S.; Haglund, L. Painful, degenerating intervertebral discs up-regulate neurite sprouting and CGRP through nociceptive factors. J. Cell Mol. Med. 2014, 18, 1213–1225. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syrett, M.; Reed, N.R.; Reed, W.R.; Richey, M.L.; Frolov, A.; Little, J.W. Sex-Related Pain Behavioral Differences following Unilateral NGF Injections in a Rat Model of Low Back Pain. Biology 2022, 11, 924. https://doi.org/10.3390/biology11060924
Syrett M, Reed NR, Reed WR, Richey ML, Frolov A, Little JW. Sex-Related Pain Behavioral Differences following Unilateral NGF Injections in a Rat Model of Low Back Pain. Biology. 2022; 11(6):924. https://doi.org/10.3390/biology11060924
Chicago/Turabian StyleSyrett, Michael, Nicholas R. Reed, William R. Reed, Madison L. Richey, Andrey Frolov, and Joshua W. Little. 2022. "Sex-Related Pain Behavioral Differences following Unilateral NGF Injections in a Rat Model of Low Back Pain" Biology 11, no. 6: 924. https://doi.org/10.3390/biology11060924