
Genetic Characterization and Alternative Preservation Ways of Locally Adapted Sheep Breeds: Cases of Private and Public Sheep Sectors in Tunisia and Italy
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Description
2.2. DNA Extraction and Genotyping
2.3. Statistical Analysis
3. Results and Discussion
3.1. Microsatellite Panel Variability
3.2. Breed Variability
3.3. Issues of Genetic Variability Analysis and Relative Conservation Decisions
3.3.1. Issue of the Genetic Variability Analysis: Case of Tunisian Local Sheep Breeds
3.3.2. Issue of the Genetic Variability Analysis: Case of the Italian Local Sheep Breeds
- The transhumant system, mainly used for the large-sized and long-legged FOZ and LAM breeds, reared with large flocks grazing on plains and coastal areas during winter, and displacing to highland pastures in summer,
3.3.3. Diversity Contribution of the Private and Public Sheep Breeding Sectors
3.4. Breed Relationships and Differentiation
3.4.1. Factorial Correspondence Analysis (FCA)
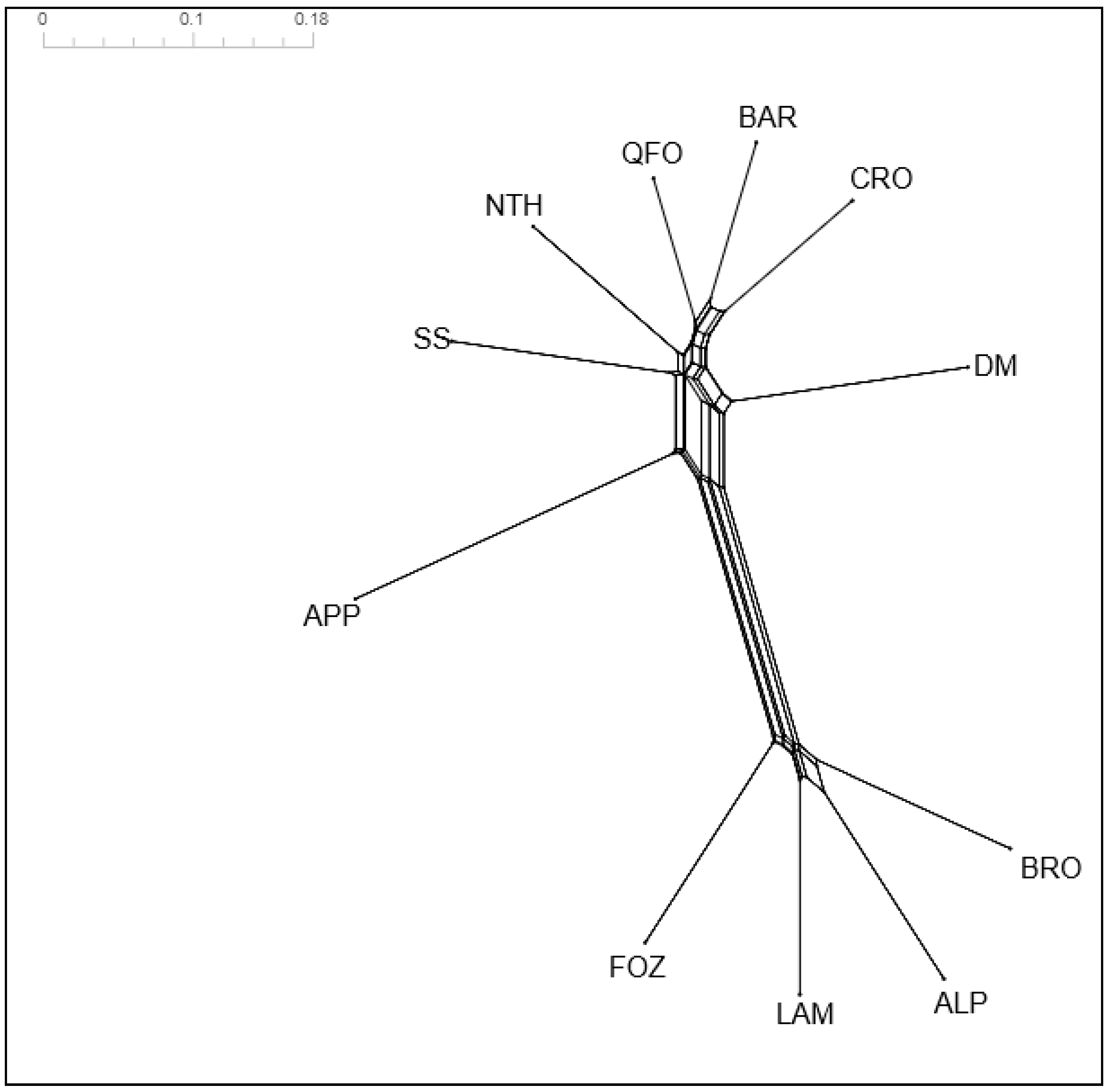
3.4.2. POSA Distances and Neighbor Network Analysis
3.4.3. Genetic Structure Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; Scherf, B.D., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2015; ISBN 9789251088203. Available online: http://www.fao.org/3/a-i4787e (accessed on 22 April 2022).
- FAO. Coping with Climate Change–The Roles of Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2015; ISBN 9789251084410. [Google Scholar]
- Cao, J.; Baumung, R.; Boettcher, P.; Scherf, B.; Besbes, B.; Leroy, G. Monitoring and progress in the implementation of the global plan of action on animal genetic resources. Sustainability 2021, 13, 775. [Google Scholar] [CrossRef]
- FAO. Global Plan of Action for Animal Genetic Resources and the Interlaken Declaration; FAO: Rome, Italy, 2007. [Google Scholar]
- FAO. Domestic Animal Diversity Information System (DAD-IS)|Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/dad-is/en/ (accessed on 9 September 2022).
- Pilling, D.; Bélanger, J.; Diulgheroff, S.; Koskela, J.; Leroy, G.; Mair, G.; Hoffmann, I. Global status of genetic resources for food and agriculture: Challenges and research needs. Genet. Resour. 2020, 1, 4–16. [Google Scholar] [CrossRef]
- Sanna, D.; Barbato, M.; Hadjisterkotis, E.; Cossu, P.; Decandia, L.; Trova, S.; Pirastru, M.; Leoni, G.G.; Naitana, S.; Francalacci, P.; et al. The First Mitogenome of the Cyprus Mouflon (Ovis gmelini ophion): New Insights into the Phylogeny of the Genus Ovis. PLoS ONE 2015, 10, e0144257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, E.; Crepaldi, P.; Nicoloso, L.; Lasagna, E.; Sarti, F.M.; Moioli, B.; Napolitano, F.; Carta, A.; Usai, G.; D’Andrea, M.; et al. Genome-wide analysis of Italian sheep diversity reveals a strong geographic pattern and cryptic relationships between breeds. Anim. Genet. 2014, 45, 256–266. [Google Scholar] [CrossRef]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-Wide Analysis of the World’s Sheep Breeds Reveals High Levels of Historic Mixture and Strong Recent Selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [Green Version]
- FAO. Browse by Country and Species|Domestic Animal Diversity Information System (DAD-IS)|Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/dad-is/browse-by-country-and-species/en/ (accessed on 21 April 2022).
- Chalh, A.; El Gazzah, M.; Djemali, M.; Chalbi, N. Genetic and Phenotypic Characterization of the Tunisian Noire De Thibar Lambs on Their Growth Traits. J. Biol. Sci. 2007, 7, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Rekik, M.; Lassoued, N.; Yacoubi, C. Reproductive performances in ewe lambs of the Queue Fine de l’Ouest breed and their D’Man crosses following synchronisation. Small Rumin. Res. 2002, 45, 75–78. [Google Scholar] [CrossRef]
- Ben Sassi-Zaidy, Y.; Mohamed-Brahmi, A.; Chaouch, M.; Maretto, F.; Cendron, F.; Charfi-Cheikhrouha, F.; Ben Abderrazak, S.; Djemali, M.; Cassandro, M. Historical Westward Migration Phases of Ovis aries Inferred from the Population Structure and the Phylogeography of Occidental Mediterranean Native Sheep Breeds. Genes 2022, 13, 1421. [Google Scholar] [CrossRef]
- Bondesan, V.; Tormen, N.; Bittante, G.; Ribeca, C.; Pellettero, E.; Stelletta, C.; Vencato, Y.; Schiavon, E.; Mutinelli, F.; Granato, A.; et al. Conservazione e Caratterizzazione delle Razze Ovine Venete–Programma Bionet. Rete Regionale per la Conservazione e Caratterizzazione della Biodiversità di Interesse Agrario; Gruppo di Lavoro Ovini, Veneto Agricoltura: Legnaro, Italy, 2014. [Google Scholar]
- Djemali, M. Genetic improvement objectives of sheep and goats in Tunisia. Lessons learned. In Proceedings of the Analysis and Definition of the Objectives in Genetic Improvement Programmes in Sheep and Goats. An Economic Approach to Increase Their Profitability. (Options Méditerranéennes: Série A. Séminaires Méditerranéens; n. 43); Gabiña, D., Ed.; Zaragoza CIHEAM: Zaragoza, Spain, 2000; pp. 121–127. [Google Scholar]
- Khaldi, Z.; Haddad, B.; Souid, S.; Rouissi, H.; Ben Gara, A.; Rekik, B. Caracterisation Phenotypique de la Population Ovine du Sud Ouest de la Tunisie. Anim. Genet. Resour. 2011, 49, 1–8. [Google Scholar] [CrossRef]
- Ben Sassi-Zaidy, Y.; Maretto, F.; Zanetti, E.; Hajji, G.M.; Charfi-Cheikrouha, F.; Cassandro, M. Genetic structure and variability within and among populations of the fat-tailed Barbarine sheep breed using microsatellites markers. Afr. J. Biotechnol. 2014, 13, 44–54. [Google Scholar] [CrossRef]
- OTD–Acceuil. Available online: http://www.otd.nat.tn/fr/ (accessed on 17 April 2022).
- OTD; OTD Sheep Management, Office des Terres Domaniales, Direction de Production Animale, Tunis, Tunisia. Personal communication, 2016.
- Ben Sassi-Zaidy, Y. Caractérisation Génétique des Races Ovines en Tunisie. Ph.D. Thesis, Université de Tunis El Manar, Faculté des Sciences de Tunis, Tunis, Tunisie, 2016. [Google Scholar]
- Dalvit, C.; De Marchi, M.; Zanetti, E.; Cassandro, M. Genetic variation and population structure of Italian native sheep breeds undergoing in situ conservation. J. Anim. Sci. 2009, 87, 3837–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISAG Standing Committee. Applied Genetics in Sheep and Goats. Workshop Report; ISAG conference: Xi’an, China, 2014. [Google Scholar]
- Dalvit, C.; Saccà, E.; Cassandro, M.; Gervaso, M.; Pastore, E.; Piasentier, E. Genetic diversity and variability in Alpine sheep breeds. Small Rumin. Res. 2008, 80, 45–51. [Google Scholar] [CrossRef]
- Ben Sassi-Zaidy, Y.; Maretto, F.; Charfi-cheikhrouha, F.; Mohamed-Brahmi, A.; Cassandro, M. Contribution of microsatellites markers in the clarification of the origin, genetic risk factors, and implications for conservation of Tunisian native sheep breeds. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Ben Sassi-Zaidy, Y.; Mohamed-Brahmi, A.; Nouairia, G.; Charfi-Cheikhrouha, F.; Djemali, M.; Cassandro, M. Genetic Variability and Population Structure of the Tunisian Sicilo-Sarde Dairy Sheep Breed Inferred from Microsatellites Analysis. Genes 2022, 13, 304–317. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.; Wills, D.; Shipley, P. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, Logiciel Sous Windows TM Pour la Génétique des Populations. Laboratoire Génome, Populations, Interaction. CNRS UMR 5000, Université de Montpellier II, Montpellier, France. Available online: http://www.genetix.univ-montp2.fr/genetix/intro.htm (accessed on 10 April 2022).
- Kalinowski, S.T. hp-rare 1.0: A computer program for performing rarefaction on measures of allelic richness. Mol. Ecol. Notes 2005, 5, 187–189. [Google Scholar] [CrossRef]
- Dieringer, D.; Schlötterer, C. Microsatellite analyser (MSA): A platform independent analysis tool for large microsatellite data sets. Mol. Ecol. Notes 2003, 3, 167–169. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Gutierrez, J.P.; Royo, L.J.; Alvarez, I.; Goyache, F. MolKin v2.0: A Computer Program for Genetic Analysis of Populations Using Molecular Coancestry Information. J. Hered. 2005, 96, 718–721. [Google Scholar] [CrossRef] [Green Version]
- Caballero, A.; Toro, M.A. Analysis of genetic diversity for the management of conserved subdivided populations. Conserv. Genet. 2002, 3, 289–299. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- Olschewsky, A.; Hinrichs, D. An overview of the use of genotyping techniques for assessing genetic diversity in local farm animal breeds. Animals 2021, 11, 2016. [Google Scholar] [CrossRef]
- Salinas-Rios, T.; Hernández-Bautista, J.; Mariscal-Méndez, A.; Aquino-Cleto, M.; Martínez-Martínez, A.; Rodríguez-Magadán, H.M. Genetic characterization of a sheep population in Oaxaca, Mexico: The chocholteca creole. Animals 2021, 11, 1172. [Google Scholar] [CrossRef]
- Castillo-Rodríguez, R.G.; Segura-León, O.L.; Hernández-Rodríguez, M.; Serna-Lagunes, R.; Salinas-Ruiz, J.; Salazar-Ortiz, J. Genetic Diversity of Creole Sheep Managed by Indigenous Communities of the Central Region of Veracruz, Mexico. Animals 2022, 12, 456. [Google Scholar] [CrossRef]
- Marina, H.; Suarez-Vega, A.; Pelayo, R.; Gutiérrez-Gil, B.; Reverter, A.; Esteban-Blanco, C.; Arranz, J.J. Accuracy of Imputation of Microsatellite Markers from a 50K SNP Chip in Spanish Assaf Sheep. Animals 2021, 11, 86. [Google Scholar] [CrossRef]
- Laoun, A.; Harkat, S.; Lafri, M.; Gaouar, S.B.S.; Belabdi, I.; Ciani, E.; De Groot, M.; Blanquet, V.; Leroy, G.; Rognon, X.; et al. Inference of breed structure in farm animals: Empirical comparison between SNP and microsatellite performance. Genes 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Eusebi, P.G.; Martinez, A.; Cortes, O. Genomic Tools for Effective Conservation of Livestock Breed Diversity. Diversity 2020, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, S.J.; Aldridge, C.L.; Oyler-Mccance, S.J. An empirical comparison of population genetic analyses using microsatellite and SNP data for a species of conservation concern. BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, M.G.C.D.; Carvalho, M.R.S.; Egito, A.A.; Steinberg, R.S.; Bruneli, F.Â.T.; Machado, M.A.; Santos, F.C.; Rosse, I.C.; Fonseca, P.A.S. Genetic diversity and population genetic structure of a guzerá (Bos indicus) meta-population. Animals 2021, 11, 1125. [Google Scholar] [CrossRef] [PubMed]
- Bittante, G.; Pellattiero, E.; Cecchinato, A.; Tagliapietra, F.; Sturaro, E.; Ramanzin, M.; Pazzola, M.; Massimo Vacca, G.; Schiavon, S. Performance, carcass conformation and meat quality of suckling, weaned and heavy lambs, and culled fattened ewes of autochthonous alpine sheep breeds. Ital. J. Anim. Sci. 2021, 20, 970–984. [Google Scholar] [CrossRef]
- Bozzi, R.; Degl’Innocenti, P.; Rivera Diaz, P.; Nardi, L.; Crovetti, A.; Sargentini, C.; Giorgetti, A. Genetic characterization and breed assignment in five Italian sheep breeds using microsatellite markers. Small Rumin. Res. 2009, 85, 50–57. [Google Scholar] [CrossRef]
- Lasagna, E.; Landi, V.; Bianchi, M.; Martínez Martínez, A.; Sarti, F.M. Genetic characterization of Appenninica sheep breed by microsatellites. Ital. J. Anim. Sci. 2009, 8, 96–98. [Google Scholar] [CrossRef]
- Elloumi, M.; Alary, V.; Selmi, S. Policies and strategies of livestock farmers in Sidi Bouzid Governorate (central Tunisia). Afr. Contemp. 2006, 219, 63–79. [Google Scholar] [CrossRef]
- FAO. Characterization and Value Addition to Local Breeds and Their Products in the Near East and North Africa–Regional Workshop, Rabat, Morocco, 19–21 November 2012. Animal Production and Health Report No. 3; FAO: Rome, Italy, 2014; ISBN 9789251081808. [Google Scholar]
- Ghernouti, N.; Bodinier, M.; Ranebi, D.; Maftah, A.; Petit, D.; Gaouar, S.B.S. Control Region of mtDNA identifies three migration events of sheep breeds in Algeria. Small Rumin. Res. 2017, 155, 66–71. [Google Scholar] [CrossRef]
- Wanjala, G.; Bagi, Z.; Kusza, S. Meta-Analysis of Mitochondrial DNA Control Region Diversity to Shed Light on Phylogenetic Relationship and Demographic History of African Sheep (Ovis aries) Breeds. Biology 2021, 10, 762. [Google Scholar] [CrossRef]
- Horsburgh, K.A.; Beckett, D.B.; Gosling, A.L. Maternal Relationships among Ancient and Modern Southern African Sheep: Newly Discovered Mitochondrial Haplogroups. Biology 2022, 11, 428. [Google Scholar] [CrossRef] [PubMed]
- Leroy, G.; Besbes, B.; Boettcher, P.; Hoffmann, I.; Pilling, D.; Baumung, R.; Scherf, B. Factors and determinants of animal genetic resources management activities across the world. Livest. Sci. 2016, 189, 70–77. [Google Scholar] [CrossRef]
- Kawęcka, A.; Pasternak, M.; Miksza-Cybulska, A.; Puchała, M. Native Sheep Breeds in Poland-Importance and Outcomes of Genetic Resources Protection Programmes. Animals 2022, 12, 1510. [Google Scholar] [CrossRef]
- Agriculture Ministry. Ministère de l’Agriculture, des Ressources Hydroliques et de la Pêche. Enquête sur les Structures des Exploitations Agricoles (2004–2005); General Direction of Studies and Agricultural Developpement: Tunis, Tunisia, 2006.
- Rekik, M.; Aloulou, R.; Ben Hamouda, M. Small ruminant breeds of Tunisia. In Characterisation of Small Ruminant Breeds in West Asia and North Africa, (2) North Africa; Iniguez, L., Ed.; International Centre for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 2005; pp. 91–140. ISBN 92-9127-177-3. [Google Scholar]
- OEP; Office d’élevage et des paturages, Tunis, Tunisia. Personal communication, 2022.
- Ben Yedder, M. Results of the Sheep Growth Monitoring Program for the Years 2019 and 2020. Office d’élevage et des Paturages (OEP), Direction de L’améliorations Génétique. In Proceedings of the Livestock Sector Facing the Risks of Climate Change and Epidemics; “Le Secteur de L’élevage Face aux Risques du Changement Climatique et des Epidémies”, Hammamet, Tunisia, 10–11 June 2021. [Google Scholar]
- Ben Salem, H.; Lassoued, N.; Rekik, M. Merits of the fat-tailed Barbarine sheep raised in different production systems in Tunisia: Digestive, productive and reproductive characteristics. Trop. Anim. Health Prod. 2011, 43, 1357–1370. [Google Scholar] [CrossRef] [PubMed]
- Babelon, E. Carthage et l’Archéologie Punique en Tunisie. Am. J. Archaeol. Hist. Fine Arts 1885, 1, 173–181. [Google Scholar] [CrossRef]
- Atti, N.; Bocquier, F.; Khaldi, G. Performance of the fat-tailed Barbarine sheep in its environment: Adaptive capacity to alternation of underfeeding and re-feeding periods. A review. Anim. Res. 2004, 53, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Bedhiaf-Romdhani, S.; Djemali, M.; Zaklouta, M.; Iniguez, L. Monitoring crossbreeding trends in native Tunisian sheep breeds. Small Rumin. Res. 2008, 74, 274–278. [Google Scholar] [CrossRef]
- Khaldi, G. The Barbary sheep. In Small Ruminant in the Near East, Vol. 3. North Africa; Animal Production and Health paper 74 (A); FAO: Rome, Italy, 1989; pp. 96–135. [Google Scholar]
- Atti, N.; Ben Hammouda, M. Relationship among carcass composition and tail measurements in fat-tailed Barbarine sheep. Small Rumin. Res. 2004, 53, 151–155. [Google Scholar] [CrossRef]
- Jemaa, T.; Huguenin, J.; Moulin, C.H.; Najar, T. Sheep breeding in Central Tunisia: Varied strategies and adaptations to changes in land use. Les systèmes d’élevage de petits ruminants en Tunisie Centrale: Stratégies différenciées et adaptations aux transformations du territoire. Cah. Agric. 2016, 25, 45005. [Google Scholar] [CrossRef] [Green Version]
- Najari, S.; Jaouad, M.; Khaldi, G.; Ben Hammouda, M.; Djemali, M. Traditional small ruminant farming in the arid regions of Tunisia: Farmers’ expertise and sustainable exploitation of pasture resources. Conduite traditionnelle des petits ruminants dans les régions arides tunisiennes: Savoir-faire du berger et exploitat. In Réhabilitation des pâturages et des Parcours en Milieux Méditerranéens; Ferchichi, A., Ed.; Cahiers Options Méditerranéennes, n. 62; Zaragoza CIHEAM: Zaragoza, Spain, 2004; Volume 62, pp. 249–253. [Google Scholar]
- Tunis Sheep–The Livestock Conservancy. Available online: https://livestockconservancy.org/heritage-breeds/heritage-breeds-list/tunis-sheep/ (accessed on 12 May 2022).
- Ben Sassi-Zaidy, Y.; Maretto, F.; Charfi-Cheikrouha, F.; Cassandro, M. Genetic diversity, structure, and breed relationships in Tunisian sheep. Small Rumin. Res. 2014, 119, 52–56. [Google Scholar] [CrossRef]
- Djemali, M.; Bedhiaf-Romdhani, S.; Wurzinger, M.; Iñiguez, L. Impact of butchers and market trends onthe integrity of small ruminant genetic resources. In Proceedings of the 57th Annual Meeting of the European Association for Animal Production, Antalya, Turkey, 17–20 September 2006; pp. 1–6. [Google Scholar]
- Gaouar, S.B.S.; Lafri, M.; Djaout, A.; El-Bouyahiaoui, R.; Bouri, A.; Bouchatal, A.; Maftah, A.; Ciani, E.; Da Silva, A.B. Genome-wide analysis highlights genetic dilution in Algerian sheep. Heredity 2016, 118, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, A.A.; Ata, N.; Benyoucef, M.T.; Djaout, A.; Azzi, N.; Yilmaz, O.; Cemal, İ.; Gaouar, S.B.S. New genetic identification and characterisation of 12 Algerian sheep breeds by microsatellite markers. Ital. J. Anim. Sci. 2018, 17, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Belabdi, I.; Ouhrouch, A.; Lafri, M.; Gaouar, S.B.S.; Ciani, E.; Benali, A.R.; Ould Ouelhadj, H.; Haddioui, A.; Pompanon, F.; Blanquet, V.; et al. Genetic homogenization of indigenous sheep breeds in Northwest Africa. Sci. Rep. 2019, 9, 7920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassoued, N.; Rekik, M. Differences in reproductive efficiency between female sheep of the Queue Fine de l’Ouest purebreed and their first cross with the D’Man. Anim. Res. 2001, 50, 373–381. [Google Scholar] [CrossRef]
- Djemali, M.; Alhadrami, G. Considerations beyond breeding goals in breeding sheep in relation to the environment. Options Mediterr. Ser. A Semin. Mediterr. 1997, 33, 171–174. [Google Scholar]
- Rekik, M.; Ben Salem, I.; Khbou-Khamassi, M.; Letaïef, S.; Chebbi, M. Place des biotechnologies de la reproduction dans la gestion des programmes d ’ amélioration génétique des ovins en Tunisie. Options Méditerranéennes. Séries A. Mediterr. Semin. 2011, 101, 95–101. [Google Scholar]
- OEP. Livestock Sector Indicators Produced by the Information System of the OEP (Office de l’Elevage et des Pasurages). Performance Monitoring Results (2019–2020); OEP: Tunis, Tunisia, 2021.
- Mohamed, A.; Khaldi, S.; Rekik, B.; Khaldi, G. Normal and Residual Milk Yields in Sicilo-Sarde Ewes: Effects of Litter Size and the Weaning Age of Lambs. Res. J. Anim. Sci. 2008, 2, 144–148. [Google Scholar]
- Djemali, M.; Bedhiaf-Romdhani, S.; Iniguez, L.; Inounou, I. Saving threatened native breeds by autonomous production, involvement of farmers organization, research and policy makers: The case of the Sicilo-Sarde breed in Tunisia, North Africa. Livest. Sci. 2009, 120, 213–217. [Google Scholar] [CrossRef]
- Mohamed, A.; Khaldi, R.; Khaldi, G. The adoption of technical and organizational innovations and their impact on the dairy sheep breeding in Tunisia. New Medit 2009, 8, 36–40. [Google Scholar]
- Aloulou, R.; Marnet, P.-G.; M’Sadak, Y. Revue des connaissances sur la micro-filière ovine laitière en Tunisie: État des lieux et perspectives de relance de la race sicilo-sarde. Biotechnol. Agron. Soc. Environ. 2018, 22, 188–198. [Google Scholar] [CrossRef]
- Chniter, M.; Hammadi, M.; Khorchani, T.; Krit, R.; Lahsoumi, B.; Sassi, M.B.; Nowak, R.; Hamouda, M. Ben Phenotypic and seasonal factors influence birth weight, growth rate and lamb mortality in D’man sheep maintained under intensive management in Tunisian oases. Small Rumin. Res. 2011, 99, 166–170. [Google Scholar] [CrossRef]
- Pastore, E.; Fabbris, L. L’allevamento Ovi-Caprino nel Veneto, Analisi e Prospettive Future di un Settore Ricco di Storia; Veneto Agricoltura: Legnaro, Italy, 1999. [Google Scholar]
- Montanari, V. L’allevamento del Bestiame nel Veneto Friuli Venezia Giulia. Ministero dell’Agricoltura e delle Foreste; Ispettorato Agrario Compartimentale delle Venezie: Venetia, Italy, 1954. [Google Scholar]
- Teston, M.; Orsi, M.; Bittante, G.; Cecchinato, A.; Gallo, L.; Gatto, P.; Mota, L.F.M.; Ramanzin, M.; Raniolo, S.; Tormen, A.; et al. Added Value of Local Sheep Breeds in Alpine Agroecosystems. Sustainability 2022, 14, 4698. [Google Scholar] [CrossRef]
- Tormen, N. Conservazione e Valorizzazione delle Risorse Genetiche Animali Autoctone. Master’s Thesis, Scienze e Tecnologie Animali, Universita degli Studi di Padova, Faculty of Agraria, Padova, Italy, 2013. [Google Scholar]
- Ceccobelli, S.; Di Lorenzo, P.; Panella, F.; Lasagna, E.; Sarti, F.M. Morphological and genetic characterisation of Pagliarola breed and its genetic relationships with other three indigenous Italian sheep breeds. Ital. J. Anim. Sci. 2016, 15, 47–54. [Google Scholar] [CrossRef]
- OTD; Office des Terres Domaniales, direction de production animale, Tunis, Tunisia. Personal communication, 2022.
- Djemali, M.; Hamrouni, A. Ingredients and Pathways for Sustainable Sheep Breeding Strategies under Low Input Production Systems: Te Example of Two Distinct Sheep Breeds. J. Vet. Sci. Ani. Husb. 2019, 7, 301. [Google Scholar]
- Ben Abdallah, I.; Hamrouni, A.; Djemali, M. Estimation of genetic parameters and adjustment factors for growth characters of Barbarine high lambs in low input production systems. J. New Sci. 2018, 50, 3042–3047. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Country | Breed | Classification/Origin [20,21] | Vocation/Environment [20,21] | Risk Status [5] | Conservation Program [5] |
|---|---|---|---|---|---|
| Tunisia | BAR | Local rustic fat-tailed/very old (Phoenician) | Typical flavored Meat/well adapted to all climate stage (Humid to Saharan) | Not at Risk | In Situ |
| QFO | Local (Algerian Ouled Djellel) | Meat/Western High altitude (Northern and Eastern pervasive) | Not at Risk | No | |
| CRO | Recent Crossbred (BAR × QFO) | Meat/created in Center (North and East spreading | - | No | |
| NTH | Local (Last century) | Meat/Sub humid to Semi-arid | Not at Risk | No | |
| SS | Local (Last century) | Traditional sheese/Sub humid | Unknown | No | |
| DM | Maghrebian/(Oasis Morocco) | Meat and prolificacy/Saharan | - | No | |
| Italy | ALP | Local | Typical Meat/Alpago mountains | At Risk endangered | In Situ |
| BRO | Local | Typical cheeses/Lissini mountains | At Risk endangered | In Situ | |
| FOZ | Local | Wool/Transhumance in Asiago mountains | At Risk Critical | In Situ | |
| LAM | Local | Typical smoked meat/Semi-wild state: migration between hill, plain and mountains | At Risk endangered | In Situ | |
| APP | Local | Meat/Central Italy | Not at Risk | In Situ |
| Microsatellites | Tunisian Breeds | Italian Breeds | Whole Data | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Locus | Size | Ch | TNA | AR | PIC | TNA | AR | PIC | TNA | AR | PIC |
| Inra023 | 195–221 | 1 | 14 | 11.46 | 0.89 | 16 | 8.31 | 0.84 | 17 | 10.25 | 0.87 |
| Inra063 | 168–208 | 14 | 20 | 12.71 | 0.84 | 25 | 7.82 | 0.79 | 28 | 10.40 | 0.82 |
| OarCP49 | 71–137 | 17 | 29 | 18.02 | 0.90 | 24 | 8.41 | 0.79 | 31 | 13.34 | 0.85 |
| OarFCB304 | 145–201 | 19 | 21 | 11.70 | 0.81 | 18 | 6.43 | 0.69 | 24 | 8.96 | 0.76 |
| OarFCB20 | 85–121 | 2 | 17 | 11.94 | 0.87 | 17 | 7.42 | 0.8 | 19 | 10.26 | 0.84 |
| MAF65 | 113–139 | 15 | 13 | 8.53 | 0.76 | 18 | 5.81 | 0.75 | 21 | 7.75 | 0.75 |
| ILST087 | 134–184 | 6 | 25 | 15.67 | 0.90 | 23 | 9.33 | 0.85 | 26 | 13.17 | 0.88 |
| OarAE119 | 145–185 | 19 | 14 | 10.53 | 0.81 | 20 | 4.82 | 0.66 | 20 | 9.51 | 0.82 |
| MCM527 | 164–190 | 5 | 13 | 8.77 | 0.81 | 15 | 6.34 | 0.76 | 17 | 7.89 | 0.78 |
| MAF214 | 182–262 | 16 | 15 | 6.97 | 0.62 | 36 | 5.71 | 0.6 | 38 | 6.96 | 0.61 |
| OarAE129 | 135–165 | 5 | 8 | 5.67 | 0.60 | 15 | 7.92 | 0.84 | 16 | 5.26 | 0.63 |
| OarCP34 | 93–117 | 3 | 10 | 6.55 | 0.77 | 18 | 6.44 | 0.78 | 18 | 6.62 | 0.77 |
| OarAE54 | 120–152 | 25 | 16 | 10.97 | 0.79 | 19 | 9.31 | 0.86 | 19 | 10.44 | 0.82 |
| TGLA | 125–163 | 12 | 15 | 10.78 | 0.85 | 14 | 7.72 | 0.82 | 16 | 9.67 | 0.84 |
| URB | 159–211 | 13 | 22 | 10.95 | 0.84 | 20 | 9.91 | 0.88 | 24 | 9.93 | 0.86 |
| CSRD | 208–262 | 14 | 26 | 10.88 | 0.85 | 24 | 9.11 | 0.83 | 28 | 9.74 | 0.84 |
| HSC | 260–296 | 20 | 19 | 9.86 | 0.86 | 19 | 8.92 | 0.85 | 21 | 9.33 | 0.85 |
| Average | - | - | 17.47 | 10.70 | 0.810 | 20.06 | 7.63 | 0.788 | 22.53 | 9.38 | 0.80 |
| SD | - | - | 5.75 | 3.07 | 0.086 | 5.29 | 1.49 | 0.076 | 6.07 | 2.09 | 0.08 |
| Breed | N | AR | PAR | He | Ho | FIS | HWE | GD Loss (−)/Gain (+)/Conservation Decision * |
|---|---|---|---|---|---|---|---|---|
| BAR | 64 | 10.05 ± 3.51 | 0.34 | 0.83 ± 0.10 | 0.75 ± 0.18 | 0.073 ± 0.062 | 1 *** | +0.227/No |
| QFO | 41 | 10.40 ± 3.05 | 0.30 | 0.82 ± 0.03 | 0.74 ± 0.14 | 0.101 ± 0.143 | 2 *** | +0.119/No |
| CRO | 30 | 09.73 ± 3.01 | 0.25 | 0.80 ± 0.10 | 0.68 ± 0.17 | 0.139 ± 0.192 | 2 *** | +0.213/No |
| NTH | 41 | 09.29 ± 2.34 | 0.16 | 0.82 ± 0.04 | 0.75 ± 0.15 | 0.071 ± 0.121 | 1 *** | +0.123/No |
| SS | 45 | 10.05 ± 2.95 | 0.38 | 0.82 ± 0.02 | 0.73 ± 0.13 | 0.102 ± 0.103 | 2 *** | −0.064/Cons |
| DM | 28 | 10.38 ± 3.40 | 0.45 | 0.82 ± 0.01 | 0.72 ± 0.17 | 0.123 ± 0.138 | 1 *** | −0.041/Cons |
| ALP | 250 | 09.01 ± 1.71 | 0.36 | 0.77 ± 0.11 | 0.67 ± 0.16 | 0.134 ± 0.132 | 12 *** | −0.280/Cons |
| BRO | 186 | 08.97 ± 1.72 | 0.38 | 0.79 ± 0.08 | 0.70 ± 0.10 | 0.109 ± 0.112 | 8 *** | −0.372/Cons |
| FOZ | 118 | 08.91 ± 1.83 | 0.33 | 0.81 ± 0.09 | 0.72 ± 0.15 | 0.107 ± 0.150 | 4 *** | −0.493/Cons |
| LAM | 141 | 09.01 ± 1.73 | 0.29 | 0.80 ± 0.08 | 0.72 ± 0.11 | 0.101 ± 0.101 | 6 *** | −0.470/Cons |
| APP | 31 | 07.56 ± 2.41 | 0.21 | 0.76 ± 0.08 | 0.66 ± 0.14 | 0.119 ± 0.184 | 3 *** | +0.151/No |
| Sheep Populations | N | He | Ho | FIS | Loss (−)/Gain (+) of GD | C&T Conservation Decision | ||
|---|---|---|---|---|---|---|---|---|
| Tunisian sheep breeds | NTH | NTH | 26 | 0.800 ± 0.091 | 0.750 ± 0.158 | 0.063 | −0.130 | Cons |
| NTHO | 15 | 0.800 ± 0.101 | 0.747 ± 0.172 | 0.068 | −0.056 | Cons | ||
| SS | SS | 30 | 0.812 ± 0.097 | 0.738 ± 0.133 | 0.092 | −0.384 | Cons | |
| SSO | 15 | 0.808 ± 0.123 | 0.706 ± 0.198 | 0.129 | −0.110 | Cons | ||
| QFO | QFO | 34 | 0.828 ± 0.068 | 0.741 ± 0.134 | 0.107 | −0.284 | Con | |
| QFOO | 15 | 0.795 ± 0.149 | 0.738 ± 0.236 | 0.076 | 0.017 | No | ||
| BTR | BTR | 31 | 0.804 ± 0.094 | 0.739 ± 0.137 | 0.083 | 0.112/−0.034 * | No/Cons * | |
| BTRO | 17 | 0.804 ± 0.090 | 0.755 ± 0.156 | 0.054 | 0.012/−0.068 * | No/Cons * | ||
| BTN | BTN | 31 | 0.798 ± 0.103 | 0.762 ± 0.152 | 0.047 | 0.161/0.251 * | No | |
| BTNO | 15 | 0.800 ± 0.109 | 0.799 ± 0.161 | 0.012 | 0.108/0.056 * | No | ||
| Venetian sheep breeds | ALP | ALPP | 186 | 0.773 ± 0.113 | 0.662 ± 0.162 | 0.139 | −4.922 | Cons |
| ALPI | 64 | 0.717 ± 0.158 | 0.704 ± 0.190 | 0.025 | 0.051 | No | ||
| BRO | BROP | 128 | 0.793 ± 0.076 | 0.696 ± 0.085 | 0.109 | −5.763 | Cons | |
| BROI | 58 | 0.740 ± 0.094 | 0.712 ± 0.140 | 0.035 | 0.549 | No | ||
| FOZ | FOZP | 76 | 0.801 ± 0.086 | 0.730 ± 0.176 | 0.087 | −6.080 | Cons | |
| FOZI | 42 | 0.760 ± 0.097 | 0.707 ± 0.141 | 0.073 | −0.167 | Cons | ||
| LAM | LAMP | 100 | 0.740 ± 0.094 | 0.712 ± 0.140 | 0.035 | 0.549 | No | |
| LAMI | 41 | 0.784 ± 0.063 | 0.747 ± 0.131 | 0.036 | −1.377 | Cons | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Sassi-Zaidy, Y.; Mohamed-Brahmi, A.; Aloulou, R.; Charfi-Cheikhrouha, F.; Cendron, F.; Tormen, N.; Cassandro, M. Genetic Characterization and Alternative Preservation Ways of Locally Adapted Sheep Breeds: Cases of Private and Public Sheep Sectors in Tunisia and Italy. Biology 2022, 11, 1623. https://doi.org/10.3390/biology11111623
Ben Sassi-Zaidy Y, Mohamed-Brahmi A, Aloulou R, Charfi-Cheikhrouha F, Cendron F, Tormen N, Cassandro M. Genetic Characterization and Alternative Preservation Ways of Locally Adapted Sheep Breeds: Cases of Private and Public Sheep Sectors in Tunisia and Italy. Biology. 2022; 11(11):1623. https://doi.org/10.3390/biology11111623
Chicago/Turabian StyleBen Sassi-Zaidy, Yousra, Aziza Mohamed-Brahmi, Rafik Aloulou, Faouzia Charfi-Cheikhrouha, Filippo Cendron, Nicola Tormen, and Martino Cassandro. 2022. "Genetic Characterization and Alternative Preservation Ways of Locally Adapted Sheep Breeds: Cases of Private and Public Sheep Sectors in Tunisia and Italy" Biology 11, no. 11: 1623. https://doi.org/10.3390/biology11111623