
Diversity and Metabolism of Microbial Communities in a Hypersaline Lake along a Geochemical Gradient

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction and Sequencing
2.2. Bioinformatic Analysis
2.3. Geochemical Methods
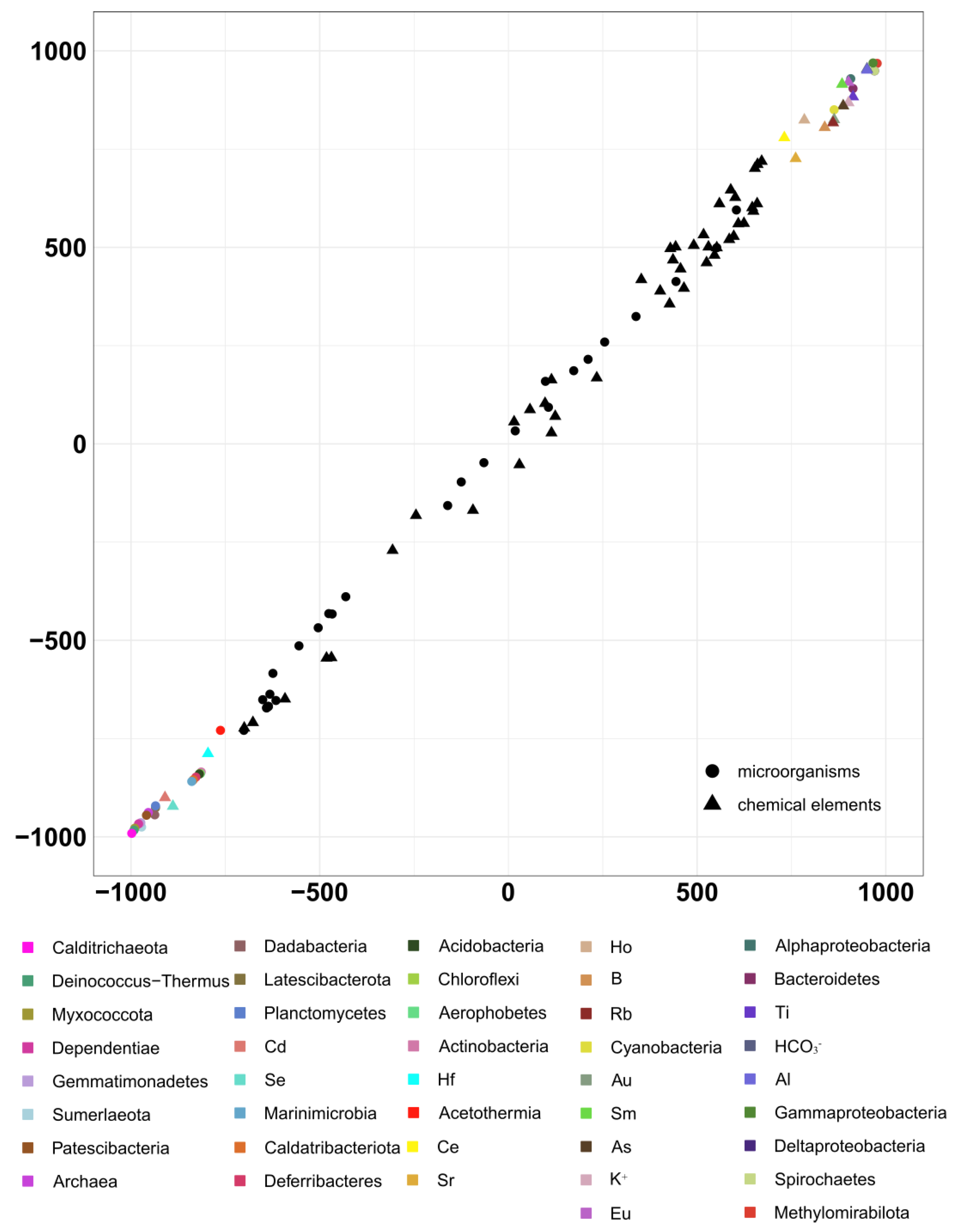
2.4. Statistical Analysis
3. Results
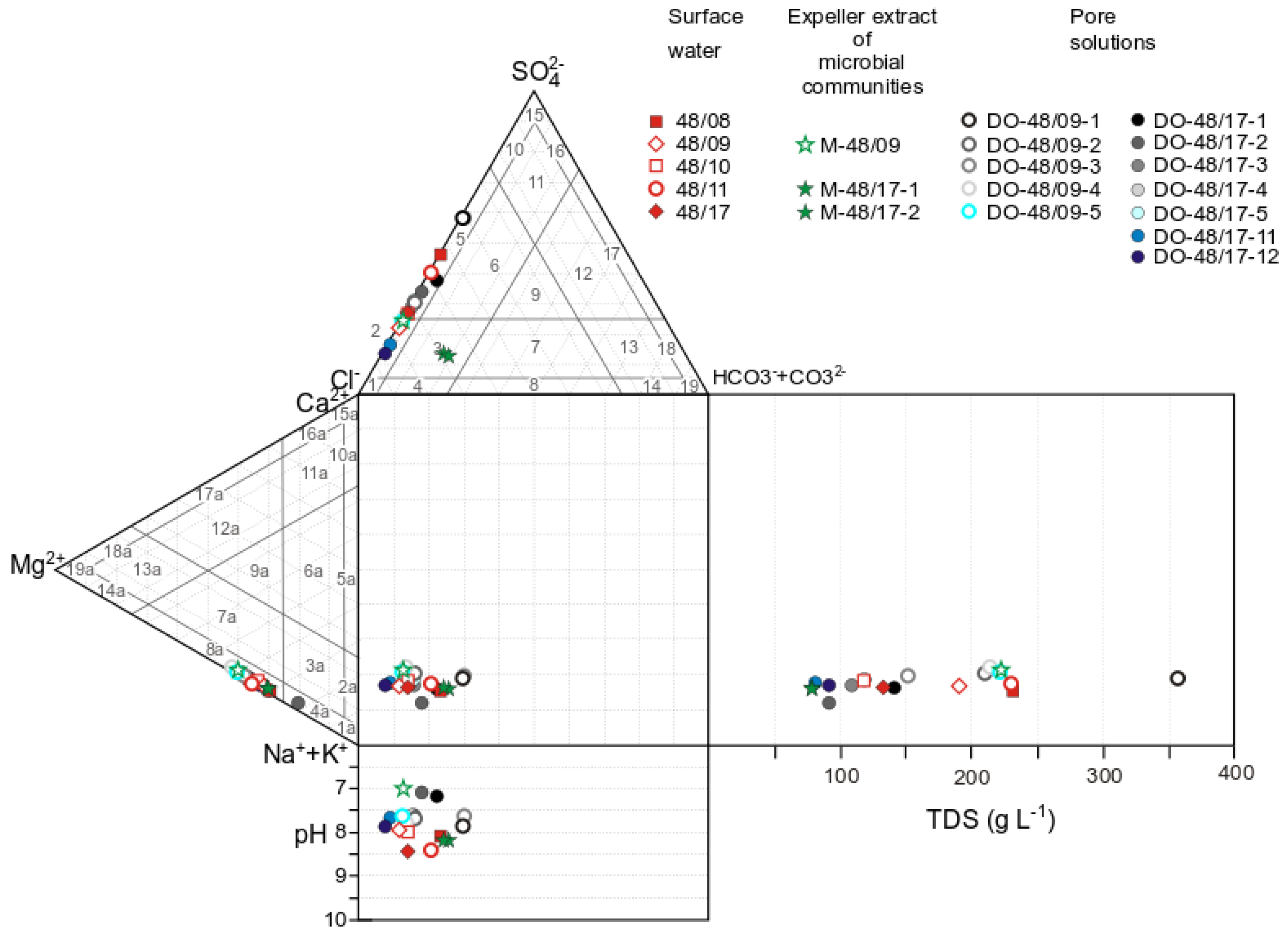
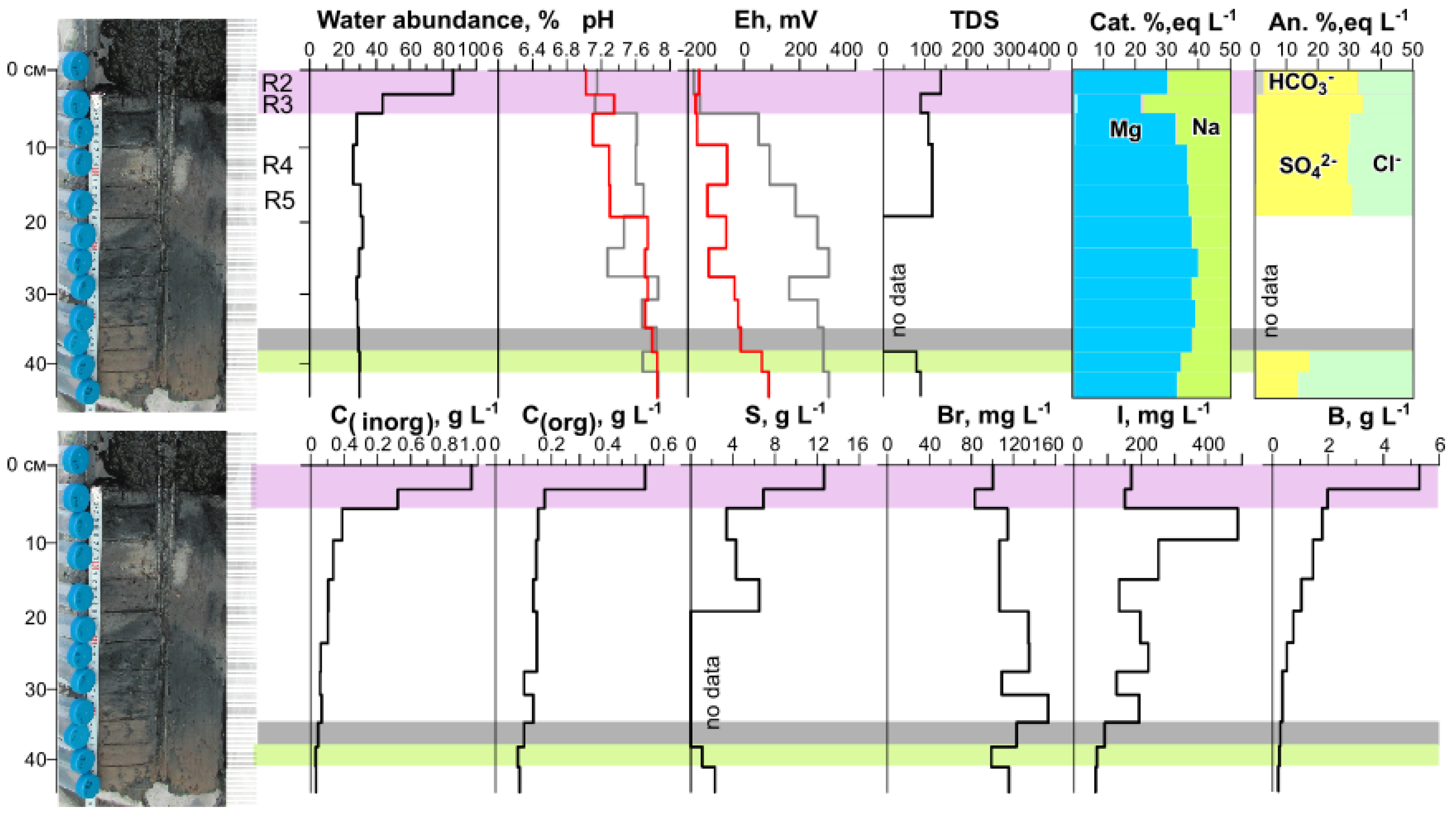
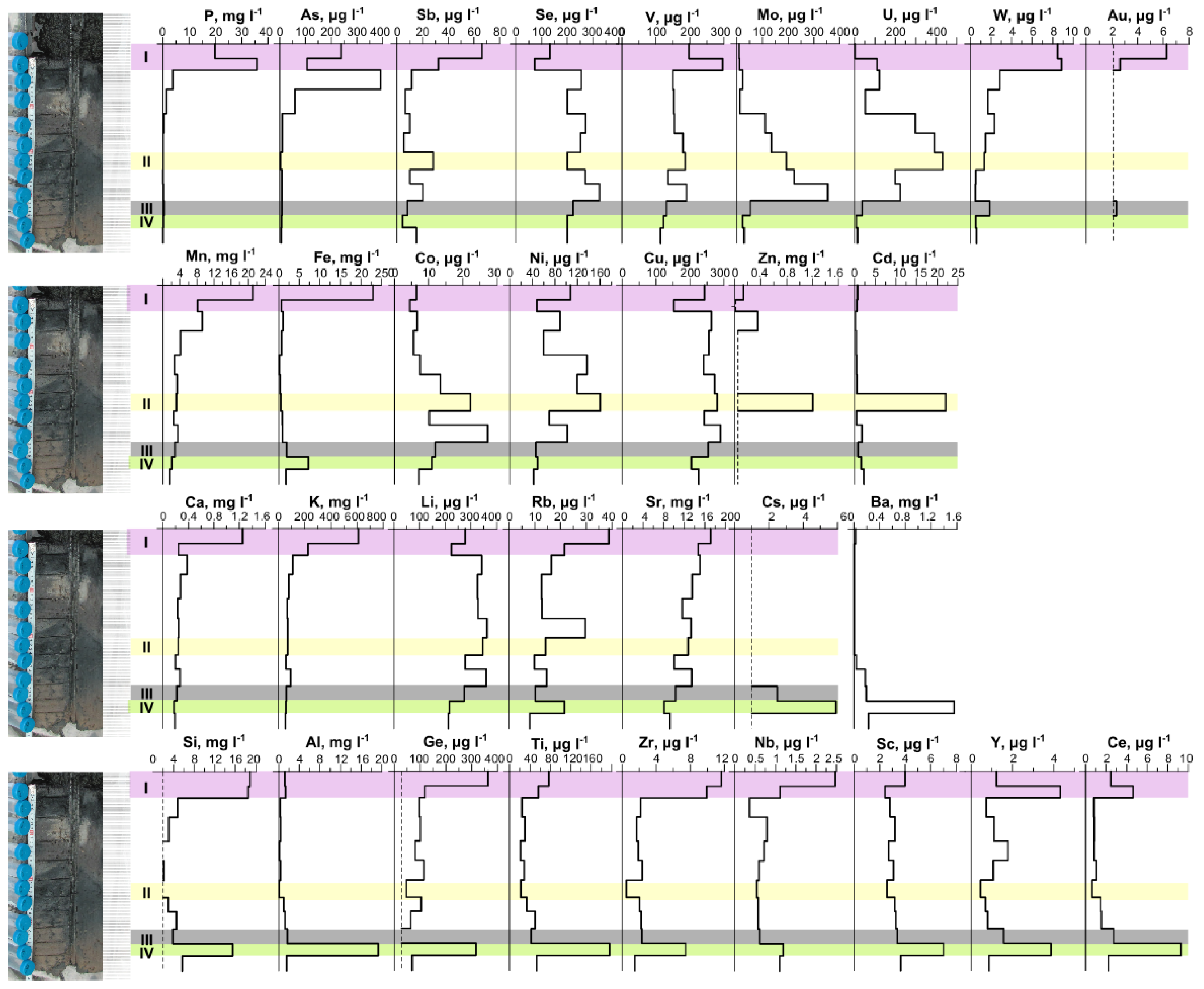
3.1. Physicochemical Parameters of Lake Solenoe (Water, Bottom Sediments, and Floating Microbial Communities)
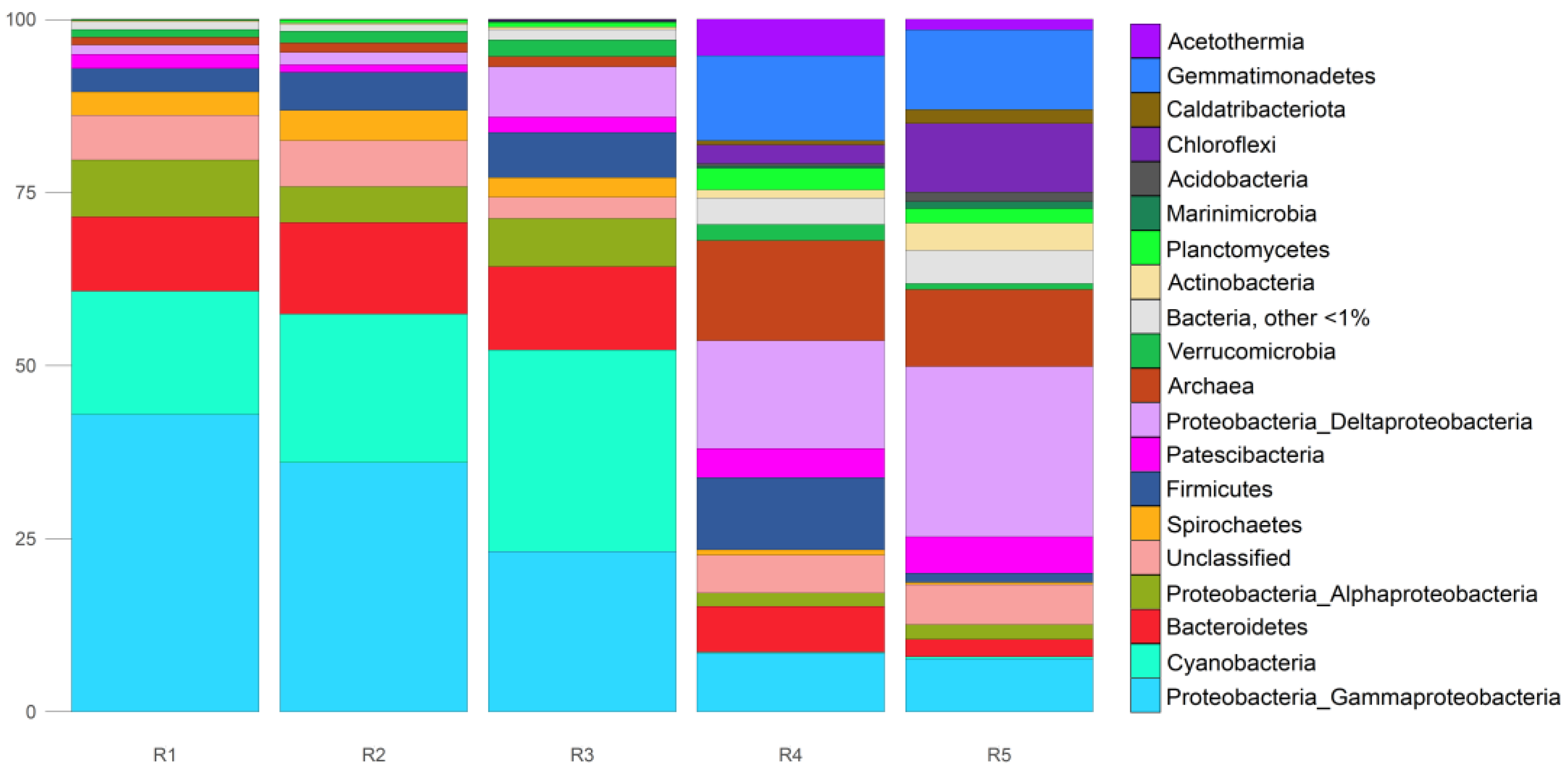
3.2. Composition and Structure of Microbial Communities. Metagenomic Results
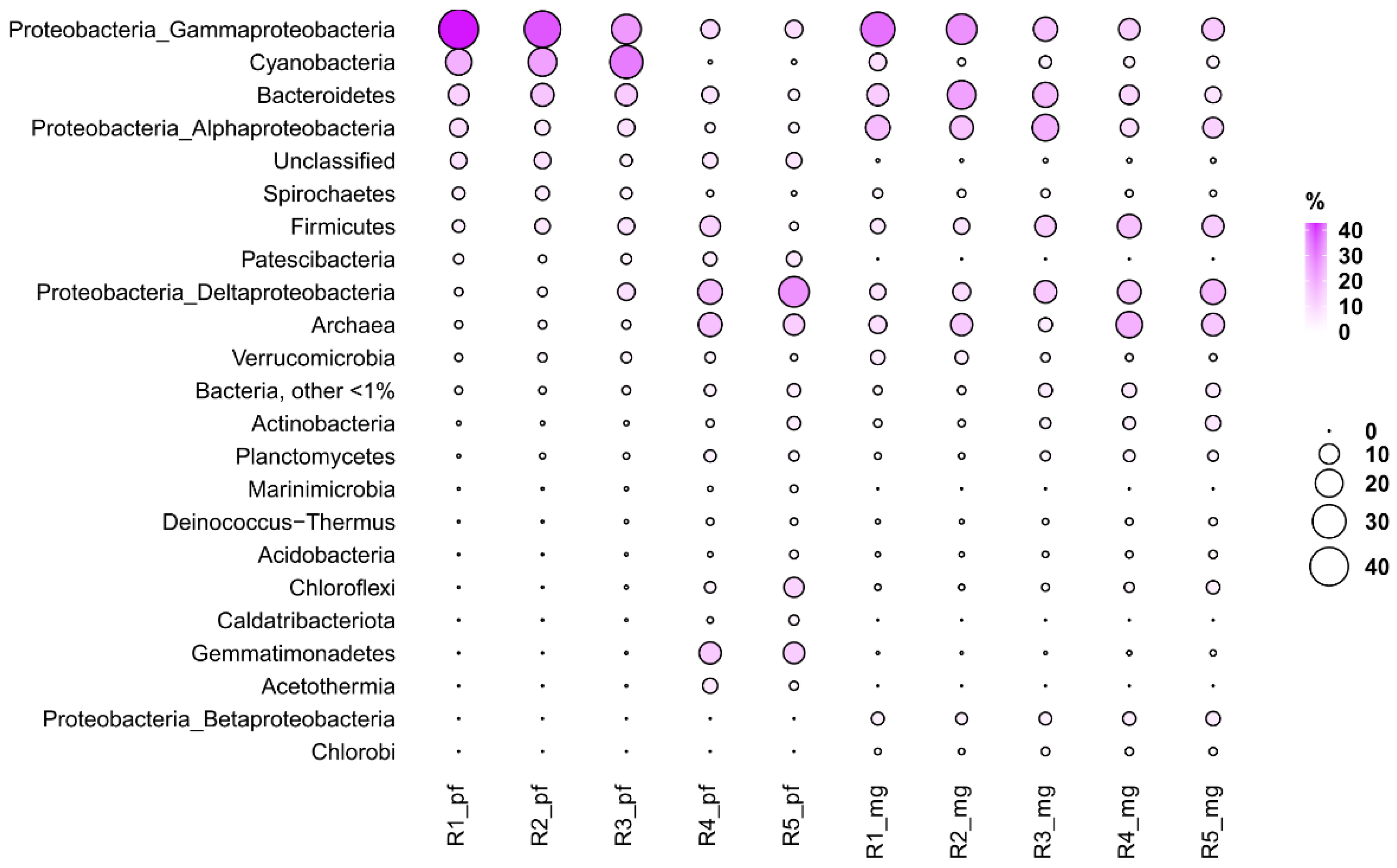
3.3. Taxonomic Profiling Based on 16S rRNA Sequences Obtained from Reads (phyloFlash)
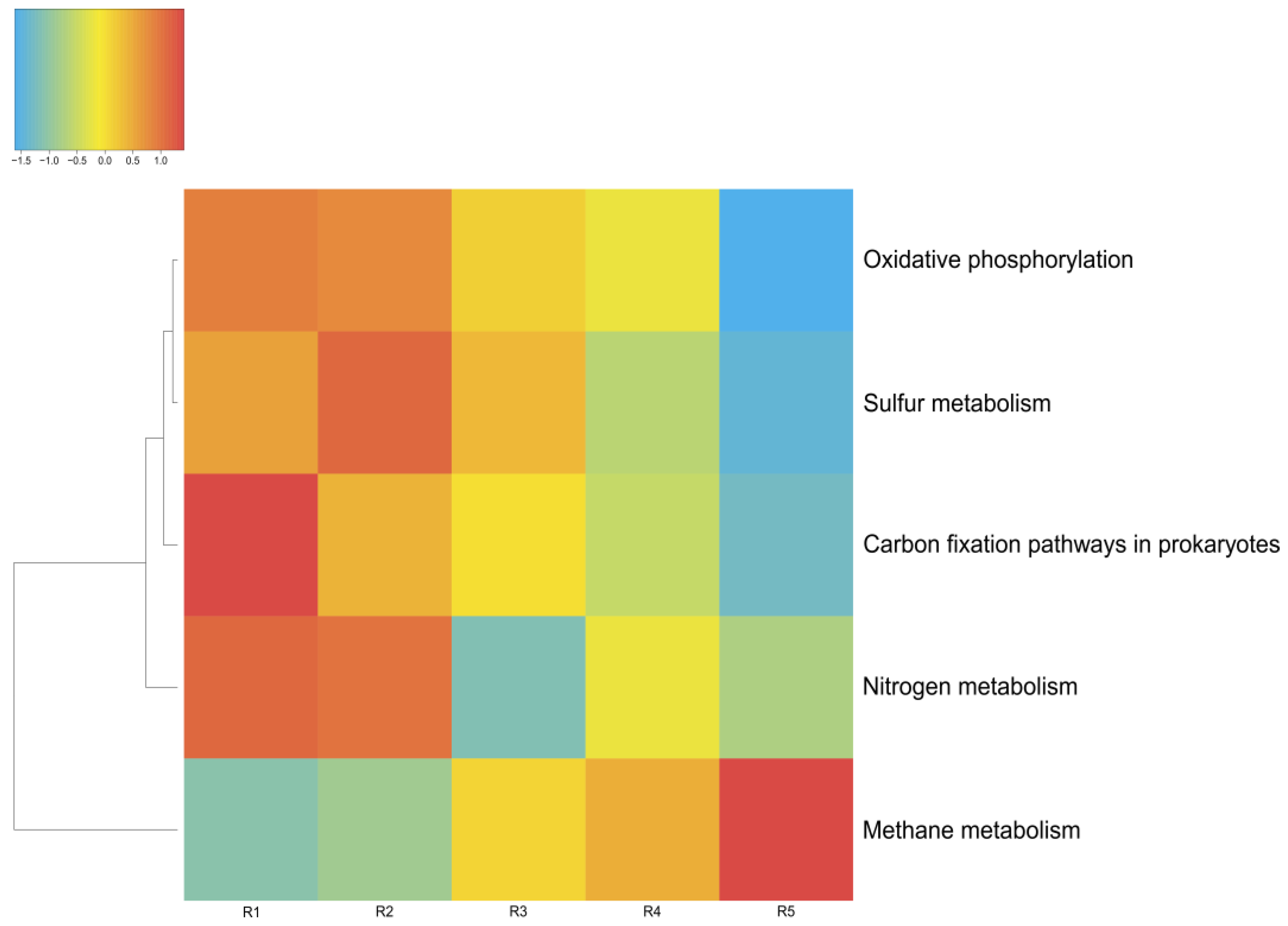
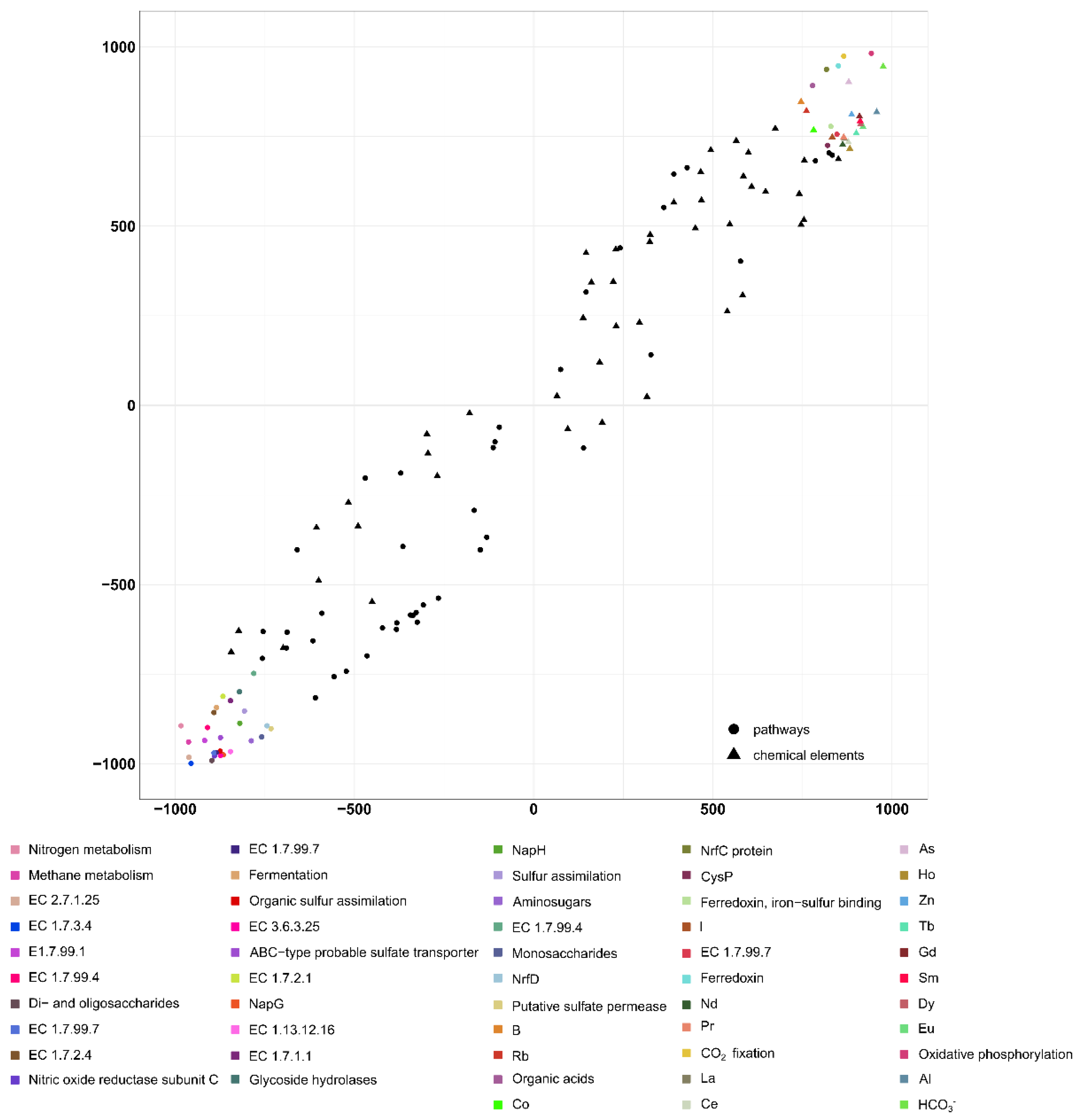
3.4. Metabolic Features of the Microbial Communities
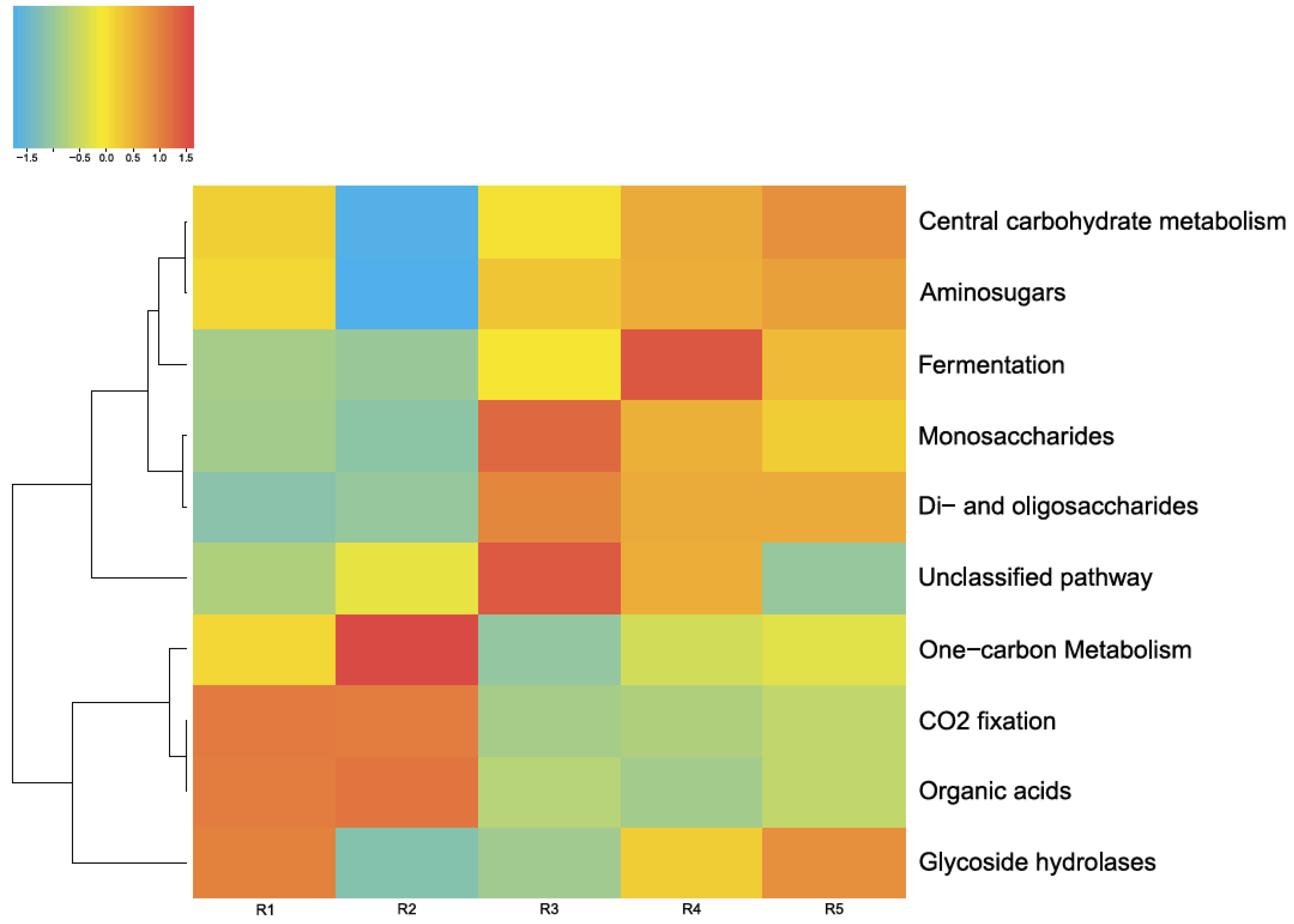
3.4.1. Carbon Metabolism
3.4.2. Nitrogen Metabolism
3.4.3. Sulfur Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horikoshi, K.; Grant, W.D. Extremophiles: Microbial Life in Extreme Environments; Wiley-Liss: Hoboken, NJ, USA, 1998; pp. 112–125. [Google Scholar]
- Oren, A. Industrial and Environmental Applications of Halophilic Microorganisms. Environ. Technol. 2010, 31, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, W.D.; Sorokin, D.Y. Distribution and Diversity of Soda Lake Alkaliphiles; Horikoshi, K., Antranikian, G., Bull, A., Robb, F., Stetter, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Tourova, T.P.; Grechnikova, M.A.; Kuznetsov, B.B.; Sorokin, D.Y. Phylogenetic Diversity of Bacteria in Soda Lake Stratified Sediments. Microbiology 2014, 83, 869–879. [Google Scholar] [CrossRef]
- Mirete, S.; Morgante, V.; González-Pastor, J.E. Functional Metagenomics of Extreme Environments. Curr. Opin. Biotechnol. 2016, 38, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Vavourakis, C.D.; Ghai, R.; Rodriguez-Valera, F.; Sorokin, D.Y.; Tringe, S.G.; Hugenholtz, P.; Muyzer, G. Metagenomic Insights into the Uncultured Diversity and Physiology of Microbes in Four Hypersaline Soda Lake Brines. Front. Microbiol. 2016, 7, 211. [Google Scholar] [CrossRef] [Green Version]
- Hug, L.A.; Baker, B.J.; Anantharaman, K.; Brown, C.T.; Probst, A.J.; Castelle, C.J.; Butterfield, C.N.; Hernsdorf, A.W.; Amano, Y.; Ise, K.; et al. A New View of the Tree of Life. Nat. Microbiol. 2016, 1, 16048. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A Communal Catalogue Reveals Earth’s Multiscale Microbial Diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Vavourakis, C.D.; Andrei, A.S.; Mehrshad, M.; Ghai, R.; Sorokin, D.Y.; Muyzer, G. A Metagenomics Roadmap to the Uncultured Genome Diversity in Hypersaline Soda Lake Sediments. Microbiome 2018, 6, 168. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Han, R.; Long, Q.; Gao, X.; Xing, J.; Shen, G.; Li, Y.; Wang, R. An Evaluation of the Core Bacterial Communities Associated with Hypersaline Environments in the Qaidam Basin, China. Arch. Microbiol. 2020, 202, 2093–2103. [Google Scholar] [CrossRef]
- Sorokin, D.Y. Anaerobic Haloalkaliphiles. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 1–16. [Google Scholar]
- Kompantseva, E.I.; Bryantseva, I.A.; Komova, A.V.; Namsaraev, B.B. The Structure of Phototrophic Communities of Soda Lakes of the Southeastern Transbaikal Region. Microbiology 2007, 76, 211–219. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial Diversity and Biogeochemical Cycling in Soda Lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [Green Version]
- Lazareva, E.V.; Bryanskaya, A.V.; Taran, O.P.; Kolmogorov, Y.P.; Malup, T.K.; Peltek, S.E.; Zhmodik, S.M. Investigation of Element Distribution between Components of a Salt-Lake System by SR-XRF. J. Surf. Investig. 2012, 6, 1009–1018. [Google Scholar] [CrossRef]
- Bryanskaya, A.V.; Malup, T.K.; Lazareva, E.V.; Taran, O.P.; Rozanov, A.S.; Efimov, V.M.; Peltek, S.E. The Role of Environmental Factors for the Composition of Microbial Communities of Saline Lakes in the Novosibirsk Region (Russia). BMC Microbiol. 2016, 16, S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryanskaya, A.V.; Shipova, A.A.; Rozanov, A.S.; Volkova, O.A.; Lazareva, E.V.; Uvarova, Y.E.; Goryachkovskaya, T.N.; Peltek, S.E. Metagenomics Dataset Used to Characterize Microbiome in Water and Sediments of the Lake Solenoe (Novosibirsk Region, Russia). Data Br. 2021, 34, 106709. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. MetaSPAdes: A New Versatile Metagenomic Assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [Green Version]
- Gruber-Vodicka, H.R.; Seah, B.K.B.; Pruesse, E. PhyloFlash: Rapid Small-Subunit RRNA Profiling and Targeted Assembly from Metagenomes. mSystems 2020, 5, e00920-20. [Google Scholar] [CrossRef]
- Meyer, F.; Paarmann, D.; D’Souza, M. The Metagenomics RAST Server—A Public Resource for the Automatic Phylogenetic and Functional Analysis of Metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Abdi, H. Partial Least Squares Regression and Projection on Latent Structure Regression (PLS Regression). Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 97–106. [Google Scholar] [CrossRef]
- Ryan, P.D.; Hammer, Ø.; Harper, D.A. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 543–551. [Google Scholar] [CrossRef]
- GOWER, J.C. Some Distance Properties of Latent Root and Vector Methods Used in Multivariate Analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Partial Least Squares Methods: Partial Least Squares Correlation and Partial Least Square Regression. Methods Mol. Biol. 2013, 930, 549–579. [Google Scholar] [CrossRef] [PubMed]
- Eugster, H.P.; Hardie, L.A. Saline Lakes. In Lakes; Springer: New York, NY, USA, 1978; pp. 237–293. [Google Scholar]
- McKee, K.L.; Mendelssohn, I.A.; Hester, M.W. Reexamination of Pore Water Sulfide Concentrations and Redox Potentials near the Aerial Roots of Rhizophora Mangle and Avicennia Germinans. Am. J. Bot. 1988, 75, 1352–1359. [Google Scholar] [CrossRef]
- Bychkov, A.Y. A Geochemical Model of Contemporary Ore Formation in the Uzon Caldera (Kamchatka); GEOS: Moscow, Russia, 2009. [Google Scholar]
- Brandt, K.K.; Vester, F.; Jensen, A.N.; Ingvorsen, K. Sulfate Reduction Dynamics and Enumeration of Sulfate-Reducing Bacteria in Hypersaline Sediments of the Great Salt Lake (Utah, USA). Microb. Ecol. 2001, 41, 1–11. [Google Scholar] [CrossRef]
- Klinkhammer, G.P.; Palmer, M.R. Uranium in the Oceans: Where It Goes and Why. Geochim. Cosmochim. Acta 1991, 55, 1799–1806. [Google Scholar] [CrossRef]
- Mangini, A.; Jung, M.; Laukenmann, S. What Do We Learn from Peaks of Uranium and of Manganese in Deep Sea Sediments? Mar. Geol. 2001, 177, 63–78. [Google Scholar] [CrossRef]
- Anderson, R.F.; Lehuray, A.P.; Fleisher, M.Q.; Murray, J.W. Uranium Deposition in Saanich Inlet Sediments, Vancouver Island. Geochim. Cosmochim. Acta 1989, 53, 2205–2213. [Google Scholar] [CrossRef]
- Lazareva, E.V.; Vosel, Y.S.; Bryanskaya, A.V.; Taran, O.P.; Chikunov, A.S.; Rozanov, A.S.; Peltek, S.E.; Zhmodik, S.M. Physicochemical Features of Pore Solutions of Bottom Sediments of Lake Solenoe (Novosibirsk Region). In Proceedings of the Geology of Seas and Oceans: Proceedings of XXII International Conference on Marine Geology, Moscow, Russia, 20–24 November 2017; pp. 112–116. [Google Scholar]
- Lovley, D.R. Dissimilatory Metal Reduction. Annu. Rev. Microbiol. 1993, 47, 263–290. [Google Scholar] [CrossRef]
- Lovley, D.R.; Roden, E.E.; Phillips, E.J.P.; Woodward, J.C. Enzymatic Iron and Uranium Reduction by Sulfate-Reducing Bacteria. Mar. Geol. 1993, 113, 41–53. [Google Scholar] [CrossRef]
- Vosel, Y.; Vosel, S.; Melgunov, M.; Lazareva, E.; Kropacheva, M.; Strakhovenko, V. Discussions on the Driving Mechanism of Postdepositional Migration of 241Am and 137Cs in Organomineral Sediments (Lake Krugloe, Tomsk Region, Russia). Environ. Sci. Pollut. Res. 2019, 26, 19180–19188. [Google Scholar] [CrossRef] [PubMed]
- Liger, E.; Charlet, L.; Van Cappellen, P. Surface Catalysis of Uranium(VI) Reduction by Iron(II). Geochim. Cosmochim. Acta 1999, 63, 2939–2955. [Google Scholar] [CrossRef]
- Southam, G.; Saunders, J.A. The Geomicrobiology of Ore Deposits. Econ. Geol. 2005, 100, 1067–1084. [Google Scholar] [CrossRef]
- Pokrovsky, O.S.; Schott, J.; Dupré, B. Trace Element Fractionation and Transport in Boreal Rivers and Soil Porewaters of Permafrost-Dominated Basaltic Terrain in Central Siberia. Geochim. Cosmochim. Acta 2006, 70, 3239–3260. [Google Scholar] [CrossRef]
- Pourret, O.; Davranche, M.; Gruau, G.; Dia, A. Rare Earth Elements Complexation with Humic Acid. Chem. Geol. 2007, 243, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Davranche, M.; Pourret, O.; Gruau, G.; Dia, A.; Jin, D.; Gaertner, D. Competitive Binding of REE to Humic Acid and Manganese Oxide: Impact of Reaction Kinetics on Development of Cerium Anomaly and REE Adsorption. Chem. Geol. 2008, 247, 154–170. [Google Scholar] [CrossRef]
- Bräuer, S.L.; Basiliko, N.; Siljanen, H.M.P.; Zinder, S.H. Methanogenic Archaea in Peatlands. FEMS Microbiol. Lett. 2020, 367, fnaa172. [Google Scholar] [CrossRef]
- Dmitrieva, O.M.; Barkhutova, D.D. Characteristics of Organic Matter Composition in Microbial Mats of Alkaline Water Ecosystems of the Baikal Region. Mod. Probl. Sci. Educ. 2017, 3, 154. [Google Scholar]
- DelSontro, T.; Beaulieu, J.J.; Downing, J.A. Greenhouse Gas Emissions from Lakes and Impoundments: Upscaling in the Face of Global Change. Limnol. Oceanogr. Lett. 2018, 3, 64–75. [Google Scholar] [CrossRef]
- Antony, C.P.; Kumaresan, D.; Hunger, S.; Drake, H.L.; Murrell, J.C.; Shouche, Y.S. Microbiology of Lonar Lake and Other Soda Lakes. ISME J. 2013, 7, 468–476. [Google Scholar] [CrossRef]
- Schneider, J.; Campion-Alsumard, T. Le Construction and Destruction of Carbonates by Marine and Freshwater Cyanobacteria. Eur. J. Phycol. 1999, 34, 417–426. [Google Scholar] [CrossRef]
- Vavourakis, C.D.; Mehrshad, M.; Balkema, C.; Van Hall, R.; Andrei, A.Ş.; Ghai, R.; Sorokin, D.Y.; Muyzer, G. Metagenomes and Metatranscriptomes Shed New Light on the Microbial-Mediated Sulfur Cycle in a Siberian Soda Lake. BMC Biol. 2019, 17, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carini, S.; Bano, N.; LeCleir, G.; Joye, S.B. Aerobic Methane Oxidation and Methanotroph Community Composition during Seasonal Stratification in Mono Lake, California (USA). Environ. Microbiol. 2005, 7, 1127–1138. [Google Scholar] [CrossRef]
- Kaluzhnaya, M.; Khmelenina, V.; Eshinimaev, B.; Suzina, N.; Nikitin, D.; Solonin, A.; Lin, J.L.; McDonald, I.; Murrell, C.; Trotsenko, Y. Taxonomic Characterization of New Alkaliphilic and Alkalitolerant Methanotrophs from Soda Lakes of the Southeastern Transbaikal Region and Description of Methylomicrobium Buryatense Sp.Nov. Syst. Appl. Microbiol. 2001, 24, 166–176. [Google Scholar] [CrossRef]
- Ljungdahl, L.; Irion, E.; Wood, H.G. Total Synthesis of Acetate from CO2. I. Co-Methylcobyric Acid and CO-(Methyl)-5-Methoxybenzimidazolylcobamide as Intermediates with Clostridium Thermoaceticum. Biochemistry 1965, 4, 2771–2780. [Google Scholar] [CrossRef] [PubMed]
- Berg, I.A. Ecological Aspects of the Distribution of Different Autotrophic CO2 Fixation Pathways. Appl. Environ. Microbiol. 2011, 77, 1925–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, G. Alternative Pathways of Carbon Dioxide Fixation: Insights into the Early Evolution of Life? Annu. Rev. Microbiol. 2011, 65, 631–658. [Google Scholar] [CrossRef]
- Jansen, K.; Stupperich, E.; Fuchs, G. Carbohydrate Synthesis from Acetyl CoA in the Autotroph Methanobacterium Thermoautotrophicum. Arch. Microbiol. 1982, 132, 355–364. [Google Scholar] [CrossRef]
- Matschiavelli, N.; Oelgeschläger, E.; Cocchiararo, B.; Finke, J.; Rother, M. Function and Regulation of Isoforms of Carbon Monoxide Dehydrogenase/Acetyl Coenzyme A Synthase in Methanosarcina Acetivorans. J. Bacteriol. 2012, 194, 5377–5387. [Google Scholar] [CrossRef] [Green Version]
- Zarzycki, J.; Brecht, V.; Müller, M.; Fuchs, G. Identifying the Missing Steps of the Autotrophic 3-Hydroxypropionate CO2 Fixation Cycle in Chloroflexus Aurantiacus. Proc. Natl. Acad. Sci. USA 2009, 106, 21317–21322. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domain | Layer | ||||
|---|---|---|---|---|---|
| R1 | R2 | R3 | R4 | R5 | |
| Archaea | 1.08 | 1.32 | 1.43 | 14.17 | 10.72 |
| Bacteria | 97.19 | 97.25 | 96.48 | 84.67 | 87.28 |
| Eukaryota | 1.73 | 1.43 | 2.09 | 1.16 | 2.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryanskaya, A.V.; Shipova, A.A.; Rozanov, A.S.; Kolpakova, O.A.; Lazareva, E.V.; Uvarova, Y.E.; Efimov, V.M.; Zhmodik, S.M.; Taran, O.P.; Goryachkovskaya, T.N.; et al. Diversity and Metabolism of Microbial Communities in a Hypersaline Lake along a Geochemical Gradient. Biology 2022, 11, 605. https://doi.org/10.3390/biology11040605
Bryanskaya AV, Shipova AA, Rozanov AS, Kolpakova OA, Lazareva EV, Uvarova YE, Efimov VM, Zhmodik SM, Taran OP, Goryachkovskaya TN, et al. Diversity and Metabolism of Microbial Communities in a Hypersaline Lake along a Geochemical Gradient. Biology. 2022; 11(4):605. https://doi.org/10.3390/biology11040605
Chicago/Turabian StyleBryanskaya, Alla V., Aleksandra A. Shipova, Alexei S. Rozanov, Oxana A. Kolpakova, Elena V. Lazareva, Yulia E. Uvarova, Vadim M. Efimov, Sergey M. Zhmodik, Oxana P. Taran, Tatyana N. Goryachkovskaya, and et al. 2022. "Diversity and Metabolism of Microbial Communities in a Hypersaline Lake along a Geochemical Gradient" Biology 11, no. 4: 605. https://doi.org/10.3390/biology11040605