
Effect of Potassium Deficiency on Physiological Responses and Anatomical Structure of Basil, Ocimum basilicum L.

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Application of Potassium Treatments
- To apply 1 mM K+, the KNO3 concentration was reduced from 1.25 mM to 0.75 mM, and 0.5 mM NaNO3 was added;
- To apply 0.5 mM K+, the KNO3 concentration was reduced from 1.25 mM to 0.25 mM, and 1 mM NaNO3 was added; and
- To apply total potassium deficiency, KNO3 salt (1.25 mM) and KH2PO4 salt (0.25 mM) were replaced by NaNO3 (1.25 mM) and NaH2PO4 (0.25 mM), respectively.
2.3. Analytical Techniques
2.4. Preparation of Anatomical Sections
2.5. Methods of Statistical Analysis
3. Results
3.1. Effect of Potassium Deficiency on Plant Growth and Development
3.1.1. Plant Aspect
3.1.2. Morphological Parameters
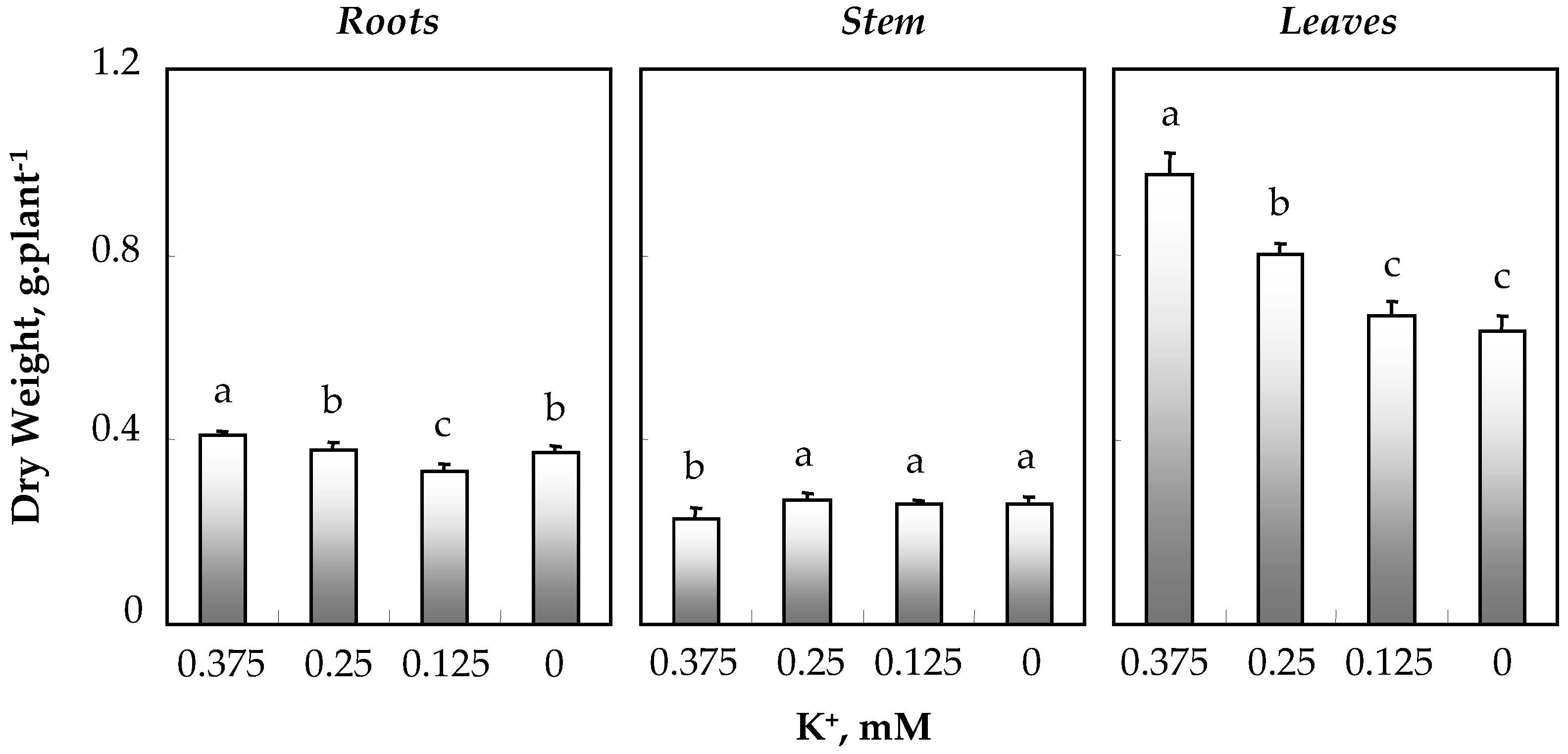
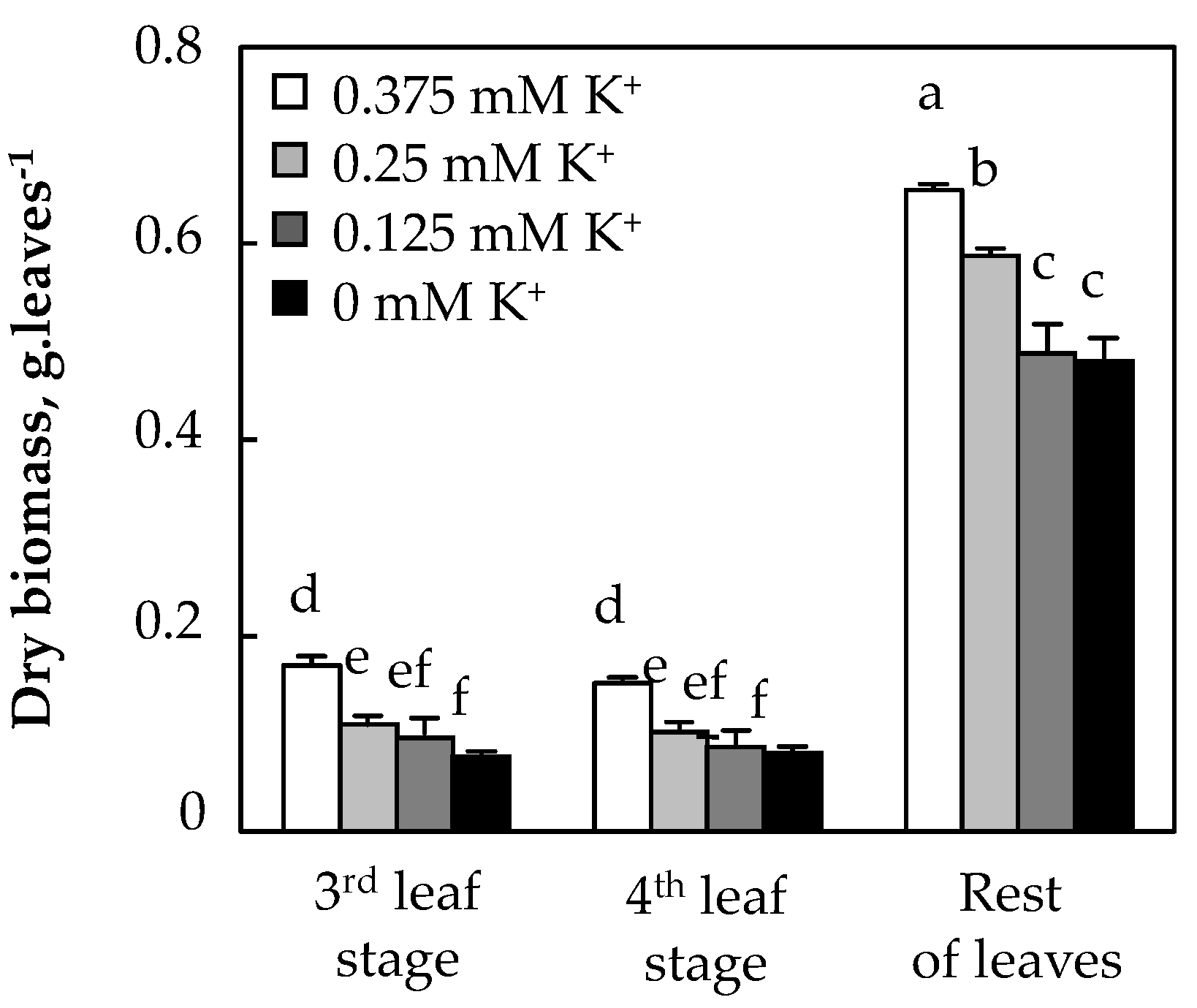
3.1.3. Biomass Production
3.1.4. Potassium Concentrations and Amounts
3.2. Effect of K+ Concentration in the Medium on Chlorophylls and Carotenoids
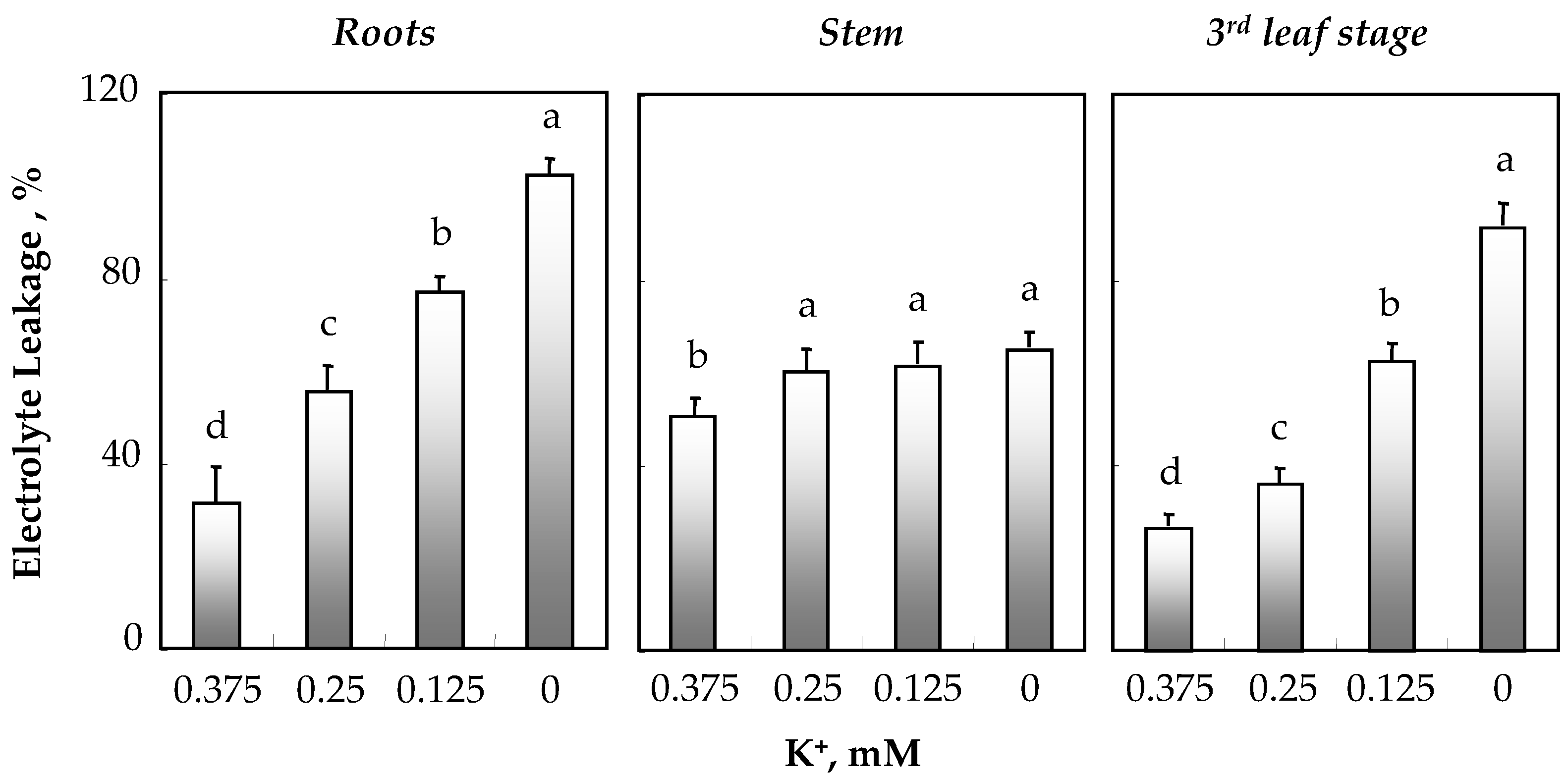
3.3. Effect of K+ Deficiency on Membrane Integrity
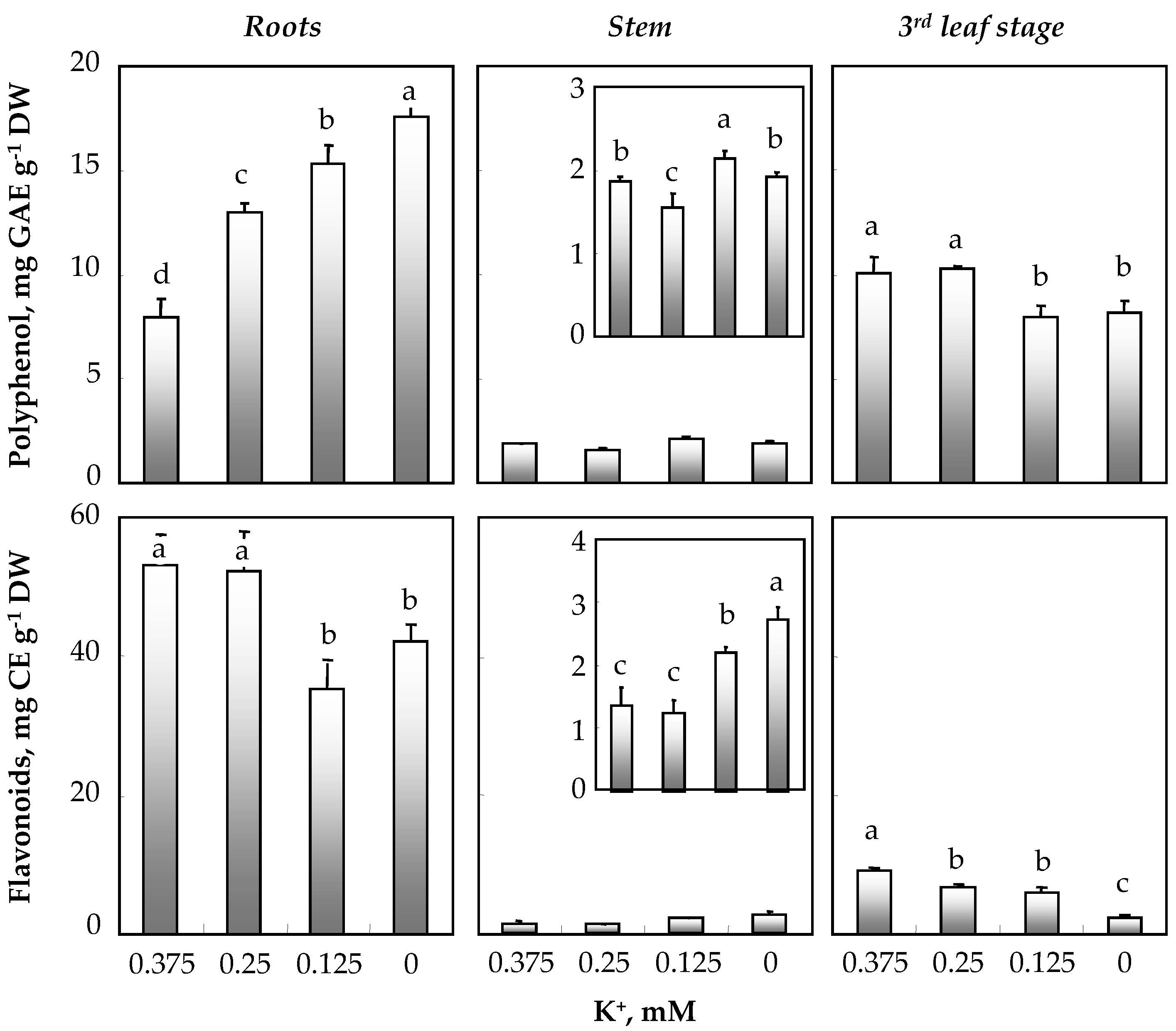
3.4. Effect of K+ Concentration in the Medium on Total Polyphenols and Flavonoids
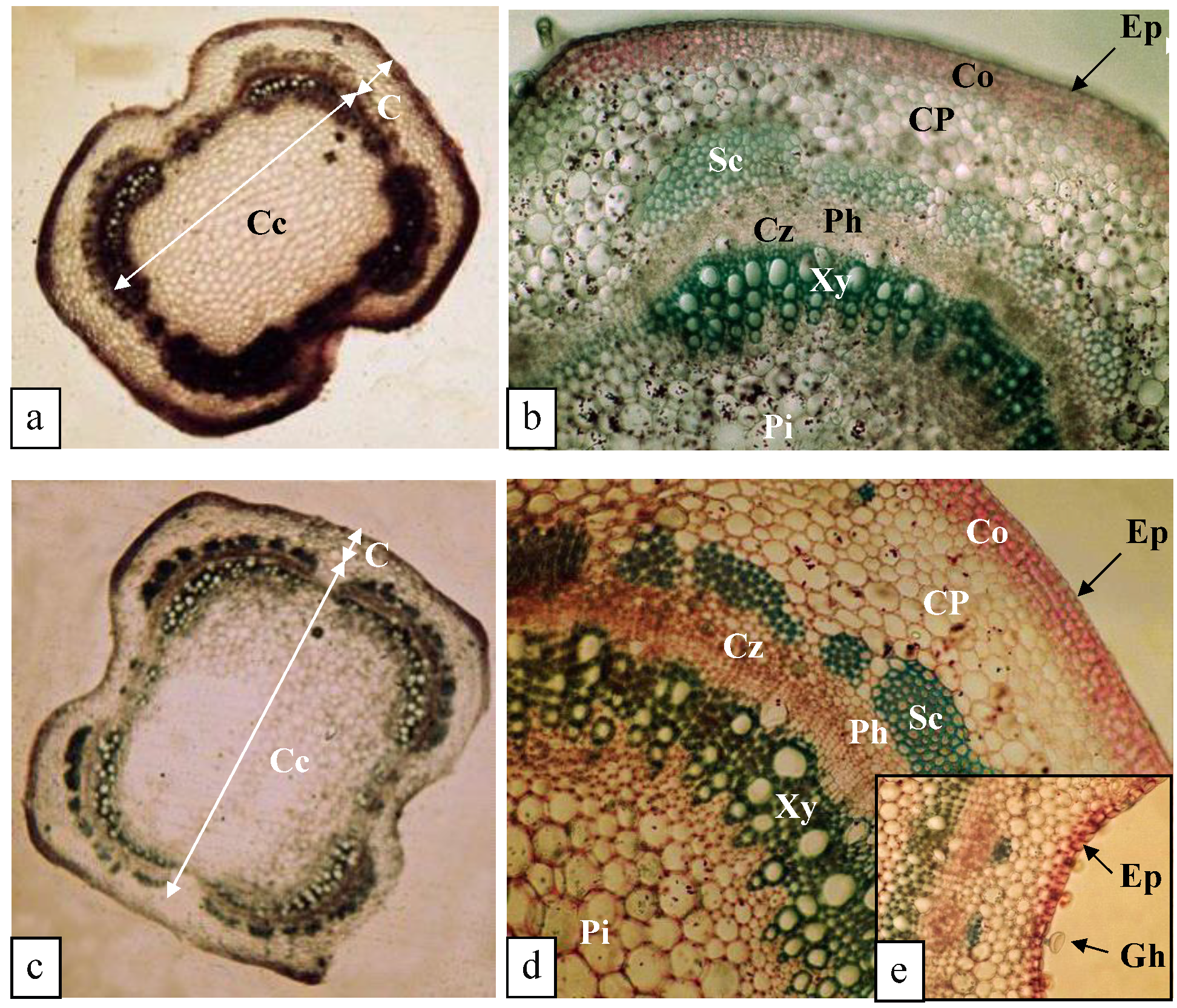
3.5. Effect of K+ Deficiency on the Anatomical Structure of O. Basilicum Stems
3.5.1. Structure of the Stem (0.375 mM K+)
3.5.2. Structure of the Stem (0.25 mM K+)
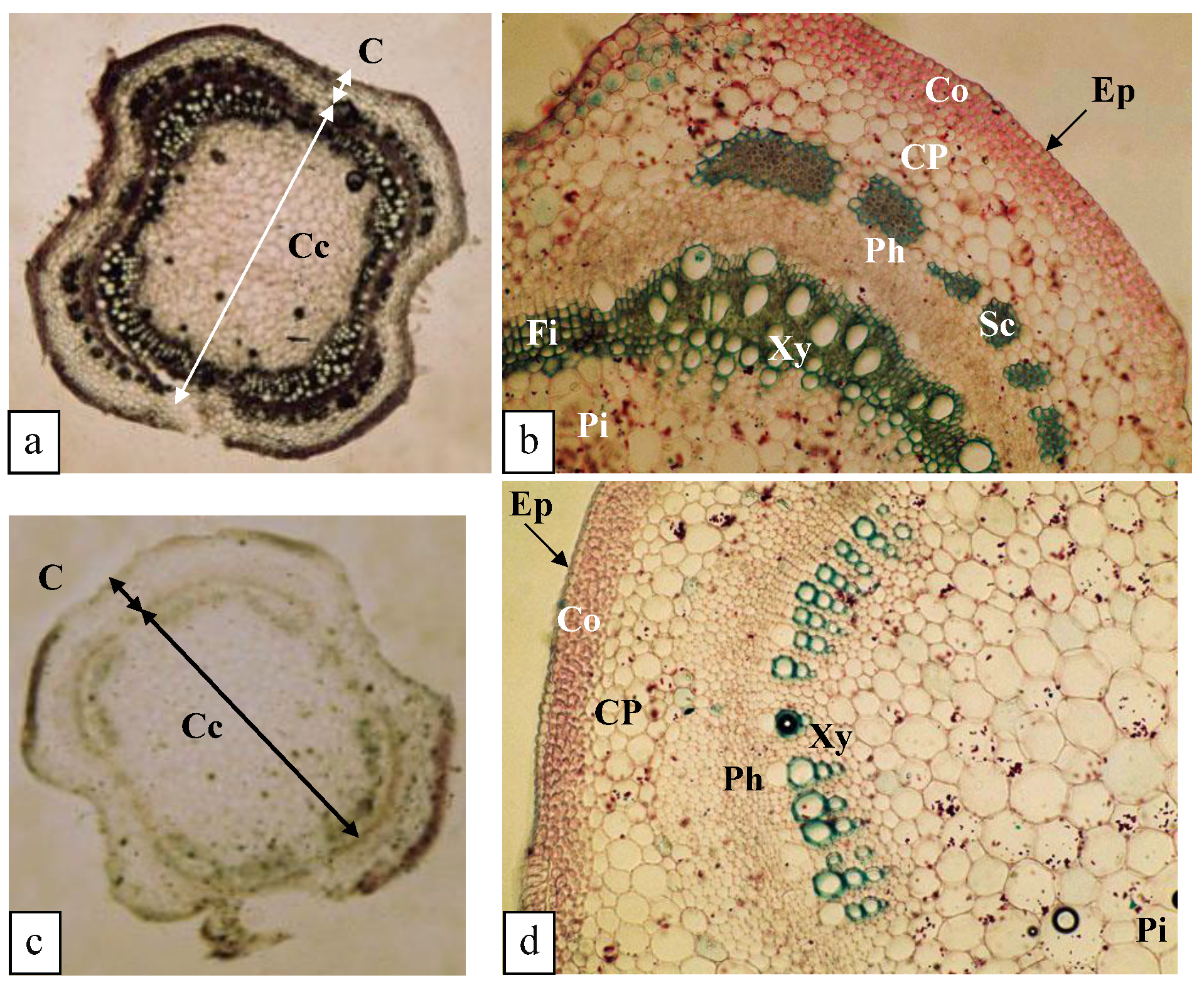
3.5.3. Structure of the Stem (0.125 mM K+)
3.5.4. Structure of the Stem (0 mM K+)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hafsi, C.; Romero-Puertas, M.C.; del Rio, L.A.; Sandalio, L.M.; Abdelly, C. Differential antioxidative response in barley leaves subjected to the interactive effects of salinity and potassium deprivation. Plant Soil 2010, 334, 449–460. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Borhannuddin-Bhuyan, M.H.M.; Nahar, K.; Shahadat-Hossain, M.; Al Mahmud, J.; Shahadat-Hossen, M.; Chowdhury-Masud, A.A.; Moumita Fujita, M. Potassium: A Vital Regulator of Plant Responses and Tolerance to Abiotic Stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Damalas, C.A. Improving drought tolerance in sweet basil (Ocimum basilicum) with salicylic acid. Sci. Hortic. 2019, 246, 360–365. [Google Scholar] [CrossRef]
- Barickman, T.C.; Olorunwa, O.J.; Sehgal, A.; Walne, C.H.; Reddy, K.R.; Gao, W. Yield, Physiological Performance, and Phytochemistry of Basil (Ocimum basilicum L.) under Temperature Stress and Elevated CO2 Concentrations. Plants 2021, 10, 1072. [Google Scholar] [CrossRef] [PubMed]
- Attia, H.; Ouhibi, C.; Ellili, A.; Msilini, N.; Bouzaïen, G.; Karray, N.; Lachaâl, M. Analysis of salinity effects on basil leaf surface area, photosynthetic activity, and growth. Acta Physiol. Plant. 2011, 33, 823–833. [Google Scholar] [CrossRef]
- Attia, H.; Alamer, K.; Algethami, B.; Ellouzi, H.; Lachaâl, M. Effects of growth and salinity on some morphological parameters, pigment and flavonoid concentrations in basil. Agrochimica 2020, 64, 121–138. [Google Scholar] [CrossRef]
- Zahedifar, M.; Moosavi, A.A.; Shafigh, M.; Zarei, Z.; Karimian, F. Cadmium accumulation and partitioning in Ocimum basilicum as influenced by the application of various potassium fertilizers. Arch. Agron. Soil Sci. 2016, 62, 663–673. [Google Scholar] [CrossRef]
- Borges, B.M.M.N.; Flores, R.A.; Almeida, H.J.; Moda, L.R.; Prado, R.M. Macronutrient Omission and the Development and Nutritional Status of Basil in Nutritive Solution. J. Plant Nutr. 2016, 39, 1627–1633. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Ródenas, R.; Lara, A.; Martínez, V.; Rubio, F. The combination of K+ deficiency with other environmental stresses: What is the outcome? Physiol. Plant. 2019, 165, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Martínez, V.; Benito, B.; Rubio, F. Comparison between Arabidopsis and rice for main pathways of K+ and Na+ uptake by roots. Front. Plant Sci. 2016, 7, 992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, D.; Zhao, X.; Le, X.; Jiang, C.; Wang, X.; Yi, H.; Wang, J.; Yu, H. Effects of potassium deficiency on photosynthesis, chloroplast ultrastructure, ROS, and antioxidant activities in maize (Zea mays L.). J. Integr. Agric. 2019, 18, 395–406. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, R.L.L.; de Mello Prado, R.; Felisberto, G.; Checchio, M.V.; Gratão, P.L. Silicon mitigates manganese deficiency stress by regulating the physiology and activity of antioxidant enzymes in sorghum plants. J. Soil Sci. Plant Nutr. 2019, 19, 524–534. [Google Scholar] [CrossRef]
- Hafsi, C.; Bettaib, J.; Falleh, H.; Zorrig, W.; Ksouri, R.; Abdelly, C.; Debez, A. Phenolic accumulation and related antioxidant capacity in stems and roots of the Tunisian extremophile Sulla arnosa as influenced by potassium application under salinity stress. Arab. J. Geosci. 2021, 14, 34. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Bok, V.V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Hafsi, C.; Falleh, H.; Saada, M.; Rabhi, M.; Mkadmini, K.; Ksouri, R.; Abdelly, C.; Smaoui, A. Effects of potassium supply on growth, gas exchange, phenolic composition, and related antioxidant properties in the forage legume Sulla carnosa. Flora 2016, 223, 38–45. [Google Scholar] [CrossRef]
- Arnon, D.I.; Hoagland, D.R. Crops production in artificial solution and in soils with special reference to factors affecting yields and absorption of inorganic nutrients. Soil Sci. 1940, 50, 463–484. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzy. 1987, 148, 350–382. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoids contents in mulberry and their scavenging effects on superoxide radicals. J. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Cicco, N.; Lanorte, M.; Paraggio, M.; Viggiano, M. A reproducible, rapid and inexpensive Folin-Ciocalteu micro-method in determining phenolics of plant methanol extracts. Microchem. J. 2009, 91, 107–110. [Google Scholar] [CrossRef]
- dos Santos Sarah, M.M.; de Mello Prado, R.; de Souza Júnior, J.P.; Teixeira, G.C.M.; dos Santos Duarte, J.C.; de Medeiros, R.L.S. Silicon Supplied via Root or Leaf Relieves Potassium Deficiency Effects in Common Bean. Sci Rep. 2021, 11, 19690. [Google Scholar] [CrossRef] [PubMed]
- Marathe, R.A.; Murkute, A.A.; Dhinesh, B.K. Mineral Nutrient Deficiencies and Nutrient Interactions in Pomegranate. Natl. Acad. Sci. Lett. 2016, 39, 407–410. [Google Scholar] [CrossRef]
- Hetherington, F.M.; Kakkar, M.; Topping, J.F.; Lindsey, K. Gibberellin signaling mediates lateral root inhibition in response to K+-deprivation. Plant Physiol. 2021, 185, 1198–1215. [Google Scholar] [CrossRef]
- Coffey, O.; Bonfield, R.; Corre, F.; Althea Sirigiri, J.; Meng, D.; Fricke, W. Root and cell hydraulic conductivity, apoplastic barriers and aquaporin gene expression in barley (Hordeum vulgare L.) grown with low supply of potassium. Ann. Bot. 2018, 122, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Peuke, A.D.; Jeschke, W.D.; Hartung, W. Flows of elements, ions and abscisic acid in Ricinus communis and site of nitrate reduction under potassium limitation. J. Exp. Bot. 2002, 53, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Demidchik, V. Mechanisms and physiological roles of K+ efflux from root cells. J. Plant Physiol. 2014, 171, 696–707. [Google Scholar] [CrossRef]
- Cui, J.; Abadie, C.; Carroll, A.; Lamade, E.; Tcherkez, G. Responses to K deficiency and waterlogging interact via respiratory and nitrogen metabolism. Plant Cell Environ. 2019, 42, 647–658. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef] [Green Version]
- Quian-Ulloa, R.; Stange, C. Carotenoid biosynthesis and plastid development in plants: The role of light. Int. J. Mol. Sci. 2021, 22, 1184. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Cao, B.; Qi, L.; Yin, L.; Wang, S.; Deng, X. Silicon-moderated K-defciency-induced leaf chlorosis by decreasing putrescine accumulation in sorghum. Ann. Bot. 2016, 118, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.M.; Kwee, E.M.; Niemeyer, E.D. Potassium rate alters the antioxidant capacity and phenolic concentration of basil leaves (Ocimum basilicum L.). Food Chem. 2010, 123, 1235–1241. [Google Scholar] [CrossRef]
- Benslima, W.; Zorrig, W.; Bagues, M.; Abdelly, C.; Hafsi, C. Silicon mitigates potassium deficiency in Hordeum vulgare by improving growth and photosynthetic activity but not through polyphenol accumulation and the related antioxidant potential. Plant Soil 2021, 477, 153–170. [Google Scholar] [CrossRef]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Cabane, M.; Afif, D.; Hawkin, S. Lignins and Abiotic Stresses. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Liu, Y.; Yin, Q.; Dai, B.; Wang, K.L.; Lu, L.; Qaseem, M.F.; Wang, J.; Li, H.; Wu, A.M. The key physiology and molecular responses to potassium deficiency in Neolamarckia cadamba. Ind. Crops Prod. 2021, 162, 113260. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K+, mM | ||||
|---|---|---|---|---|
| Parameter | 0.375 | 0.25 | 0.125 | 0 |
| Root elongation, cm | 29 ± 2 b | 28 ± 1 b | 26 ± 1 c | 32 ± 1 a |
| Leaf area, cm2 · plant−1 | 25 ± 2 a | 16 ± 1 b | 14 ± 1 c | 11 ± 1 d |
| Thickness, cm | 0.02 c | 0.03 b | 0.04 a | 0.03 b |
| K+, mM | ||||
|---|---|---|---|---|
| Pigment | 0.375 | 0.25 | 0.125 | 0 |
| Chlorophyll a | 19.8 ± 1.0 a | 14.1 ± 1.2 b | 10.6 ± 1.2 c | 7.2 ± 1.4 d |
| Chlorophyll b | 7.7 ± 0.7 a | 6.4 ± 0.5 b | 4.7 ± 0.5 c | 4.2 ± 0.6 c |
| Chlorophyll total | 27.0 ± 1.2 a | 18.3 ± 1.7 b | 14.3 ± 1.7 c | 13.3 ± 1.8 c |
| Chl a/b | 2.6 ± 0.2 a | 2.2 ± 0.1 a | 2.3 ± 0.5 a | 1.7 ± 0.2 b |
| Carotenoid (Car) | 6.1 ± 0.2 a | 4.4 ± 0.3 b | 4.2 ± 0.5 b | 3.4 ± 0.3 c |
| Chl/Car | 4.5 ± 0.3 a | 4.2 ± 0.6 a | 3.5 ± 0.8 a | 4.0 ± 0.5 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attia, H.; Rebah, F.; Ouhibi, C.; Saleh, M.A.; Althobaiti, A.T.; Alamer, K.H.; Ben Nasri, M.; Lachaâl, M. Effect of Potassium Deficiency on Physiological Responses and Anatomical Structure of Basil, Ocimum basilicum L. Biology 2022, 11, 1557. https://doi.org/10.3390/biology11111557
Attia H, Rebah F, Ouhibi C, Saleh MA, Althobaiti AT, Alamer KH, Ben Nasri M, Lachaâl M. Effect of Potassium Deficiency on Physiological Responses and Anatomical Structure of Basil, Ocimum basilicum L. Biology. 2022; 11(11):1557. https://doi.org/10.3390/biology11111557
Chicago/Turabian StyleAttia, Houneida, Fedia Rebah, Chayma Ouhibi, Muneera A. Saleh, Ashwaq T. Althobaiti, Khalid H. Alamer, Mouhiba Ben Nasri, and Mokhtar Lachaâl. 2022. "Effect of Potassium Deficiency on Physiological Responses and Anatomical Structure of Basil, Ocimum basilicum L." Biology 11, no. 11: 1557. https://doi.org/10.3390/biology11111557