
The Effect of Date Palm Genotypes on Rhizobacterial Community Structures under Saline Environments


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Collections, Seed Sterilization, and Plant Growth Conditions
2.2. Soil Physiochemical Analysis
2.3. Gas Exchange Measurements
2.4. Root Analysis
2.5. DNA Extraction and Illumina MiSeq Sequencing of 16S rDNA
2.6. Data Analysis
3. Results
3.1. Soil Physiochemical Properties Affected by Salinity
3.2. Salinity Causes a Significant Reduction in Plant Growth
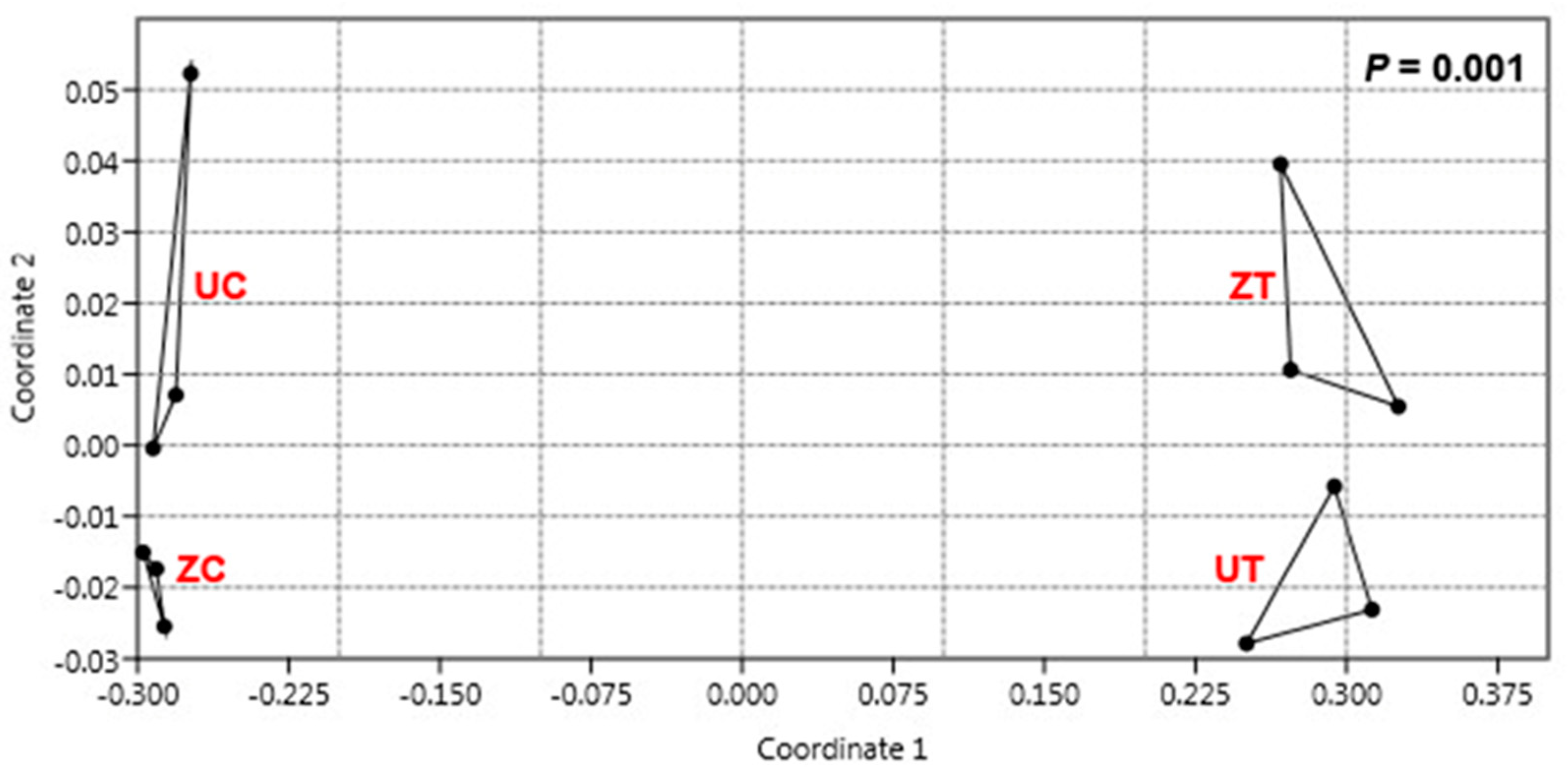
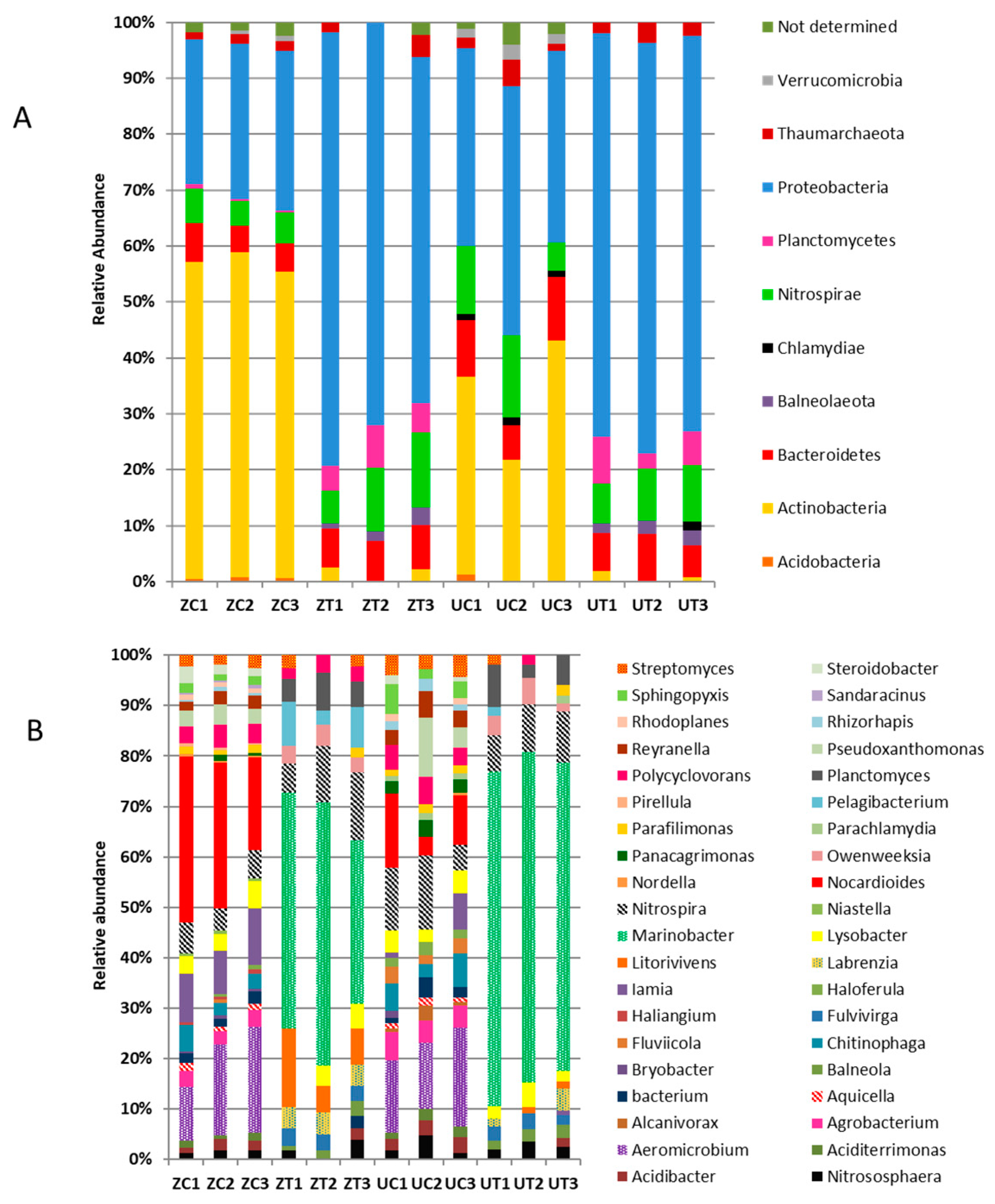
3.3. Metagenomic Analysis Revealed Differential Bacterial Community Structures in Different Cultivars
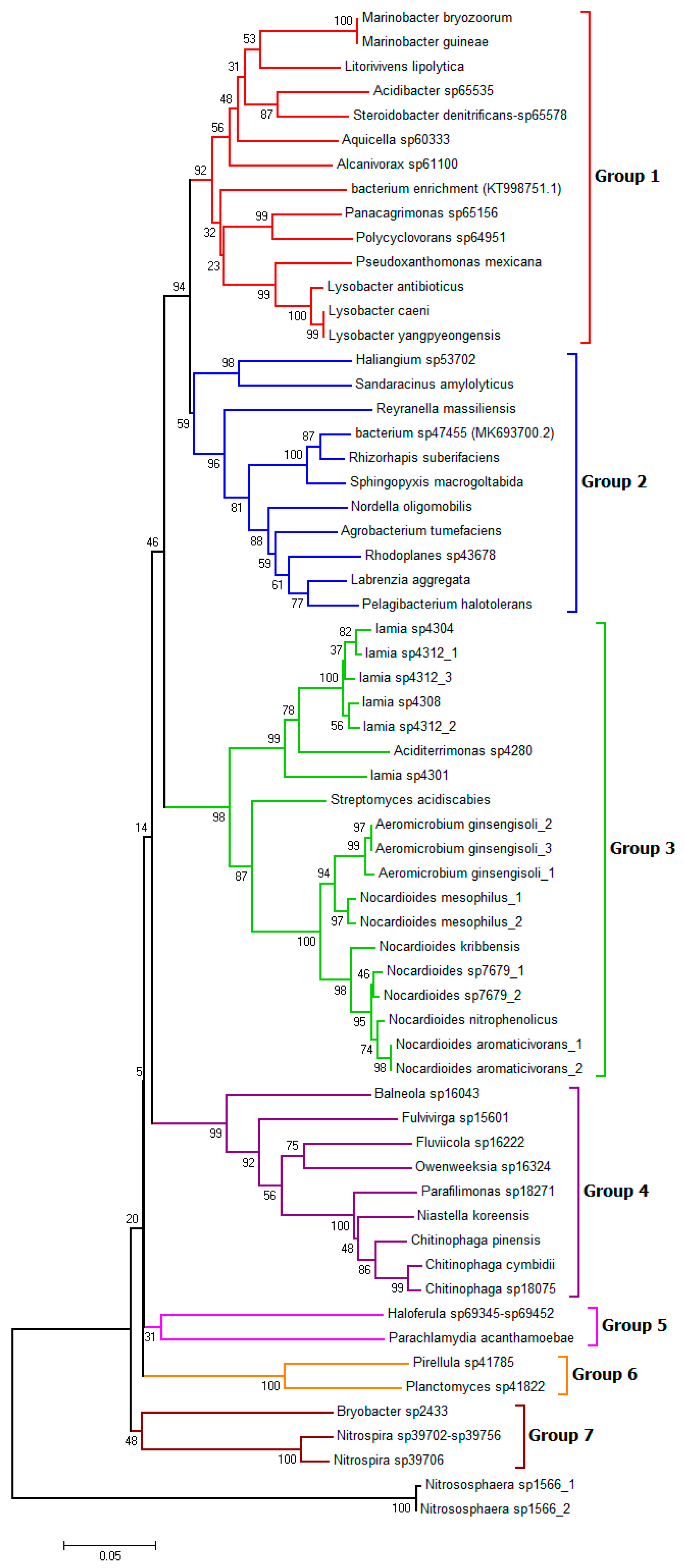
3.4. Metagenomic Analysis Revealed Differential Bacterial Abundance in the Cultivars
3.5. Salinity Affects the Structure of the Bacterial Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chao, C.T.; Krueger, R.R. The date palm (Phoenix dactylifera L.): Overview of biology, uses, and cultivation. HortScience 2007, 42, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Haplogypsids, T.C.; Haplocalcids, T.; Torrifluvents, T. Causes, origin, genesis and extent of soil salinity in the Sultanate of Oman. Pak. J. Agri. Sci 2006, 43, 1–2. [Google Scholar]
- Ben Salah, M.; Ibrahim, A. Atlas of the Main Date Palm Varieties in the Arab States of the Gulf: Beirut, Lebanon: International Center for Agricultural Research in the Dry Areas (ICARDA). 2018. Available online: https://hdl.handle.net/20.500.11766/10133 (accessed on 10 November 2022).
- Jana, G.A.; Glick, B.R.; Yaish, M.W. Chapter 14—Salt tolerance in plants: Using OMICS to assess the impact of plant growth-promoting bacteria (PGPB). In Mitigation of Plant Abiotic Stress by Microorganisms; Santoyo, G., Kumar, A., Aamir, M., Uthandi, S., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 299–320. [Google Scholar]
- Gamalero, E.; Glick, B.R. Recent Advances in Bacterial Amelioration of Plant Drought and Salt Stress. Biology 2022, 11, 437. [Google Scholar] [CrossRef]
- Sagar, A.; Rai, S.; Ilyas, N.; Sayyed, R.; Al-Turki, A.I.; El Enshasy, H.A.; Simarmata, T. Halotolerant rhizobacteria for salinity-stress mitigation: Diversity, mechanisms and molecular approaches. Sustainability 2022, 14, 490. [Google Scholar] [CrossRef]
- Dodd, I.C.; Pérez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [Green Version]
- Yaghoubi Khanghahi, M.; AbdElgawad, H.; Verbruggen, E.; Korany, S.M.; Alsherif, E.A.; Beemster, G.T.; Crecchio, C. Biofertilisation with a consortium of growth-promoting bacterial strains improves the nutritional status of wheat grain under control, drought and salinity stress conditions. Physiol. Plant. 2022, 174, e13800. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.; Malfanova, N.; Kamilova, F.; Berg, G. Plant growth promotion by microbes. Mol. Microb. Ecol. Rhizosphere 2013, 2, 561–573. [Google Scholar]
- Smith, D.L.; Gravel, V.; Yergeau, E. Signaling in the Phytomicrobiome. Front. Plant Sci. 2017, 8, 611. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Garcia, B.J.; Furches, A.; Tuskan, G.A.; Jacobson, D. Plant Host-Associated Mechanisms for Microbial Selection. Front. Plant Sci. 2019, 10, 862. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R.; Gamalero, E. Recent Developments in the Study of Plant Microbiomes. Microorganisms 2021, 9, 1533. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yao, Z.; Sun, Y.; Wang, E.; Tian, C.; Sun, Y.; Liu, J.; Sun, C.; Tian, L. Current Studies of the Effects of Drought Stress on Root Exudates and Rhizosphere Microbiomes of Crop Plant Species. Int. J. Mol. Sci. 2022, 23, 2374. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M.W.; Antony, I.; Glick, B.R. Isolation and characterization of endophytic plant growth-promoting bacteria from date palm tree (Phoenix dactylifera L.) and their potential role in salinity tolerance. Antonie Van Leeuwenhoek 2015, 107, 1519–1532. [Google Scholar] [CrossRef] [PubMed]
- Gavlak, R.; Horneck, D.; Miller, R.O.; Kotuby-Amacher, J. Soil, Plant and Water Reference Methods for the Western Region, 2nd ed.; WCC-103 Publication: Fort Collins, CO, USA, 2003. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- IBM. IBM SPSS Statistics for Windows, Version 21.0; IBM: Armonk, NY, USA, 2012.
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Al Kharusi, L.; Yahyai, R.A.; Yaish, M.W. Antioxidant response to salinity in salt-tolerant and salt-susceptible cultivars of date palm. Agriculture 2019, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Pii, Y.; Borruso, L.; Brusetti, L.; Crecchio, C.; Cesco, S.; Mimmo, T. The interaction between iron nutrition, plant species and soil type shapes the rhizosphere microbiome. Plant Physiol. Biochem. 2016, 99, 39–48. [Google Scholar] [CrossRef]
- Zhang, K.; Shi, Y.; Cui, X.; Yue, P.; Li, K.; Liu, X.; Tripathi, B.M.; Chu, H. Salinity is a key determinant for soil microbial communities in a desert ecosystem. MSystems 2019, 4, e00225-18. [Google Scholar] [CrossRef] [Green Version]
- Borruso, L.; Bacci, G.; Mengoni, A.; De Philippis, R.; Brusetti, L. Rhizosphere effect and salinity competing to shape microbial communities in Phragmites australis (Cav.) Trin. ex-Steud. FEMS Microbiol. Lett. 2014, 359, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Liu, R.; Zhang, H.; Yun, J. The diversity and abundance of bacteria and oxygenic phototrophs in saline biological desert crusts in Xinjiang, Northwest China. Microb. Ecol. 2013, 66, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Ponsford, J.C.B.; Hubbard, C.J.; Harrison, J.G.; Maignien, L.; Buerkle, C.A.; Weinig, C. Whole-genome duplication and host genotype affect rhizosphere microbial communities. bioRxiv 2020, 822726. [Google Scholar] [CrossRef] [PubMed]
- Berlanas, C.; Berbegal, M.; Elena, G.; Laidani, M.; Cibriain, J.F.; Sagües, A.; Gramaje, D. The fungal and bacterial rhizosphere microbiome associated with grapevine rootstock genotypes in mature and young vineyards. Front. Microbiol. 2019, 10, 1142. [Google Scholar] [CrossRef] [Green Version]
- Aira, M.; Gómez-Brandón, M.; Lazcano, C.; Bååth, E.; Domínguez, J. Plant genotype strongly modifies the structure and growth of maize rhizosphere microbial communities. Soil Biol. Biochem. 2010, 42, 2276–2281. [Google Scholar] [CrossRef]
- Liu, F.; Hewezi, T.; Lebeis, S.L.; Pantalone, V.; Grewal, P.S.; Staton, M.E. Soil indigenous microbiome and plant genotypes cooperatively modify soybean rhizosphere microbiome assembly. BMC Microbiol. 2019, 19, 201. [Google Scholar] [CrossRef] [Green Version]
- Marques, J.M.; da Silva, T.F.; Vollu, R.E.; Blank, A.F.; Ding, G.-C.; Seldin, L.; Smalla, K. Plant age and genotype affect the bacterial community composition in the tuber rhizosphere of field-grown sweet potato plants. FEMS Microbiol. Ecol. 2014, 88, 424–435. [Google Scholar] [CrossRef]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczyk, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Koch, A.L. Oligotrophs versus copiotrophs. Bioessays 2001, 23, 657–661. [Google Scholar] [CrossRef]
- Wong, V.N.; Greene, R.; Dalal, R.C.; Murphy, B.W. Soil carbon dynamics in saline and sodic soils: A review. Soil Use Manag. 2010, 26, 2–11. [Google Scholar] [CrossRef]
- Xie, K.; Deng, Y.; Zhang, S.; Zhang, W.; Liu, J.; Xie, Y.; Zhang, X.; Huang, H. Prokaryotic community distribution along an ecological gradient of salinity in surface and subsurface saline soils. Sci. Rep. 2017, 7, 13332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, D.; Loria, R. Streptomyces acidiscabies sp. nov. Int. J. Syst. Evol. Microbiol. 1989, 39, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Canfora, L.; Bacci, G.; Pinzari, F.; Papa, G.L.; Dazzi, C.; Benedetti, A. Salinity and bacterial diversity: To what extent does the concentration of salt affect the bacterial community in a saline soil? PLoS ONE 2014, 9, e114658. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, J.A.; Sagulenko, E. Beyond the bacterium: Planctomycetes challenge our concepts of microbial structure and function. Nat. Rev. Microbiol. 2011, 9, 403–413. [Google Scholar] [CrossRef]
- Yaghoubi Khanghahi, M.; Crecchio, C.; Verbruggen, E. Shifts in the rhizosphere and endosphere colonizing bacterial communities under drought and salinity stress as affected by a biofertilizer consortium. Microb. Ecol. 2022, 84, 483–495. [Google Scholar] [CrossRef]
- Collingro, A.; Köstlbacher, S.; Horn, M. Chlamydiae in the Environment. Trends Microbiol. 2020. [Google Scholar] [CrossRef]
- Yaish, M.W.; Al-Harrasi, I.; Alansari, A.S.; Al-Yahyai, R.; Glick, B.R. The use of high throughput DNA sequence analysis to assess the endophytic microbiome of date palm roots grown under different levels of salt stress. Int. Microbiol 2016, 19, 143–155. [Google Scholar]
- Xu, T.; Yu, M.; Liu, J.; Lin, H.; Liang, J.; Zhang, X.-H. Role of RpoN from Labrenzia aggregata LZB033 (Rhodobacteraceae) in formation of flagella and biofilms, motility, and environmental adaptation. Appl. Environ. Microbiol. 2019, 85, e02844-18. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, J.-M.; Kang, C.-H.; Yoon, J.-H. Litorivivens lipolytica gen. nov., sp. nov., a lipolytic bacterium isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 2015, 65, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.-W.; Huo, Y.-Y.; Wang, C.-S.; Oren, A.; Cui, H.-L.; Vedler, E.; Wu, M. Pelagibacterium halotolerans gen. nov., sp. nov. and Pelagibacterium luteolum sp. nov., novel members of the family Hyphomicrobiaceae. Int. J. Syst. Evol. Microbiol. 2011, 61, 1817–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montes, M.J.; Bozal, N.; Mercadé, E. Marinobacter guineae sp. nov., a novel moderately halophilic bacterium from an Antarctic environment. Int. J. Syst. Evol. Microbiol. 2008, 58, 1346–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, R.; Raddadi, N.; Fava, F. Mediterranean Sea bacteria as a potential source of long-chain polyunsaturated fatty acids. FEMS Microbiol. Lett. 2020, 367, fnaa132. [Google Scholar] [CrossRef] [PubMed]
- van Bruggen, A.H.; Francis, I.M.; Krag, R. The vicious cycle of lettuce corky root disease: Effects of farming system, nitrogen fertilizer and herbicide. Plant Soil 2015, 388, 119–132. [Google Scholar] [CrossRef]
- Kubota, M.; Kawahara, K.; Sekiya, K.; Uchida, T.; Hattori, Y.; Futamata, H.; Hiraishi, A. Nocardioides aromaticivorans sp. nov., a dibenzofuran-degrading bacterium isolated from dioxin-polluted environments. Syst. Appl. Microbiol. 2005, 28, 165–174. [Google Scholar] [CrossRef]
- Hara, A.; Syutsubo, K.; Harayama, S. Alcanivorax which prevails in oil-contaminated seawater exhibits broad substrate specificity for alkane degradation. Environ. Microbiol. 2003, 5, 746–753. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Physiochemical Properties | Average Content | p-Value | Average Content | p-Value | ||
|---|---|---|---|---|---|---|
| UC | UT | ZC | ZT | |||
| EC (dS/m) | 4.37 | 62.77 | 0 | 4.35 | 59.9 | 0 |
| pH | 8.01 | 7.54 | 0 | 7.93 | 7.41 | 0 |
| P (ppm) | 156.47 | 100.67 | 0.07 | 127.37 | 118.1 | 0.25 |
| K (ppm) | 136.67 | 133.33 | 0.26 | 116.67 | 123.33 | 0.27 |
| Na (ppm) | 886.67 | 3620 | 0 | 620 | 3670 | 0 |
| Coarse sand (%) | 12.67 | 13.25 | 0.19 | 9.08 | 9.98 | 0.31 |
| Fine sand (%) | 54.37 | 51.78 | 0.13 | 57.95 | 54.39 | 0.16 |
| Silt (%) | 29.33 | 26 | 0.13 | 27.33 | 24.67 | 0.22 |
| Clay (%) | 3.63 | 8.97 | 0.02 | 5.63 | 10.97 | 0.03 |
| CaCO3 (%) | 20.93 | 19.9 | 0.04 | 18.73 | 16.97 | 0.04 |
| Cl (ppm) | 498.96 | 19.721.26 | 0 | 496.86 | 20.887.83 | 0 |
| SO4 (ppm) | 438.71 | 337.48 | 0.031 | 457.66 | 391.30 | 0.14 |
| Mg (ppm) | 338.73 | 1791.12 | 0 | 331.77 | 1510.70 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Busaidi, A.; Glick, B.R.; Yaish, M.W. The Effect of Date Palm Genotypes on Rhizobacterial Community Structures under Saline Environments. Biology 2022, 11, 1666. https://doi.org/10.3390/biology11111666
Al-Busaidi A, Glick BR, Yaish MW. The Effect of Date Palm Genotypes on Rhizobacterial Community Structures under Saline Environments. Biology. 2022; 11(11):1666. https://doi.org/10.3390/biology11111666
Chicago/Turabian StyleAl-Busaidi, Aya, Bernard R. Glick, and Mahmoud W. Yaish. 2022. "The Effect of Date Palm Genotypes on Rhizobacterial Community Structures under Saline Environments" Biology 11, no. 11: 1666. https://doi.org/10.3390/biology11111666