
Ameliorating Drought Effects in Wheat Using an Exclusive or Co-Applied Rhizobacteria and ZnO Nanoparticles
 , , ,
, , ,  ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Layout and Crop Husbandry
2.2. Drought Imposition
2.3. PGPR and Nanoparticles Application
2.4. Data Recording and Related Procedures
2.4.1. Growth and Yield Parameters
2.4.2. Wheat Physiological Characteristics
2.4.3. Quality Parameters
2.4.4. Enzymatic and Non-Enzymatic Antioxidants Activity Assays
2.5. Data Processing Software Usage for Analysis
3. Results
3.1. Growth and Yield Characters
3.2. Plant Morphological and Grain Quality
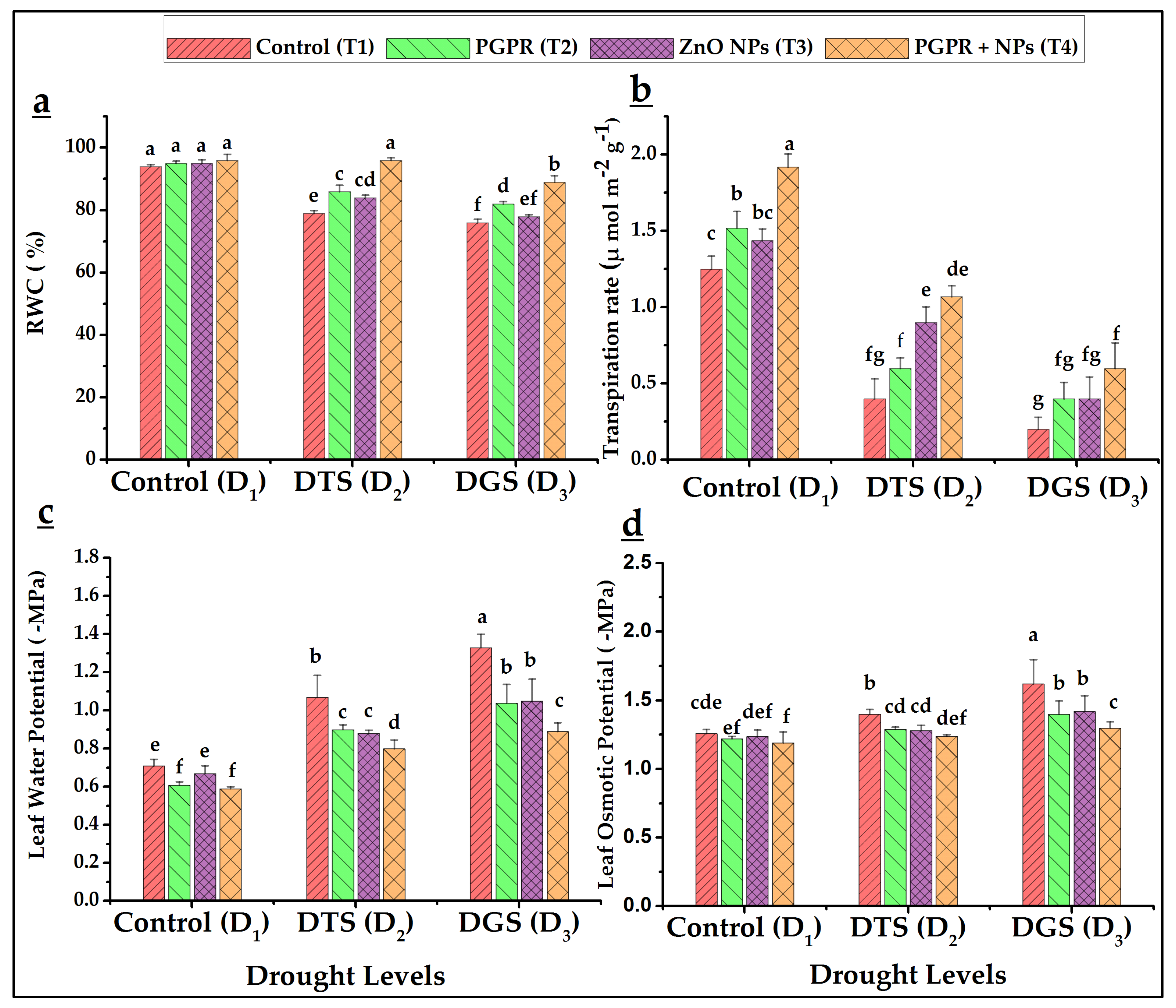
3.3. Physiological Characteristics
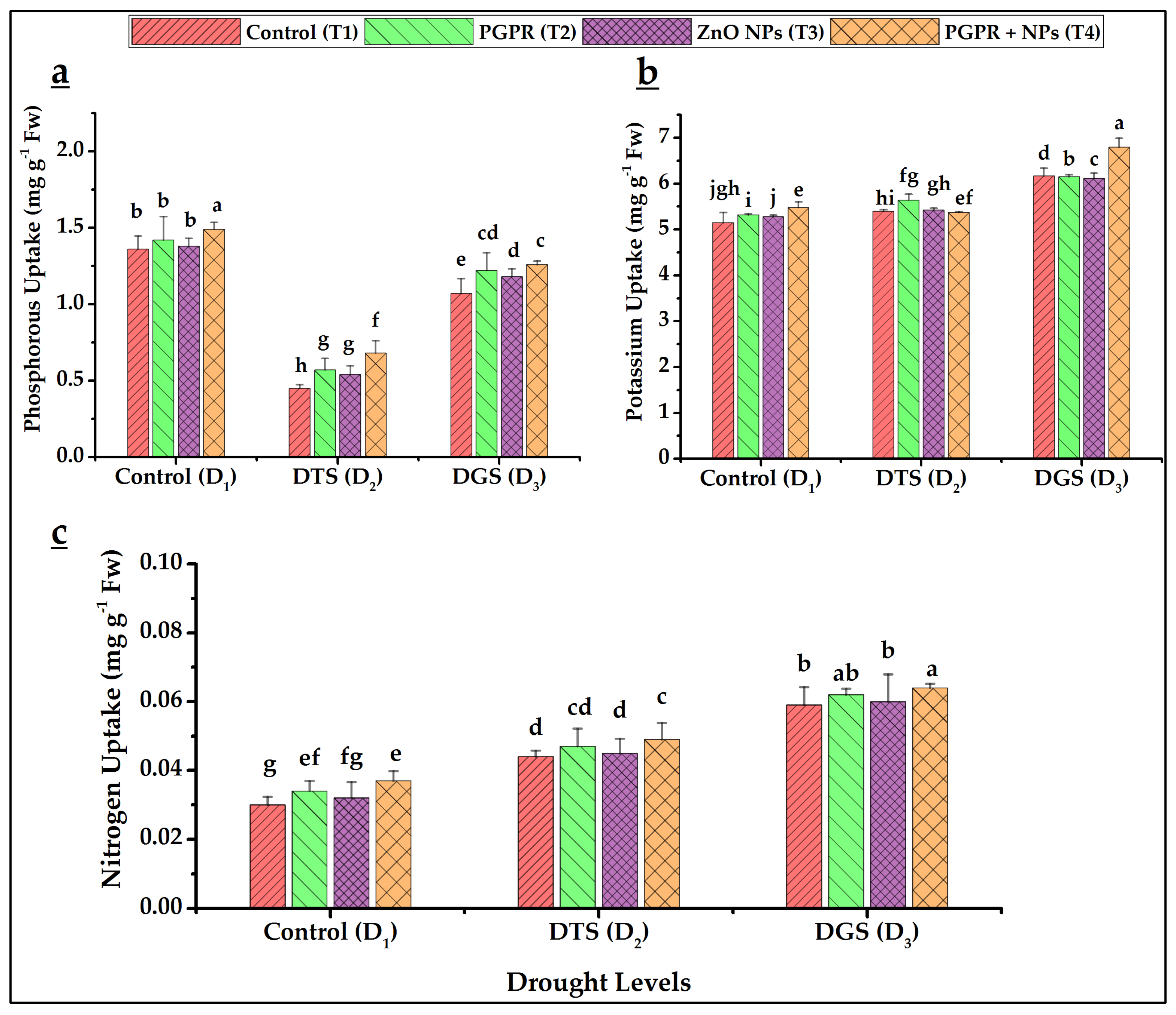
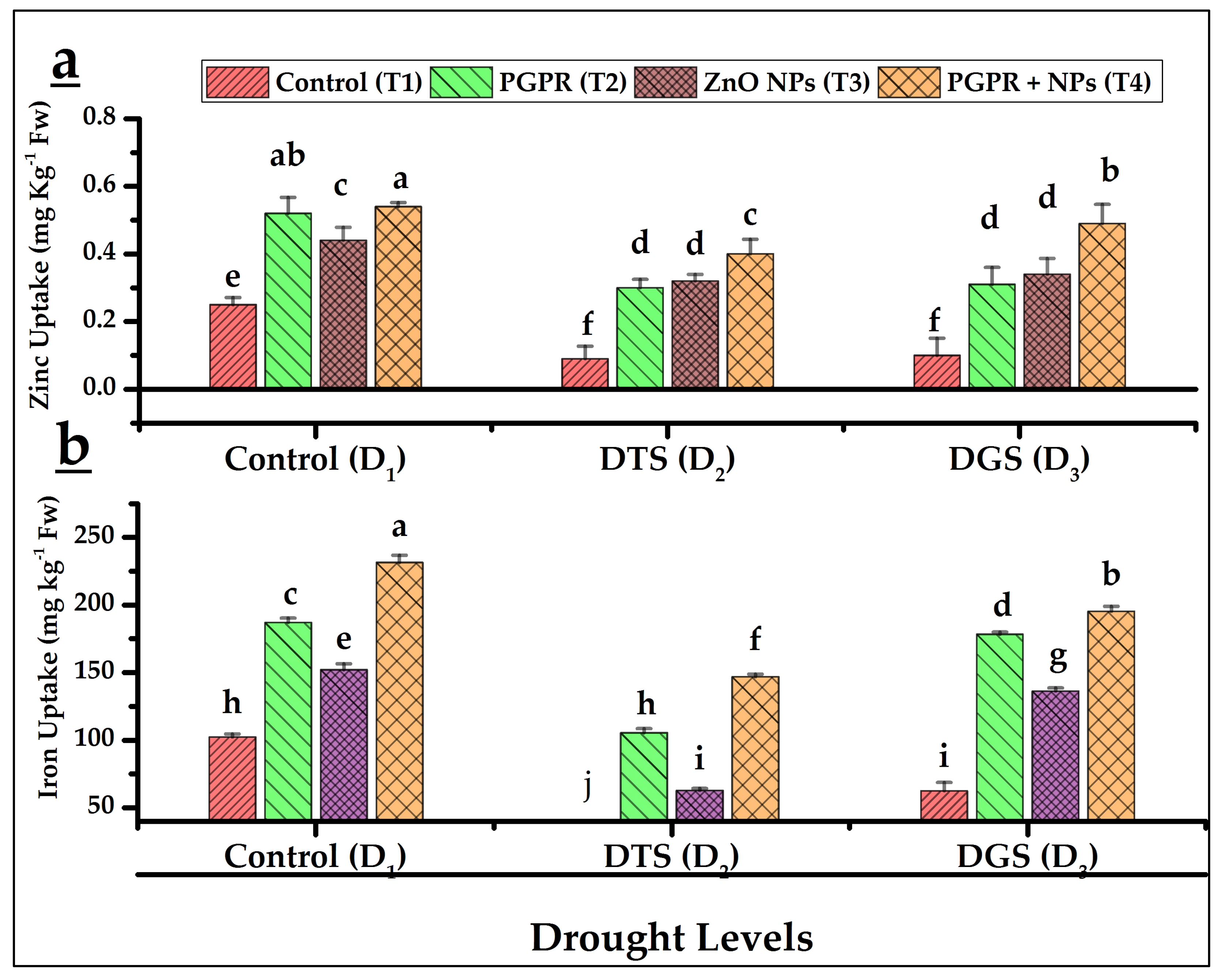
3.4. Nutrients Uptake Attributions
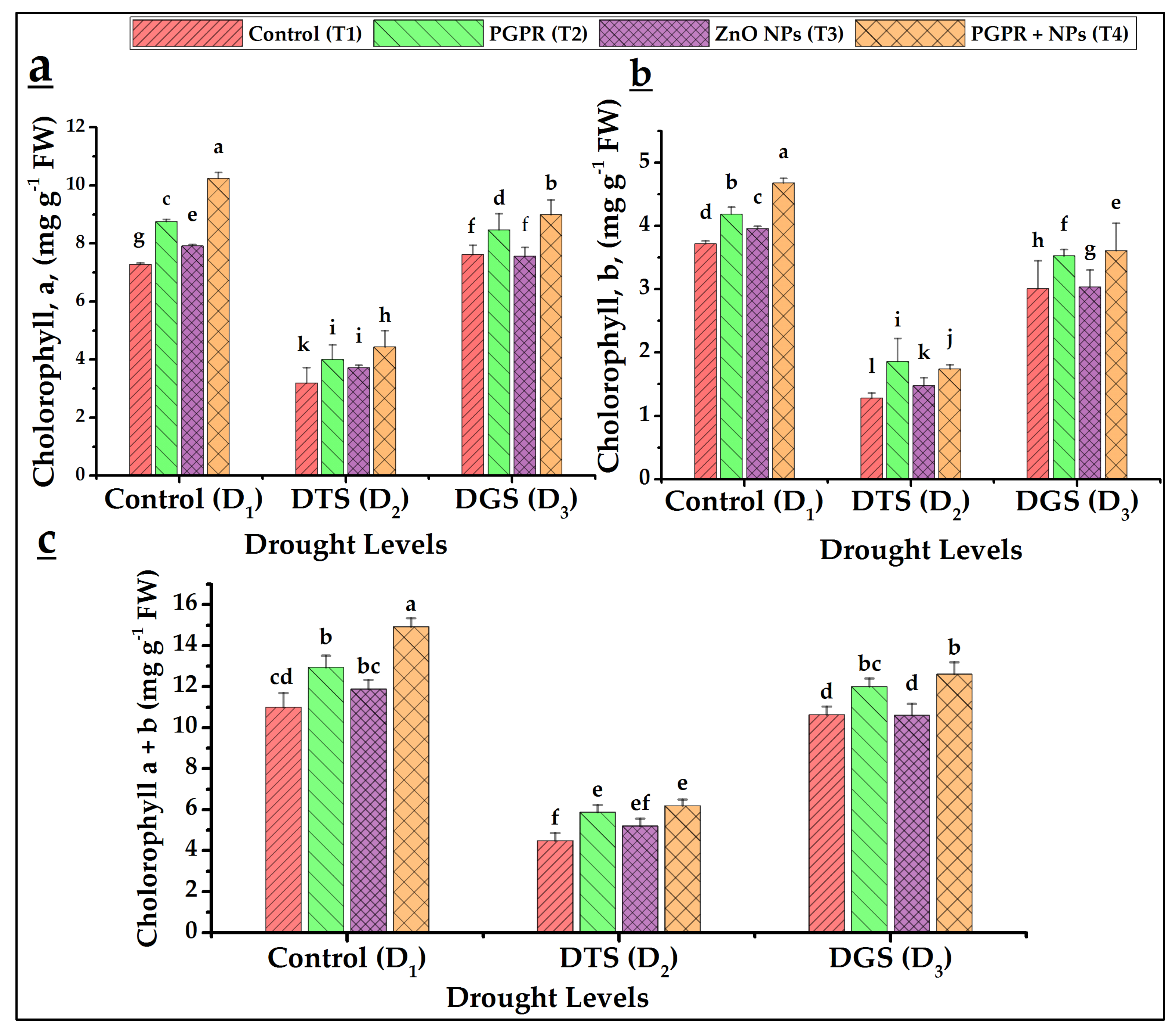
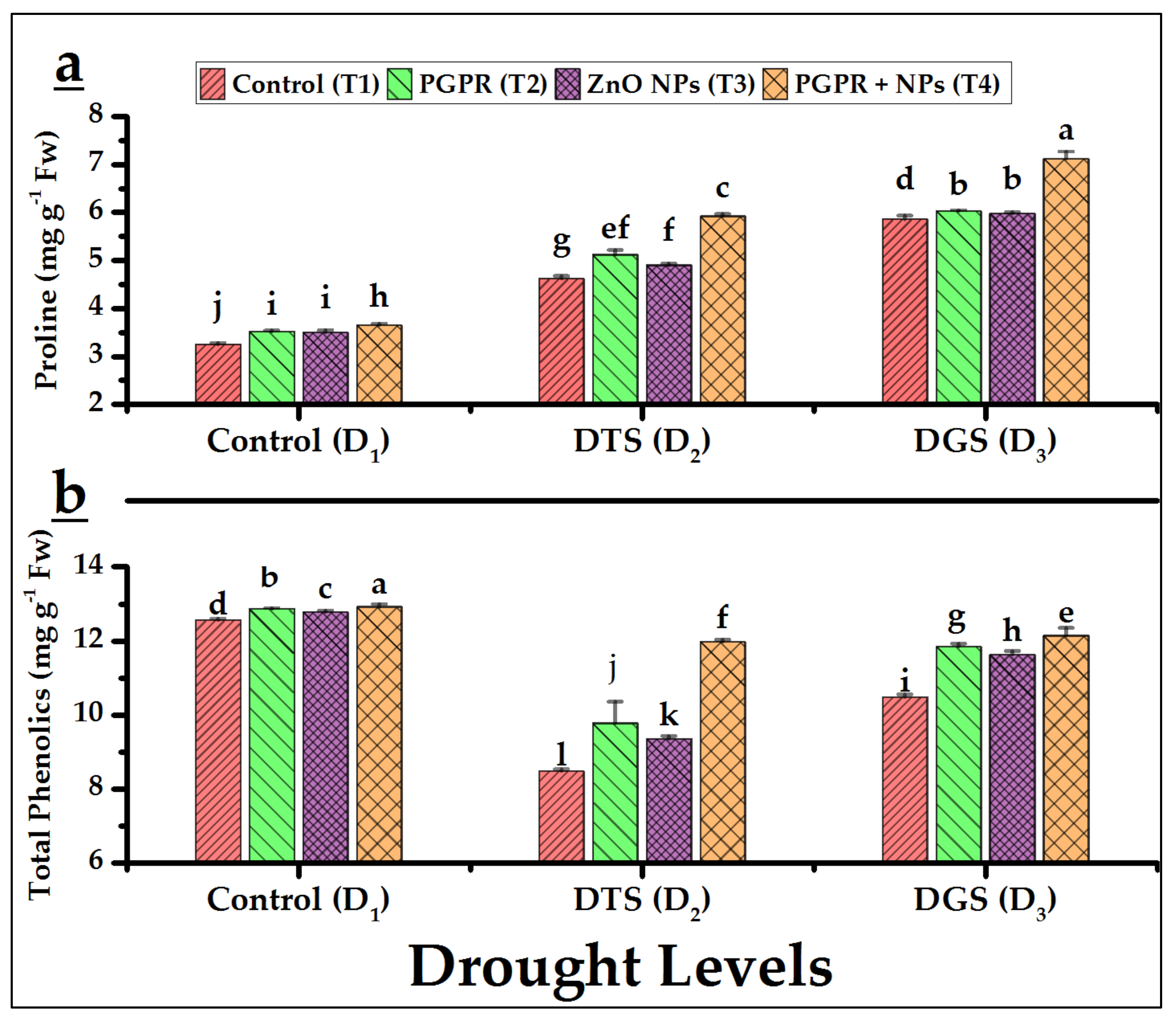
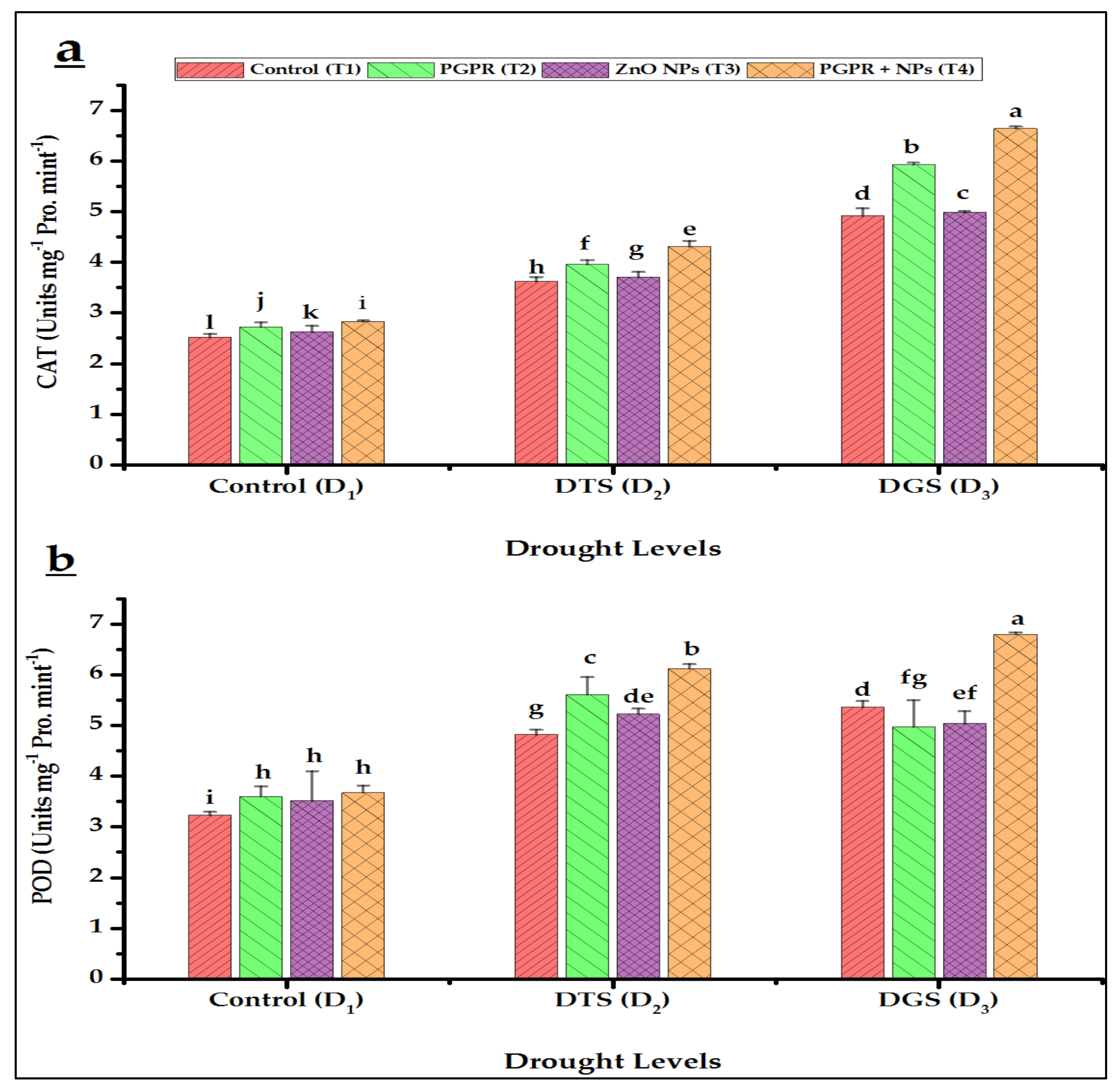
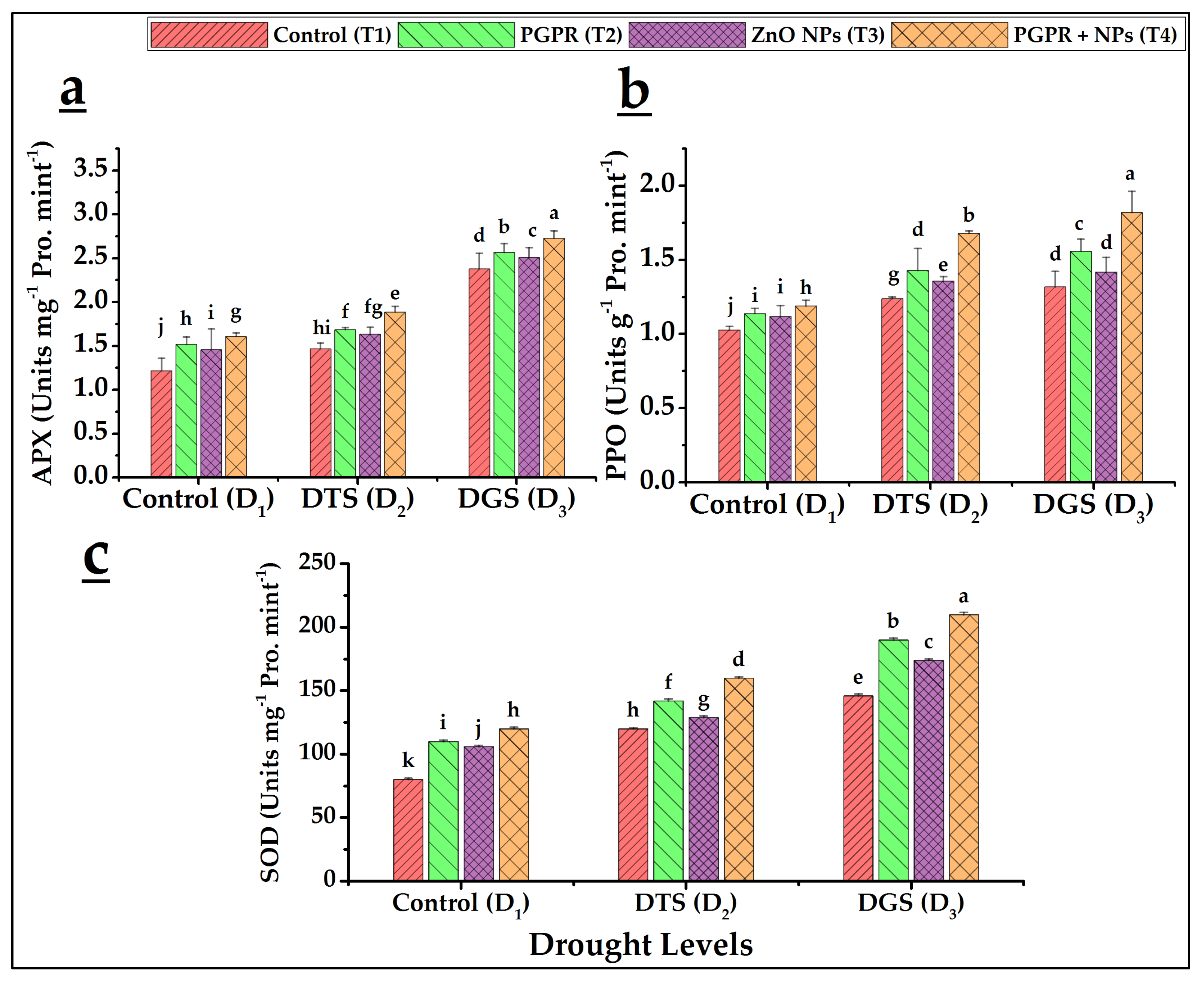
3.5. Biochemical Characteristics
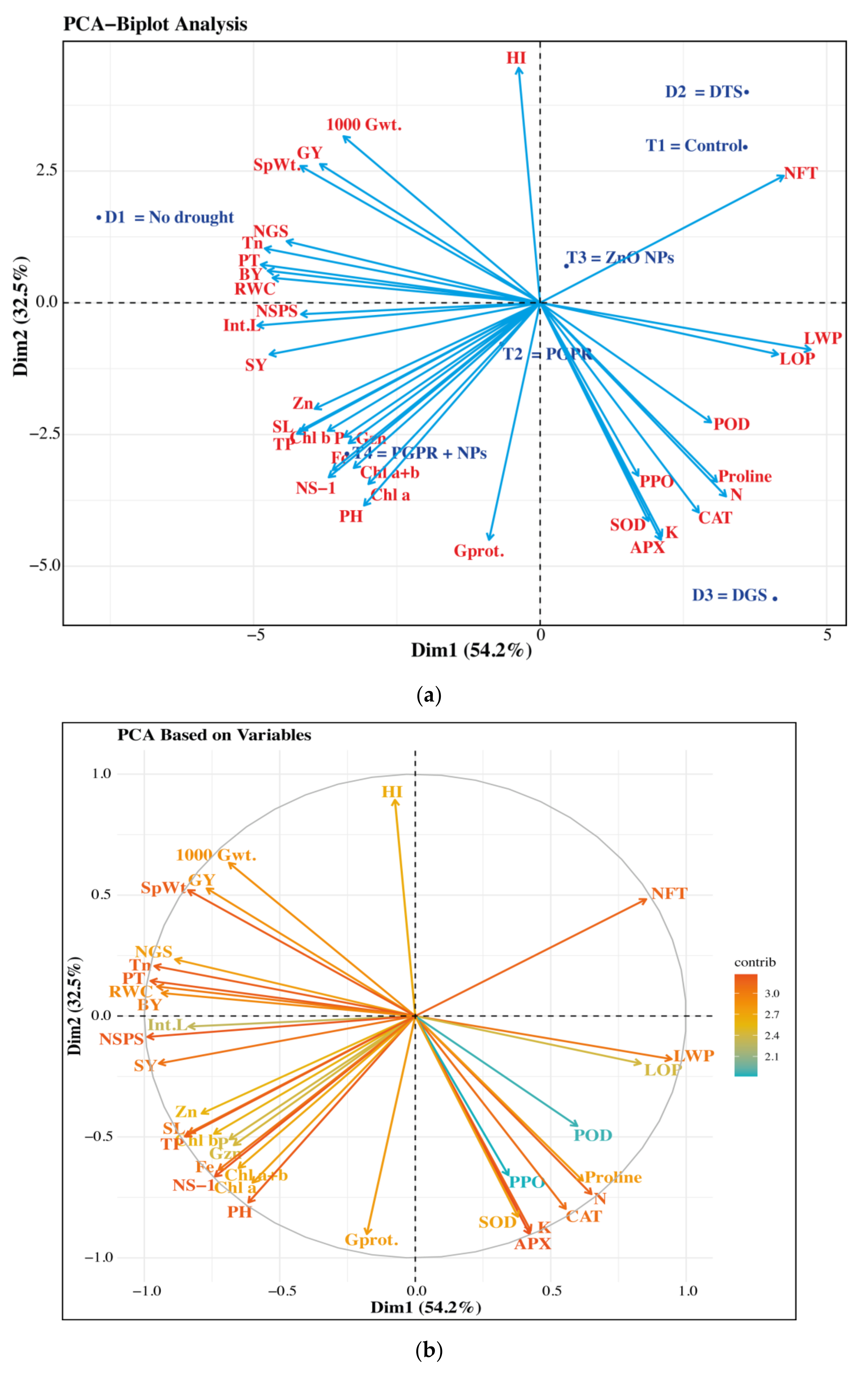
3.6. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, A.; Ullah, S.; Dar, A.A.; Sardar, M.F.; Mehmood, T.; Tufail, M.A.; Shakoor, A.; Haris, M. Nexus on climate change: Agriculture and possible solution to cope future climate change stresses. Environ. Sci. Pollut. Res. 2021, 28, 14211–14232. [Google Scholar] [CrossRef] [PubMed]
- Rashid, U.; Yasmin, H.; Hassan, M.N.; Naz, R.; Nosheen, A.; Sajjad, M.; Ilyas, N.; Keyani, R.; Jabeen, Z.; Mumtaz, S. Drought-tolerant Bacillus megaterium isolated from semi-arid conditions induces systemic tolerance of wheat under drought conditions. Plant Cell Rep. 2022, 41, 549–569. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.; Baenziger, P.S.; Börner, A. Drought stress tolerance in wheat and barley: Advances in physiology, breeding and genetics research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and abiotic stresses in plants. In Abiotic and Biotic Stress in Plants; InTechOpen: London, UK, 2019; pp. 1–19. [Google Scholar] [CrossRef] [Green Version]
- Majid, S.A.; Asghar, R.; Murtaza, G. Yield stability analysis conferring adaptation of wheat to pre-and post-anthesis drought conditions. Pak J. Bot. 2007, 39, 1623–1637. [Google Scholar]
- Tiwari, R.; Sheoran, S.; Rane, J. Wheat improvement for drought and heat tolerance. In Recent trends on Production Strategies of Wheat in India’; Shukla, R.S., Mishra, P.C., Chatrath, R., Gupta, R.K., Tomar, S.S., Sharma, I., Eds.; Directorate of Wheat Research: Karnal, India, 2014. [Google Scholar]
- Kulkarni, M.; Soolanayakanahally, R.; Ogawa, S.; Uga, Y.; Selvaraj, M.G.; Kagale, S. Drought response in wheat: Key genes and regulatory mechanisms controlling root system architecture and transpiration efficiency. Front. Chem. 2017, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Tambussi, E.A.; Bartoli, C.G.; Beltrano, J.; Guiamet, J.J.; Araus, J.L. Oxidative damage to thylakoid proteins in water-stressed leaves of wheat (Triticum aestivum). Physiol. Plant. 2000, 108, 398–404. [Google Scholar] [CrossRef]
- Mattos, L.; Moretti, C. Oxidative stress in plants under drought conditions and the role of different enzymes. Enzym. Eng. 2015, 5, 1–6. [Google Scholar] [CrossRef]
- Huseynova, I.M.; Rustamova, S.M.; Suleymanov, S.Y.; Aliyeva, D.R.; Mammadov, A.C.; Aliyev, J.A. Drought-induced changes in photosynthetic apparatus and antioxidant components of wheat (Triticum durum Desf.) varieties. Photosynth. Res. 2016, 130, 215–223. [Google Scholar] [CrossRef]
- Bakht, S.; Safdar, K.; Khair, K.; Fatima, A.; Fayyaz, A.; Ali, S.; Munir, H.; Farid, M. The response of major food crops to drought stress: Physiological and biochemical responses. In Agronomic Crops; Springer: Singapore, 2020; pp. 93–115. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Farooqi, Z.U.R.; Ayub, M.A.; ur Rehman, M.Z.; Sohail, M.I.; Usman, M.; Khalid, H.; Naz, K. Regulation of drought stress in plants. In Plant Life under Changing Environment; Tripathi, D.K., Singh, V.P., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 77–104. [Google Scholar] [CrossRef]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Abdellatif, Y.M. Effect of maltose and trehalose on growth, yield and some biochemical components of wheat plant under water stress. Ann. Agric. Sci. 2016, 61, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Bruno, E.; Zilberman, D. Compound natural and human disasters: Managing drought and COVID-19 to sustain global agriculture and food sectors. Sci. Total Environ. 2021, 754, 142210. [Google Scholar] [CrossRef]
- Mancosu, N.; Snyder, R.L.; Kyriakakis, G.; Spano, D. Water scarcity and future challenges for food production. Water 2015, 7, 975–992. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, L.; Wang, A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-biotechnology in agriculture: Use of nanomaterials to promote plant growth and stress tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef]
- Freitas, D.C.; de Andrade, A.M.; da Costa, L.F.; Azevedo, R.A.; Arruda, M.A. There is plenty of room at the plant science: A review of nanoparticles applied to plant cultures. Ann. Appl. Biol. 2021, 178, 149–168. [Google Scholar] [CrossRef]
- Chugh, G.; Siddique, K.H.; Solaiman, Z.M. Nanobiotechnology for agriculture: Smart technology for combating nutrient deficiencies with nanotoxicity challenges. Sustainability 2021, 13, 1781. [Google Scholar] [CrossRef]
- Zhao, L.; Peralta-Videa, J.R.; Rico, C.M.; Hernandez-Viezcas, J.A.; Sun, Y.; Niu, G.; Servin, A.; Nunez, J.E.; Duarte-Gardea, M.; Gardea-Torresdey, J.L. CeO2 and ZnO nanoparticles change the nutritional qualities of cucumber (Cucumis sativus). J. Agric. Food Chem. 2014, 62, 2752–2759. [Google Scholar] [CrossRef]
- Mukherjee, A.; Sun, Y.; Morelius, E.; Tamez, C.; Bandyopadhyay, S.; Niu, G.; White, J.C.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Differential toxicity of bare and hybrid ZnO nanoparticles in green pea (Pisum sativum L.): A life cycle study. Front. Plant Sci. 2016, 6, 1242. [Google Scholar] [CrossRef] [Green Version]
- Prasad, T.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Reddy, K.R.; Sreeprasad, T.; Sajanlal, P.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Subbaiah, L.V.; Prasad, T.N.V.K.V.; Krishna, T.G.; Sudhakar, P.; Reddy, B.R.; Pradeep, T. Novel effects of nanoparticulate delivery of zinc on growth, productivity, and zinc biofortification in maize (Zea mays L.). J. Agric. Food Chem. 2016, 64, 3778–3788. [Google Scholar] [CrossRef] [PubMed]
- De Vries, F.T.; Griffiths, R.I.; Knight, G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 39, 1620–1630. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.N.H.; Yong, J.W.H. Harnessing synergistic biostimulatory processes: A plausible approach for enhanced crop growth and resilience in organic farming. Biology 2021, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cook, J.; Nearing, J.T.; Zhang, J.; Raudonis, R.; Glick, B.R.; Langille, M.G.; Cheng, Z. Harnessing the plant microbiome to promote the growth of agricultural crops. Microbiol. Res. 2021, 245, 126690. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant growth-promoting bacteria: Biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef]
- Aslam, M.U.; Raza, M.A.S.; Saleem, M.F.; Waqas, M.; Iqbal, R.; Ahmad, S.; Haider, I. Improving strategic growth stage-based drought tolerance in quinoa by rhizobacterial inoculation. Commun. Soil Sci. Plant Anal. 2020, 51, 853–868. [Google Scholar] [CrossRef]
- Xi, L.; Zhang, M.; Zhang, L.; Lew, T.T.; Lam, Y.M. Novel materials for urban farming. Adv. Mater. 2022, 34, 2105009. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Nayana, A.; Joseph, B.J.; Jose, A.; Radhakrishnan, E. Nanotechnological advances with PGPR applications. In Sustainable Agriculture Reviews 41; Springer: Berlin/Heidelberg, Germany, 2020; pp. 163–180. [Google Scholar] [CrossRef]
- Bhatia, R.; Gulati, D.; Sethi, G. Biofilms and nanoparticles: Applications in agriculture. Folia Microbiol. 2021, 66, 159–170. [Google Scholar] [CrossRef]
- Awan, S.; Shahzadi, K.; Javad, S.; Tariq, A.; Ahmad, A.; Ilyas, S. A preliminary study of influence of zinc oxide nanoparticles on growth parameters of Brassica oleracea var italic. J. Saudi Soc. Agric. Sci. 2021, 20, 18–24. [Google Scholar] [CrossRef]
- Babaei, K.; Seyed Sharifi, R.; Pirzad, A.; Khalilzadeh, R. Effects of bio fertilizer and nano Zn-Fe oxide on physiological traits, antioxidant enzymes activity and yield of wheat (Triticum aestivum L.) under salinity stress. J. Plant Interact. 2017, 12, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.A.; Aslam, S.; Akbar, M.; Ahmad, A.; Khan, W.U.; Yasin, N.A.; Ali, B.; Rizwan, M.; Ali, S. Combined effect of Bacillus fortis IAGS 223 and zinc oxide nanoparticles to alleviate cadmium phytotoxicity in Cucumis melo. Plant Physiol. Biochem. 2021, 158, 1–12. [Google Scholar] [CrossRef]
- Laghari, G.M.; Oad, F.C.; Tunio, S.; Chachar, Q.; Gandahi, A.W.; Siddiqui, M.H.; Hassan, S.W.U.; Ali, A. Growth and yield attributes of wheat at different seed rates. Sarhad J. Agric. 2011, 27, 177–183. [Google Scholar]
- Khalilzadeh, R.; Seyed Sharifi, R.; Jalilian, J. Antioxidant status and physiological responses of wheat (Triticum aestivum L.) to cycocel application and bio fertilizers under water limitation condition. J. Plant Interact. 2016, 11, 130–137. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Müller, J. Dumas or Kjeldahl for reference analysis. In Comparison and Considerations for Nitrogen/Protein Analysis of Food and Feed; Foss: Hillerød, Denmark, 2017. [Google Scholar]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I. Activity of ascorbate-dependent H2O2-scavenging enzymes and leaf chlorosis are enhanced in magnesium-and potassium-deficient leaves, but not in phosphorus-deficient leaves. J. Exp. Bot. 1994, 45, 1259–1266. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, I1.1.1–I1.1.8. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Zaheer, M.S.; Saleem, M.F.; Khan, I.H.; Ahmad, S.; Iqbal, R. Drought ameliorating effect of exogenous applied cytokinin in wheat. Pak. J. Agric. Sci. 2020, 57, 725–733. [Google Scholar]
- Azmat, A.; Tanveer, Y.; Yasmin, H.; Hassan, M.N.; Shahzad, A.; Reddy, M.; Ahmad, A. Coactive role of zinc oxide nanoparticles and plant growth promoting rhizobacteria for mitigation of synchronized effects of heat and drought stress in wheat plants. Chemosphere 2022, 297, 133982. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.A.S.; Saleem, M.F.; Ashraf, M.Y.; Ali, A.; Asghar, H.N. Glycinebetaine applied under drought improved the physiological efficiency of wheat (Triticum aestivum L.). Plant Soil Environ. 2012, 31, 67–71. [Google Scholar]
- Trenberth, K.E.; Dai, A.; Van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Boonjung, H.; Fukai, S. Effects of soil water deficit at different growth stages on rice growth and yield under upland conditions. 1. Growth during drought. Field Crops Res. 1996, 48, 37–45. [Google Scholar] [CrossRef]
- Wisal, M.; Shah, S.M.; Shehzadi, S.; Shah, S.A.; Nawaz, H. Wheat and oat yields and water use efficiency as influenced by tillage under rainfed condition. Soil Environ. 2006, 25, 48–54. [Google Scholar]
- Farooq, M.; Hussain, M.; Siddique, K.H. Drought stress in wheat during flowering and grain-filling periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Raheem, A.; Shaposhnikov, A.; Belimov, A.A.; Dodd, I.C.; Ali, B. Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arch. Agron. Soil Sci. 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Munir, T.; Rizwan, M.; Kashif, M.; Shahzad, A.; Ali, S.; Amin, N.; Zahid, R.; Alam, M.; Imran, M. Effect of zinc oxide nanoparticles on the growth and Zn uptake in wheat (Triticum aestivum L.) by seed priming method. Dig. J. Nanomater. Biostructures 2018, 13, 315–323. [Google Scholar]
- Hussain, M.; Asgher, Z.; Tahir, M.; Ijaz, M.; Shahid, M.; Ali, H.; Sattar, A. Bacteria in combination with fertilizers improve growth, productivity and net returns of wheat (Triticum aestivum L.). Pak. J. Agric. Sci. 2016, 53. [Google Scholar] [CrossRef]
- Alharby, H.F.; Ali, S. Combined Role of Fe Nanoparticles (Fe NPs) and Staphylococcus aureus L. in the alleviation of chromium stress in rice plants. Life 2022, 12, 338. [Google Scholar] [CrossRef]
- Timmusk, S.; Seisenbaeva, G.; Behers, L. Titania (TiO2) nanoparticles enhance the performance of growth-promoting rhizobacteria. Sci. Rep. 2018, 8, 617. [Google Scholar] [CrossRef]
- Sial, M.A.; Arain, M.A.; Khanzada, S.; Naqvi, M.H.; Dahot, M.U.; Nizamani, N.A. Yield and quality parameters of wheat genotypes as affected by sowing dates and high temperature stress. Pak J. Bot. 2005, 37, 575. [Google Scholar]
- Farahbakhsh, H.; Sirjani, A.K. Enrichment of wheat by zinc fertilizer, mycorrhiza and preharvest drought stress. Turkish J. Field Crop. 2019, 24, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2009; pp. 153–188. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, F.; Ahmad, R.; Waraich, E.; Naeem, M.; Shabbir, R. Nutrient uptake, physiological responses, and yield attributes of wheat (Triticum aestivum L.) exposed to early and late drought stress. J. Plant Nutr. 2012, 35, 961–974. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Ahmad, S.; Saleem, M.F.; Khan, I.H.; Iqbal, R.; Zaheer, M.S.; Haider, I.; Ali, M. Physiological and biochemical assisted screening of wheat varieties under partial rhizosphere drying. Plant Physiol. Biochem. 2017, 116, 150–166. [Google Scholar] [CrossRef]
- Turan, M.; Gulluce, M.; Cakmakci, R.; Oztas, T.; Sahin, F.; Gilkes, R.; Prakongkep, N. The effect of PGPR strain on wheat yield and quality parameters. In Proceedings of the 19th World Congress of Soil Science: Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 209–212. [Google Scholar]
- Du, W.; Sun, Y.; Ji, R.; Zhu, J.; Wu, J.; Guo, H. TiO2 and ZnO nanoparticles negatively affect wheat growth and soil enzyme activities in agricultural soil. J. Environ. Monit. 2011, 13, 822–828. [Google Scholar] [CrossRef]
- Dey, J.K.; Das, S.; Mawlong, L.G. Nanotechnology and its importance in micronutrient fertilization. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 2306–2325. [Google Scholar]
- Zhang, T.; Sun, H.; Lv, Z.; Cui, L.; Mao, H.; Kopittke, P.M. Using synchrotron-based approaches to examine the foliar application of ZnSO4 and ZnO nanoparticles for field-grown winter wheat. J. Agric. Food Chem. 2018, 66, 2572–2579. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.-Y.; Doxey, S.; McLean, J.E.; Britt, D.; Watson, A.; Al Qassy, D.; Jacobson, A.; Anderson, A.J. Remodeling of root morphology by CuO and ZnO nanoparticles: Effects on drought tolerance for plants colonized by a beneficial pseudomonad. Botany 2018, 96, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Defez, R.; Andreozzi, A.; Dickinson, M.; Charlton, A.; Tadini, L.; Pesaresi, P.; Bianco, C. Improved drought stress response in alfalfa plants nodulated by an IAA over-producing Rhizobium strain. Front. Microbiol. 2017, 8, 2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, G.; Conceição, L.F.; Kombrink, E.; Dias, A.C. Xanthone biosynthesis in Hypericum perforatum cells provides antioxidant and antimicrobial protection upon biotic stress. Phytochemistry 2009, 70, 60–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Sharma, S.; Gupta, S.; Kumar, V. Mitigation of abiotic stresses in Lycopersicon esculentum by endophytic bacteria. Environ. Sustain. 2018, 1, 71–80. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munné-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.; Sairam, R.K.; Kushwaha, S.; Singh, T. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Yavas, I.; Unay, A. Effects of zinc and salicylic acid on wheat under drought stress. J. Anim. Plant Sci. 2016, 26, 1012–1101. [Google Scholar]
- Iannone, M.F.; Groppa, M.D.; de Sousa, M.E.; van Raap, M.B.F.; Benavides, M.P. Impact of magnetite iron oxide nanoparticles on wheat (Triticum aestivum L.) development: Evaluation of oxidative damage. Environ. Exp. Bot. 2016, 131, 77–88. [Google Scholar] [CrossRef]
- Arough, Y.K.; Sharifi, R.S.; Sedghi, M.; Barmaki, M. Effect of zinc and bio fertilizers on antioxidant enzymes activity, chlorophyll content, soluble sugars and proline in triticale under salinity condition. Not. Bot. Horti Agrobot Cluj Napoca 2016, 44, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Chiahi, N.; Bouloudenine, M.; El Houda, D.N.; Nesrine, G.; Louhichi, B. L’impact des nanoparticules ZnO sur les paramètres physiologiques et biochimiques chez le blé dur (Triticum turgidum ssp. durum). J. New Sci. 2016, 27, 12. [Google Scholar]
- Wang, C.-J.; Yang, W.; Wang, C.; Gu, C.; Niu, D.-D.; Liu, H.-X.; Wang, Y.-P.; Guo, J.-H. Induction of drought tolerance in cucumber plants by a consortium of three plant growth-promoting rhizobacterium strains. PLoS ONE 2012, 7, e52565. [Google Scholar] [CrossRef]
- Baniaghil, N.; Arzanesh, M.; Ghorbanli, M.; Shahbazi, M. The effect of plant growth promoting rhizobacteria on growth parameters, antioxidant enzymes and microelements of canola under salt stress. J. Appl. Environ. Biol. Sci. 2013, 3, 17–27. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Factorization | Growth Promoting Characters Under Field Trial | |||||
|---|---|---|---|---|---|---|
| Drought Levels | Treatments | PT | Int.L | PH | SL | NSPS |
| D1 = no drought/control | T1 | 356.0 ± 2.3 c | 9.4 ± 0.1 d | 88.2 ± 1.1 g | 12.3 ± 1.5 c | 20.1 ± 1.0 cd |
| T2 | 360.0 ± 1.5 ab | 9.6 ± 0.1 b | 90.1 ± 0.7 c | 13.2 ± 0.6 bc | 21.5 ± 0.5 ab | |
| T3 | 358.0 ± 1.3 bc | 9.5 ± 0.1 c | 89.1 ± 1.2 f | 13.0 ± 2.1 bc | 21.0 ± 0.6 bc | |
| T4 | 362.0 ± 3.5 a | 9.7 ± 0.1 a | 92.4 ± 2.7 a | 14.7 ± 0.6 a | 22.5 ± 0.6 a | |
| D2 = DTS | T1 | 272.0 ± 1.8 j | 8.4 ± 2.5 j | 83.1 ± 1.8 l | 8.2 ± 0.3 e | 17.3 ± 2.5 g |
| T2 | 292.0 ± 2.0 f | 8.6 ± 0.1 h | 86.0 ± 1.6 j | 9.1 ± 1.0 e | 18.3 ± 2.2 fg | |
| T3 | 285.0 ± 1.4 g | 8.3 ± 2.0 k | 84.0 ± 1.2 k | 8.9 ± 0.19 e | 18.1 ± 1.0 fg | |
| T4 | 314.0 ± 2.0 d | 8.7 ± 0.1 g | 87.1 ± 1.2 i | 10.4 ± 2.2 d | 18.7 ± 2.3 ef | |
| D3 = DGS | T1 | 278.0 ± 2.6 i | 8.5 ± 1.4 i | 88.0 ± 2.5 h | 10.4 ± 3.1 d | 15.6 ± 0.6 h |
| T2 | 282.0 ± 2.0 gh | 8.8 ± 0.1 f | 90.0 ± 2.7 d | 11.0 ± 7.1 d | 18.7 ± 2.1 ef | |
| T3 | 280.0 ± 1.6 hi | 8.8 ± 1.7 g | 89.2 ± 2.0 e | 10.9 ± 1.0 d | 18.0 ± 3.2 fg | |
| T4 | 306.0 ± 1.0 e | 9.2 ± 0.1 e | 92.2 ± 3.7 b | 13.7 ± 4.0 ab | 19.6 ± 1.6 de | |
| Experimental Factorization | Yield Promoting Characters Under Field Trial | |||||
|---|---|---|---|---|---|---|
| Drought Levels | Treatments | NGS | 1000 Gwt. | GY | SY | HI |
| D1 = no drought/control | T1 | 40.1 ± 1.8 d | 39.0 ± 1.0 cd | 4.6 ± 1.2 d | 7.9 ± 0.6 d | 36.9 ± 1.5 d |
| T2 | 42.0 ± 1.0 b | 40.0 ± 0.0 ab | 4.9 ± 0.9 b | 8.7 ± 0.2 b | 36.3 ± 1.0 e | |
| T3 | 41.0 ± 1.0 c | 39.5 ± 0.8 bc | 4.7 ± 0.1 c | 8.5 ± 2.0 c | 35.7 ± 1.1 g | |
| T4 | 43.0 ± 1.2 a | 40.5 ± 2.2 a | 5.2 ± 0.1 a | 9.3 ± 2.3 a | 36.0 ± 1.0 fg | |
| D2 = DTS | T1 | 37.0 ± 1.7 g | 36.1 ± 1.2 g | 4.1 ± 0.1 h | 5.2 ± 2.0 l | 44.6 ± 1.4 a |
| T2 | 39.0 ± 1.4 e | 38.0 ± 1.0 e | 4.3 ± 1.6 g | 7.7 ± 1.7 f | 36.0 ± 1.0 ef | |
| T3 | 38.2 ± 2.4 f | 37.0 ± 1.0 f | 4.4 ± 2.2 f | 6.8 ± 1.3 j | 39.4 ± 1.1 b | |
| T4 | 41.0 ± 0.8 cd | 38.5 ± 0.1 de | 4.6 ± 1.2 e | 7.7 ± 0.6 e | 37.3 ± 1.0 c | |
| D3 = DGS | T1 | 32.0 ± 2.0 i | 23.6 ± 2.5 k | 2.6 ± 0.9 l | 6.0 ± 1.2 k | 30.3 ± 1.3 i |
| T2 | 35.4 ± 1.3 h | 27.8 ± 1.7 i | 3.0 ± 0.1 j | 7.2 ± 1.0 h | 29.0 ± 2.1 j | |
| T3 | 35.0 ± 0.9 h | 26.2 ± 0.3 j | 2.8 ± 1.0 k | 7.1 ± 0.3 i | 28.7 ± 1.2 k | |
| T4 | 38.7 ± 1.1 ef | 29.8 ± 1.0 h | 3.6 ± 1.2 i | 7.4 ± 2.1 g | 28.7 ± 1.1 k | |
| Experimental Factorization | Morphological and Grain Quality Traits Under Field Trial | |||||
|---|---|---|---|---|---|---|
| Drought Levels | Treatments | NS−1 | NFT | SpWt. | Gprot. | Gzn |
| D1 = no drought/control | T1 | 3.3 ± 0.1 cd | 10.0 ± 1.3 de | 2.0 ± 0.1 d | 9.3 ± 0.1 l | 12.0 ± 0.1 i |
| T2 | 4.0 ± 0.1 ab | 7.0 ± 2.1 fg | 2.1 ± 1.0 b | 11.2 ± 0.1 e | 16.7 ± 0.1 d | |
| T3 | 3.9 ± 0.2 ab | 8.0 ± 1.2 ef | 2.1 ± 0.2 c | 10.3 ± 0.8 j | 14.5 ± 0.1 e | |
| T4 | 4.3 ± 0.1 a | 5.0 ± 1.2 g | 2.3 ± 0.1 a | 12.2 ± 0.1 a | 18.3 ± 0.2 b | |
| D2 = DTS | T1 | 2.3 ± 0.9 f | 22.0 ± 0.9 a | 1.5 ± 0.1 h | 10.0 ± 0.1 k | 11.0 ± 0.1 k |
| T2 | 2.6 ± 0.3 ef | 14.0 ± 2.2 bc | 1.7 ± 0.1 f | 10.8 ± 0.1 h | 12.4 ± 0.2 h | |
| T3 | 2.6 ± 0.1 ef | 16.0 ± 2.0 b | 1.6 ± 1.4 g | 10.7 ± 0.3 i | 11.6 ± 0.9 j | |
| T4 | 3.3 ± 0.1 cd | 12.0 ± 1.3 cd | 1.8 ± 0.2 e | 10.8 ± 0.1 g | 16.9 ± 0.1 c | |
| D3 = DGS | T1 | 3.0 ± 1.2 de | 11.0 ± 1.4 d | 1.0 ± 0.1 l | 11.0 ± 0.5 f | 9.0 ± 1.1 l |
| T2 | 3.8 ± 0.1 abc | 12.0 ± 1.2 cd | 1.1 ± 0.1 j | 11.9 ± 0.1 c | 13.6 ± 0.2 g | |
| T3 | 3.6 ± 0.1 bc | 11.0 ± 1.3 d | 1.0 ± 0.1 k | 11.4 ± 0.1 d | 14.3 ± 0.9 f | |
| T4 | 4.0 ± 0.1 ab | 11.0 ± 1.7 d | 1.2 ± 0.1 i | 12.1 ± 0.2 b | 19.5 ± 0.1 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muhammad, F.; Raza, M.A.S.; Iqbal, R.; Zulfiqar, F.; Aslam, M.U.; Yong, J.W.H.; Altaf, M.A.; Zulfiqar, B.; Amin, J.; Ibrahim, M.A. Ameliorating Drought Effects in Wheat Using an Exclusive or Co-Applied Rhizobacteria and ZnO Nanoparticles. Biology 2022, 11, 1564. https://doi.org/10.3390/biology11111564
Muhammad F, Raza MAS, Iqbal R, Zulfiqar F, Aslam MU, Yong JWH, Altaf MA, Zulfiqar B, Amin J, Ibrahim MA. Ameliorating Drought Effects in Wheat Using an Exclusive or Co-Applied Rhizobacteria and ZnO Nanoparticles. Biology. 2022; 11(11):1564. https://doi.org/10.3390/biology11111564
Chicago/Turabian StyleMuhammad, Faqeer, Muhammad Aown Sammar Raza, Rashid Iqbal, Faisal Zulfiqar, Muhammad Usman Aslam, Jean Wan Hong Yong, Muhammad Ahsan Altaf, Bilal Zulfiqar, Jawad Amin, and Muhammad Arif Ibrahim. 2022. "Ameliorating Drought Effects in Wheat Using an Exclusive or Co-Applied Rhizobacteria and ZnO Nanoparticles" Biology 11, no. 11: 1564. https://doi.org/10.3390/biology11111564