
Chronic Stress-Related Gastroenteric Pathology in Cheetah: Relation between Intrinsic and Extrinsic Factors

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
Literature Search Strategy and Study Selection
3. Results
4. Discussion
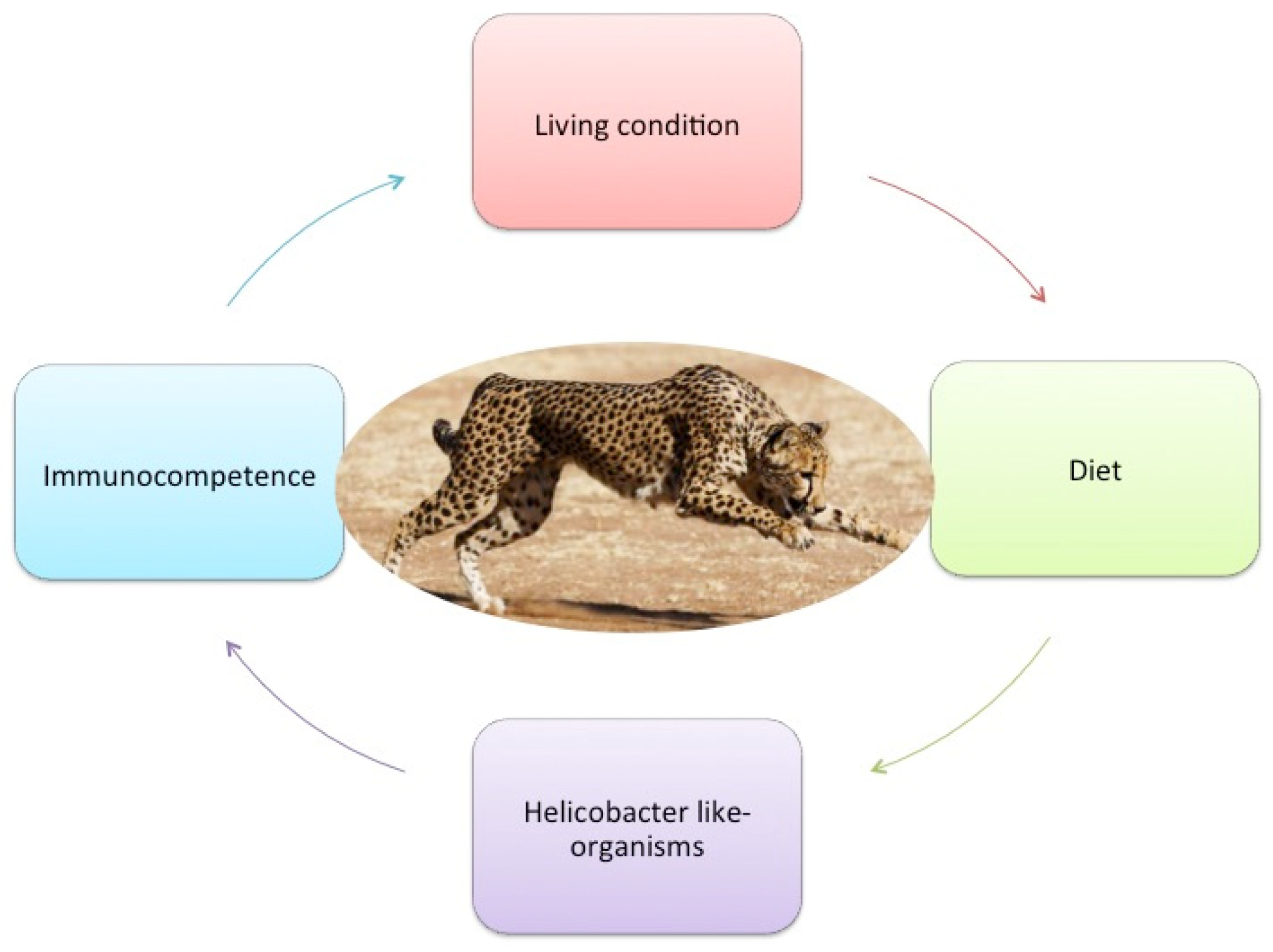
4.1. Extrinsic Factors
4.1.1. Captive Living Conditions
4.1.2. Diet
4.2. Intrinsic Factors
4.2.1. Microbiome and Gastric Helicobacter-like Organisms
4.2.2. Immunocompetence and Lack of Genetic Variation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Durant, S.M.; Mitchell, N.; Groom, R.; Pettorelli, N.; Ipavec, A.; Jacobson, A.P.; Woodroffe, R.; Bohm, M.; Hunter, L.T.; Becker, M.S.; et al. The global decline of cheetah Acinonyx jubatus and what it means for conservation. Proc. Natl. Acad. Sci. USA 2017, 114, 528–533. [Google Scholar] [CrossRef] [Green Version]
- Munson, L. Diseases of captive cheetahs (Acinonyx jubatus): Results of the cheetah research council pathology survey, 1989–1992. Zoo Biol. 1993, 12, 105–124. [Google Scholar] [CrossRef]
- Bolton, L.A.; Munson, L. Glomerulosclerosis in captive cheetahs (Acinonyx jubatus). Vet. Pathol. 1999, 36, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Eaton, K.A.; Radin, M.J.; Kramer, L.; Wack, R.; Sherding, R.; Krakowka, S.; Fox, J.G.; Morgan, D.R. Epizootic Gastritis Associated with Gastric Spiral Bacilli in Cheetahs (Acinonyx jubatus). Vet. Pathol. 1993, 30, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Terio, K.A.; Munson, L.; Marker, L.; Aldridge, B.M.; Solnick, J.V. Comparison of Helicobacter spp. in Cheetahs (Acinonyx jubatus) with and without Gastritis. J. Clin. Microbiol. 2005, 43, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terio, K.A.; Munson, L.; Moore, P.F. Characterization of the gastric immune response in cheetahs (Acinonyx jubatus) with Helicobacter-associated gastritis. Vet. Pathol. 2012, 49, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, G.; Gambi, R.; Uncini, R.; Piccinini, R.; Berardi, S.; Pengo, G.; Bassotti, G.; Cerquetella, M. Severe gastritis with double Helicobacter spp. infection associated with Barrett’s esophagus in a cheetah. Helicobacter 2014, 19, 462–464. [Google Scholar] [CrossRef]
- Lobetti, R.; Picard, J.; Kriek, N.; Rogers, P. Prevalence of helicobacteriosis and gastritis in semicaptive cheetahs (Acinonyx jubatus). J. Zoo Wildl. Med. 1999, 30, 492–496. [Google Scholar]
- Whitehouse-Tedd, K.M.; Lefebvre, S.L.; Janssens, G.P. Dietary Factors Associated with Faecal Consistency and Other Indicators of Gastrointestinal Health in the Captive Cheetah (Acinonyx jubatus). PLoS ONE 2015, 10, e0120903. [Google Scholar] [CrossRef]
- Papendick, R.E.; Munson, L.; O’Brien, T.D.; Johnson, K.H. Systemic AA amyloidosis in captive cheetahs (Acinonyx jubatus). Vet. Pathol. 1997, 34, 549–556. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, S.; Chambers, J.K.; Une, Y. Beta Amyloid Deposition and Neurofibrillary Tangles Spontaneously Occur in the Brains of Captive Cheetahs (Acinonyx jubatus). Vet. Pathol. 2012, 49, 304–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elenkov, I.J.; Chrousos, G.P. Stress Hormones, Th1/Th2 patterns, Pro/Anti-inflammatory Cytokines and Susceptibility to Disease. Trends Endocrinol. Metab. 1999, 10, 359–368. [Google Scholar] [CrossRef]
- Terio, K.A.; Marker, L.; Munson, L. Evidence for chronic stress in captive but not free-ranging cheetahs (Acinonyx jubatus) based on adrenal morphology and function. J. Wildl. Dis. 2004, 40, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, K.A.; Radin, M.J.; Kramer, L.; Wack, R.; Sherding, R.; Krakowka, S.; Morgan, D.R. Gastric spiral bacilli in captive cheetahs. Scand. J. Gastroenterol. Suppl. 1991, 181, 38–42. [Google Scholar] [CrossRef]
- Dailidiene, D.; Dailide, G.; Ogura, K.; Zhang, M.; Mukhopadhyay, A.K.; Eaton, K.A.; Cattoli, G.; Kusters, J.G.; Berg, D.E. Helicobacter acinonychis: Genetic and rodent infection studies of a Helicobacter pylori-like gastric pathogen of cheetahs and other big cats. J. Bacteriol. 2004, 186, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Lane, E.P.; Miller, S.; Lobetti, R.; Caldwell, P.; Bertschinger, H.J.; Burroughs, R.; Kotze, A.; van Dyk, A. Effect of Diet on the Incidence of and Mortality Owing to Gastritis and Renal Disease in Captive Cheetahs (Acinonyx jubatus) in South Africa. Zoo Biol. 2012, 31, 669–682. [Google Scholar] [CrossRef]
- Marker, L.; Grisham, J.; Brewer, B. A Brief History of Cheetah Conservation. Cheetahs Biol. Conserv. 2018, 3–16. [Google Scholar] [CrossRef]
- Kitchener, A.C.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae: The final report of the Cat Classification Task Force of the IUCN Cat Specialist Group. Cat News 2017. Available online: https://repository.si.edu/handle/10088/32616 (accessed on 16 February 2022).
- Marker, L.; Johnson, B. Studbook 2019; Cheetah Conservation Fund: Otjiwarongo, Namibia, 2019. [Google Scholar]
- Munson, L.; Terio, K.A.; Worley, M.; Jago, M.; Bagot-Smith, A.; Marker, L. Extrinsic factors significantly affect patterns of disease in free-ranging and captive cheetah (Acinonyx jubatus) populations. J. Wildl. Dis. 2005, 41, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Bentkowski, P.; Radwan, J. Evolution of major histocompatibility complex gene copy number. PLoS Comput. Biol. 2019, 15, e1007015. [Google Scholar] [CrossRef]
- Prost, S.; Machado, A.P.; Zumbroich, J.; Preier, L.; Mahtani-Williams, S.; Guschanski, K.; Brealey, J.C.; Fernandes, C.; Vercammen, P.; Godsall-Bottriell, L.; et al. Conservation Genomic Analyses of African and Asiatic Cheetahs (Acinonyx jubatus) Across Their Current and Historic Species Range. BioRxiv 2020. preprint. [Google Scholar] [CrossRef] [Green Version]
- Castro-Prieto, A.; Wachter, B.; Sommer, S. Cheetah paradigm revisited: MHC diversity in the world’s largest free-ranging population. Mol. Biol. Evol. 2011, 28, 1455–1468. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Blas, J.; Bortolotti, G.R.; Tella, J.L.; Baos, R.; Marchant, T.A. Stress response during development predicts fitness in a wild, long lived vertebrate. Proc. Natl. Acad. Sci. USA 2007, 104, 8880–8884. [Google Scholar] [CrossRef] [Green Version]
- Terio, K.A.; Mitchell, E.; Walzer, C.; Schmidt-Küntzel, A.; Marker, L.; Citino, S. Diseases Impacting Captive and Free-Ranging Cheetahs. Cheetahs Biol. Conserv. 2018, 349–364. [Google Scholar] [CrossRef]
- Wells, A.; Terio, K.; Ziccardi, M.; Munson, L. The stress response to environmental change in captive cheetahs (Acionyx jubatus). J. Zoo Wildl. Med. 2004, 35, 8–14. [Google Scholar] [CrossRef]
- Terio, K.A.; Citino, S.B.; Brown, J.L. Fecal cortisol metabolite analysis for noninvasive monitoring of adrenocortical function in the cheetah (Acinonyx jubatus). J. Zoo Wildl. Med. 1999, 30, 484–491. [Google Scholar]
- Gillis-Germitsch, N.; Vybiral, P.R.; Codron, D.; Clauss, M.; Kotze, A.; Mitchell, E.P. Intrinsic factors, adrenal gland morphology, and disease burden in captive cheetahs (Acinonyx jubatus) in South Africa. Zoo Biol. 2017, 36, 40–49. [Google Scholar] [CrossRef]
- Uetake, K.; Une, Y.; Ito, S.; Yamabe, M.; Toyoda, H.; Tanaka, T. Relationship of climatic conditions to fecal corticosterone levels of captive cheetahs reared in Japan. Anim. Sci. J. 2014, 85, 915–918. [Google Scholar] [CrossRef]
- Siddiqui, A.H.; Siddiqui, F. Curling Ulcer (Stress-induced Gastric). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Choi, Y.H.; Lee, J.H.; Shin, J.J.; Cho, Y.S. A revised risk analysis of stress ulcers in burn patients receiving ulcer prophylaxis. Clin. Exp. Emerg. Med. 2015, 2, 250–255. [Google Scholar] [CrossRef]
- Gibbons, A.H.; Legon, S.; Walker, M.M.; Ghatei, M.; Calam, J. The effect of gastrin-releasing peptide on gastrin and somatostatin messenger RNAs in humans infected with Helicobacter pylori. Gastroenterology 1997, 112, 1940–1947. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y.; Go, M.F.; Lew, G.M.; Genta, R.M.; Rehfeld, J.F. Helicobacter pylori infection and exaggerated gastrin release. Effects of inflammation and progastrin processing. Scand. J. Gastroenterol. 1993, 28, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Pengo, G.; Renzoni, G.; Taccini, E. Effect of Helicobacter spp. infection on G and D cells of canine gastric mucosa. In Proceedings of the 18th Meeting of European Society of Veterinary Pathology, Amsterdam, The Netherlands, 19–22 September 2000; Volume 18, p. 157, ISBN 9788361602576. [Google Scholar]
- Sun, F.P.; Song, Y.G.; Cheng, W.; Zhao, T.; Yao, Y.L. Gastrin, somatostatin, G and D cells of gastric ulcer in rats. World J. Gastroenterol. 2002, 8, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Tham, T.C.; Chen, L.; Dennison, N.; Johnston, C.F.; Collins, J.S.; Ardill, J.E.; Buchanan, K.D. Effect of Helicobacter pylori eradication on antral somatostatin cell density in humans. Eur. J. Gastroenterol. Hepatol. 1998, 10, 289–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Vosmaer, G.D.; Tytgat, G.N.; Xiao, S.D.; Ten Kate, F.J. Gastrin (G) cells and somatostatin (D) cells in patients with dyspeptic symptoms: Helicobacter pylori associated and non-associated gastritis. J. Clin. Pathol. 2005, 58, 927–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Megha, R.; Lopez, P.P. Stress-induced gastritis. In StatPearls; [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Kwiecien, S.; Ptak-Belowska, A.; Krzysiek-Maczka, G.; Targosz, A.; Jasnos, K.; Magierowski, M.; Szczyrk, U.; Brzozowski, B.; Konturek, S.J.; Konturek, P.C.; et al. Asymmetric dimethylarginine, an endogenous inhibitor of nitric oxide synthase, interacts with gastric oxidative metabolism and enhances stress-induced gastric lesions. J. Physiol. Pharmacol. 2012, 63, 515–524. [Google Scholar]
- Rossi, G.; Rossi, M.; Vitali, C.G.; Fortuna, D.; Burroni, D.; Pancotto, L.; Capecchi, S.; Sozzi, S.; Renzoni, G.; Braca, G.; et al. A conventional beagle dog model for acute and chronic infection with Helicobacter pylori. Infect. Immun. 1999, 67, 3112–3120. [Google Scholar] [CrossRef] [Green Version]
- Calam, J.; Gibbons, A.; Healey, Z.V.; Bliss, P.; Arebi, N. How does Helicobacter pylori cause mucosal damage? Its effect on acid and gastrin physiology. Gastroenterology 1997, 113 (Suppl. S6), S43–S49, discussion S50. [Google Scholar] [CrossRef]
- Ewer, R.F. The Carnivores; Cornell University Press: Ithaca, NY, USA, 1973; p. 544. [Google Scholar]
- Kerr, K.R.; Morris, C.L.; Burke, S.L.; Swanson, K.S. Influence of Dietary Fiber Type and Amount on Enery and Nutritient Digestibility, Fecal Characteristics, And Fecal Fermentative End-Product Concentrations in Captive Exotic Felids Fed a Raw Beef-Based Diet. J. Anim. Sci. 2013, 91, 2199–2210. [Google Scholar] [CrossRef]
- Verbrugghe, A.; Bakovic, M. Peculiarities of One-Carbon Metabolism in the Strict Carnivorous Cat and the Role in Feline Hepatic Lipidosis. Nutrients 2013, 5, 2811–2835. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.E. Essential fatty acid metabolism in dogs and cats. R. Bras. Zootec. 2008, 37, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.E. Fatty acid metabolism in domestic cats (Felis catus) and cheetahs (Acinonyx jubatas). Proc. Nutr. Soc. 1997, 56, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, B.C.; Cantrill, R.C.; Varaday, D. The reversal of essential fatty acid deficiency symptoms in the cheetah. S. Afr. J. Zool. 1986, 21, 161–164. [Google Scholar] [CrossRef]
- Depauw, S.; Hesta, M.; Whitehouse-Tedd, K.; Vanhaecke, L.; Verbrugghe, A.; Janssens, G.P. Animal fibre: The forgotten nutrient in strict carnivores? Firstinsights in the cheetah. J. Anim. Physiol. Anim. Nutr. 2013, 97, 146–154. [Google Scholar] [CrossRef]
- Igene, J.O.; Pearson, A.M.; Gray, L.I. Effects of length of frozen storage, cooking and holding temperatures upon component phospholipids and the fatty acid composition of meat triglycerides and phospholipids. Food Chem. 1981, 7, 289–303. [Google Scholar] [CrossRef]
- Voigt, C.C.; Thalwitzer, S.; Melzheimer, J.; Blanc, A.S.; Jago, M.; Wachter, B. The Conflict between Cheetahs and Humans on Namibian Farmland Elucidated by Stable Isotope Diet Analysis. PLoS ONE 2014, 9, e101917. [Google Scholar] [CrossRef]
- Calder, P.C. n−3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83 (Suppl. S6), 1505S–1519S. [Google Scholar] [CrossRef]
- James, M.J.; Gibson, R.A.; Cleland, L.G. Dietary polyunsaturated fatty acids and inflammatory mediator production. Am. J. Clin. Nutr. 2000, 71 (Suppl. S1), 343S–348S. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The Importance of the Ratio of Omega-6/Omega-3 Essential Fatty Acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Thompson, L.; Cockayne, A.; Spiller, R.C. Inhibitory effect of polyunsaturated fatty acids on the growth of Helicobacter pylori: A possible explanation of the effect of diet on peptic ulceration. Gut 1994, 35, 1557–1561. [Google Scholar] [CrossRef]
- Mohammed, A.; Janakiram, N.B.; Brewer, M.; Duff, A.; Lightfoot, S.; Brush, R.S.; Anderson, R.E.; Rao, C.V. Endogenous n-3 polyunsaturated fatty acids delay progression of pancreatic ductal adenocarcinoma in Fat-1-p48Cre/+-LSL-KrasG12D/+ mice. Neoplasia 2012, 14, 1249–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjari, V.; Das, U.N. Effect of polyunsaturated fatty acids on dexamethasone-induced gastric mucosal damage. Prostaglandins Leukot Essent Fat. Acids 2000, 62, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Guarner, F. Enteric Flora in Health and Disease. Digestion 2006, 73 (Suppl. S1), 5–12. [Google Scholar] [CrossRef] [PubMed]
- Wasimuddin; Menke, S.; Melzheimer, J.; Thalwitzer, S.; Heinrich, S.; Wachter, B.; Sommer, S. Gut microbiomes of free-ranging and captive Namibian cheetahs: Diversity, putative functions and occurrence of potential pathogens. Mol. Ecol. 2017, 26, 5515–5527. [Google Scholar] [CrossRef] [PubMed]
- Bizzozero, G. Ueber die schlauchfoermigen drusen des magendarmkanals und die beziehungen ihres epithels zu dem oberfachenepithel der schleimhaut. Arch. Mikrobial. Anat. 1892, 42, 82–152. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Hazell, S.L.; O’Rourke, J.; Kouprach, S. Isolation of a spiral-shaped bacterium from the cat stomach. Infect. Immun. 1988, 56, 2843–2850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salomon, H. Über das Spirillum des Säugetiermagens und sein Verhalten zu den Belegzellen; G. Fischer: Schaffhausen, Switzerland, 1896. [Google Scholar]
- Weber, A.F.; Hase, O.; Sautter, J.H. Some observations concerning the presence of spirilla in the fundic glands of dogs and cats. Am. J. Vet. Res. 1958, 18, 677–680. [Google Scholar]
- Smith, S.M. Role of Toll-like receptors in Helicobacter pylori infection and immunity. World. J. Gastrointest. Pathophysiol. 2014, 5, 133–146. [Google Scholar] [CrossRef]
- Owen, R.J. Helicobacter-species classification and identification. Br. Med. Bull. 1998, 54, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Conte, M.P.; Mastromarino, P. Microbiologia Medica: Batteriologia e Virologia; Società Editrice Esculapio: Bologna, Italy, 2015; p. 288. [Google Scholar]
- Happonen, I.; Saari, S.; Castren, L.; Tyni, O.; Hänninen, M.L.; Westermarck, E. Comparison of diagnostic methods for detecting gastric Helicobacter-like organisms in dogs and cats. J. Comp. Pathol. 1996, 115, 117–127. [Google Scholar] [CrossRef]
- Jakob, W.; Stolte, M.; Valentin, A.; Schröder, H.D. Demonstration of Helicobacter pylori-like organisms in the gastric mucosa of captive exotic carnivores. J. Comp. Pathol. 1997, 116, 21–33. [Google Scholar] [CrossRef]
- Eaton, K.A.; Dewhirst, F.E.; Radin, M.J.; Fox, J.G.; Paster, B.J.; Krakowka, S.; Morgan, D.R. Helicobacter acinonyx sp. Nov., Isolated from Cheetahs with Gastritis. Int. J. Syst. Bacteriol. 1993, 43, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangiaterra, S.; Galosi, L.; Scarpona, S.; Berardi, S.; Marini, M.C.; Cerquetella, M.; Preziuso, S.; Rossi, G. First detection of Helicobacter canis and related gastric pathology in cheetahs (Acinonyx jubatus). In Proceedings of the Atti 72° Convegno Società Italia delle Scienze Veterinarie, Torino, Italy, 20–22 June 2018; p. 225, ISBN 978-8890909214. [Google Scholar]
- Segre, J.A. What does it take to satisfy Koch’s postulates two centuries later? Microbial genomics and Propionibacteria acnes. J. Invest. Dermatol. 2013, 133, 2141–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, S.J.; Roelke, M.E.; Marker, L.; Newman, A.; Winkler, C.A.; Meltzer, D.; Colly, L.; Evermann, J.F.; Bush, M. Genetic basis for species vulnerability in the cheetah. Science 1985, 227, 1428–1434. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.V.S. The immunogenetics of human infectious diseases. Annu. Rev. Immunol. 1998, 16, 593–617. [Google Scholar] [CrossRef]
- Hale, K.A.; Briskie, J.V. Decreased immunocompetence in a severely bottlenecked population of an endemic New Zealand bird. Anim. Conserv. 2007, 10, 2–10. [Google Scholar] [CrossRef]
- Piertney, S.B.; Oliver, M.K. The evolutionary ecology of the major histocompatibility complex. Heredity 2006, 96, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Klein, J. Natural History of the Major Histocompatibility Complex, 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 1986. [Google Scholar]
- Munson, L.; Marker, L.; Dubovi, E.; Spencer, J.A.; Evermann, J.F.; O’Brien, S.J. Serosurvey of viral infections in free-ranging Namibian cheetahs (Acinonyx jubatus). J. Wildl. Dis. 2004, 40, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Thalwitzer, S.; Wachter, B.; Robert, N.; Wibbelt, G.; Müller, T.; Lonzer, J.; Meli, M.L.; Bay, G.; Hofer, H.; Lutz, H. Seroprevalences to viral pathogens in free-ranging and captive cheetahs (Acinonyx jubatus) on Namibian Farmland. Clin. Vaccine Immunol. 2010, 17, 232–238. [Google Scholar] [CrossRef] [Green Version]
- Lobetti, R. Gastritis in the captive cheetah. In Proceedings of the 28th Congress of the World Small Animal Veterinary Association, Bangkok, Thailand, 24–27 October 2003. [Google Scholar]
- Morris, J.G.; Rogers, Q.R. Arginine: An Essential Amino Acid for the Cat. J. Nutr. 1978, 108, 1944–1953. [Google Scholar] [CrossRef]
- Fletcher, M.; Ramirez, M.E.; Sierra, R.A.; Raber, P.; Thevenot, P.; Al-Khami, A.A.; Sanchez-Pino, D.; Hernandez, C.; Wyczechowska, D.D.; Ochoa, A.C.; et al. L-Arginine depletion blunts antitumor T-cell responses by inducing myeloid-derived suppressor cells. Cancer Res. 2015, 75, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarasenko, T.N.; Gomez-Rodriguez, J.; McGuire, P.J. Impaired T cell function in argininosuccinate synthetase deficiency. J. Leukoc. Biol. 2015, 97, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galván-Peña, S.; O’Neill, L.A. Metabolic reprograming in macrophage polarization. Front. Immunol. 2014, 5, 420. [Google Scholar]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoghegan, V.; Guo, A.; Trudgian, D.; Thomas, B.; Acuto, O. Comprehensive identification of arginine methylation in primary T cells reveals regulatory roles in cell signaling. Nat. Commun. 2015, 6, 6758. [Google Scholar] [CrossRef] [PubMed]
- Green, S.J.; Scheller, L.F.; Marletta, M.A.; Seguin, M.C.; Klotz, F.W.; Slayter, M.; Nelson, B.J.; Nacy, C.A. Nitric oxide: Cytokine-regulation of nitric oxide in host resistance to intracellular pathogens. Immunol. Lett. 1994, 43, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Mungrue, I.N.; Husain, M.; Stewart, D.J. The role of NOS in heart failure: Lessons from murine genetic models. Heart Fail. Rev. 2002, 7, 407–422. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Aims | No. Cheetah |
|---|---|---|
| Eaton et al., 1991 [14] | Gastritis Associated with Gastric Spiral Bacilli in Cheetahs | 25 |
| Eaton et al., 1993 [4] | Isolation of Helicobacter acinonyx from cheetah with gastritis | 4 |
| Munson, 1993 [2] | Diseases of Captive Cheetahs (Acinonyx jubatus jubatus) in South Africa | 69 |
| Lobetti et al., 1999 [8] | Prevalence of Helicobacteriosis and gastritis in Cheetahs | 28 |
| Dailidiene et al., 2004 [15] | Helicobacter pylori-like gastric pathogen of cheetahs and other big cats | 6 |
| Terio et al., 2005 [5] | Helicobacter species in captive cheetahs with gastritis | 33 |
| Lane et al., 2012 [16] | Effect of diet on the incidence of and mortality owing to gastritis and renal disease in captive cheetahs | 72 |
| Terio et al., 2012 [6] | Characterization of the gastric immune response in cheetahs with Helicobacter | 21 |
| Rossi et al., 2014 [7] | Severe gastritis with double Helicobacter spp. infection associated with Barrett’s esophagus in a cheetah | 1 |
| Whitehouse-Tedd et al., 2015 [9] | Dietary factors associated with fecal consistency and other indicators of gastrointestinal health in the cheetah | 184 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangiaterra, S.; Marker, L.; Cerquetella, M.; Galosi, L.; Marchegiani, A.; Gavazza, A.; Rossi, G. Chronic Stress-Related Gastroenteric Pathology in Cheetah: Relation between Intrinsic and Extrinsic Factors. Biology 2022, 11, 606. https://doi.org/10.3390/biology11040606
Mangiaterra S, Marker L, Cerquetella M, Galosi L, Marchegiani A, Gavazza A, Rossi G. Chronic Stress-Related Gastroenteric Pathology in Cheetah: Relation between Intrinsic and Extrinsic Factors. Biology. 2022; 11(4):606. https://doi.org/10.3390/biology11040606
Chicago/Turabian StyleMangiaterra, Sara, Laurie Marker, Matteo Cerquetella, Livio Galosi, Andrea Marchegiani, Alessandra Gavazza, and Giacomo Rossi. 2022. "Chronic Stress-Related Gastroenteric Pathology in Cheetah: Relation between Intrinsic and Extrinsic Factors" Biology 11, no. 4: 606. https://doi.org/10.3390/biology11040606