
Tristetraprolin, Inflammation, and Metabolic Syndrome in Arab Adults: A Case Control Study

 ,
,  , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Study Design and Participants
2.2. MetS Components Classification of Participants
- (1)
- Waist circumference (Central obesity) of >101.6 cm in males and >88.9 cm in females.
- (2)
- Fasting glucose (Hyperglycemia) >5.6 mmol/L.
- (3)
- Low high density lipoprotein cholesterol (HDL-c); for males <1.03 mmol/L and for females <1.30 mmol/L.
- (4)
- Fasting triglycerides (Hypertriglyceridemia) >1.7 mmol/L.
- (5)
- Hypertension; diastolic blood pressure >85 mmHg and/or systolic blood pressure >130 mmHg.
2.3. Anthropometrics and Biochemical Analyses
2.4. Statistical Analysis
3. Results
3.1. General Comparison between MetS Patients and Controls
3.2. Comparison between Male and Female Participants in Both Groups
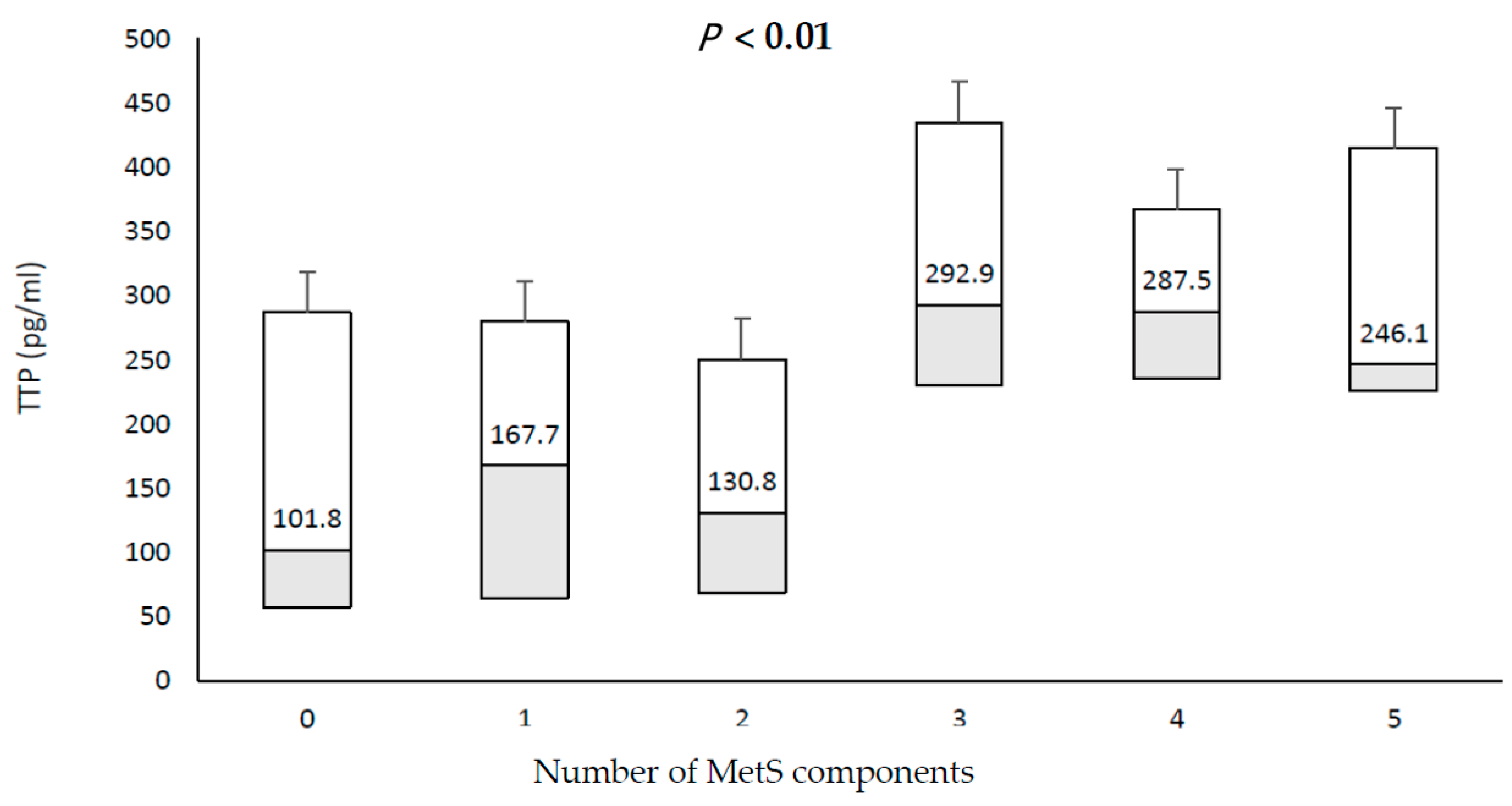
3.3. Relationship between TTP and Individual Components of MetS
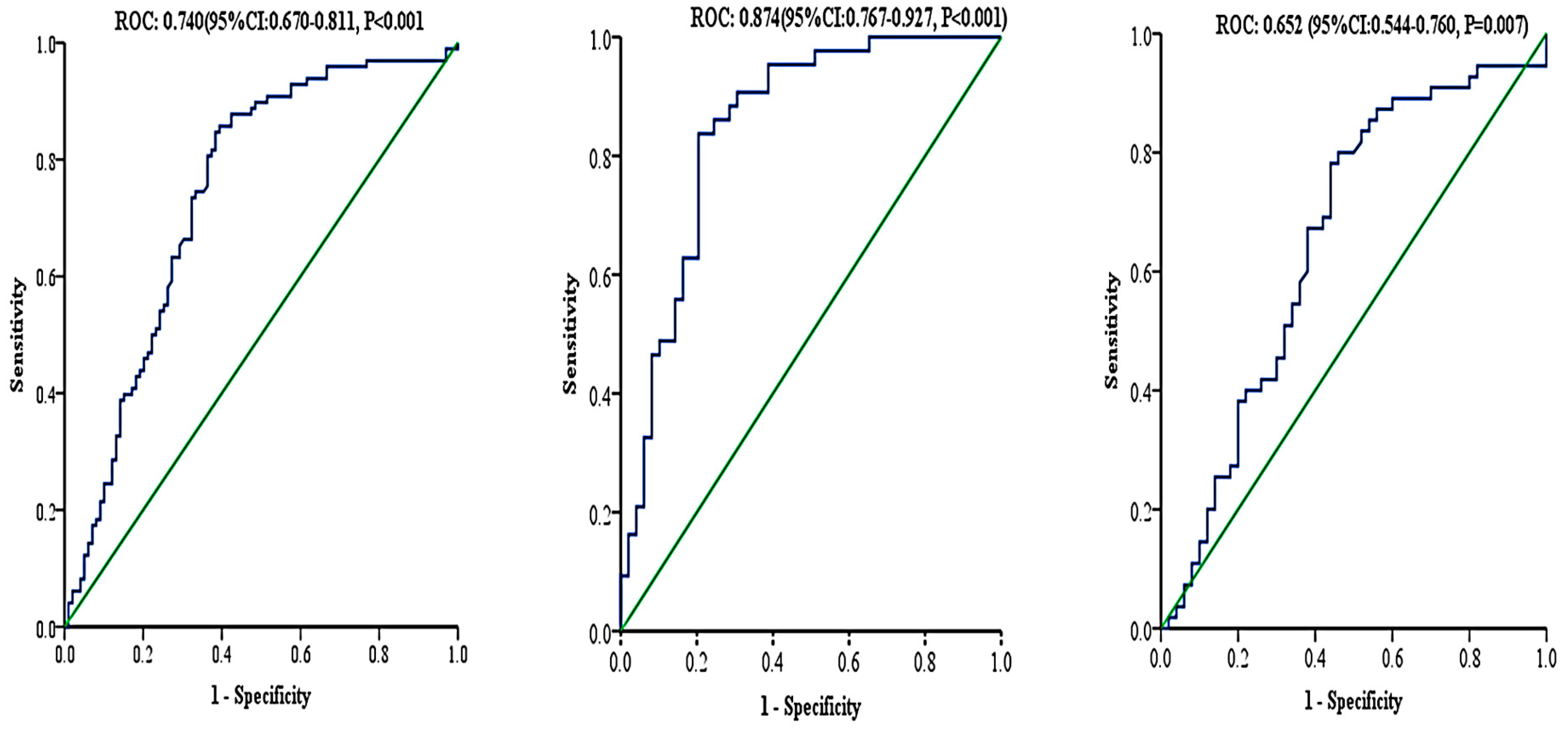
3.4. Associations of TTP with Anthropometrics and Clinical Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Merchant, N. Metabolic syndrome and cardiovascular disease in South Asians. Vasc. Health Risk Manag. 2009, 5, 731–743. [Google Scholar] [CrossRef]
- Iso, H.; Sato, S.; Kitamura, A.; Imano, H.; Kiyama, M.; Yamagishi, K.; Cui, R.; Tanigawa, T.; Shimamoto, T. Metabolic Syndrome and the Risk of Ischemic Heart Disease and Stroke Among Japanese Men and Women. Stroke 2007, 38, 1744–1751. [Google Scholar] [CrossRef] [Green Version]
- Mente, A.; Yusuf, S.; Islam, S.; McQueen, M.J.; Tanomsup, S.; Onen, C.L.; Rangarajan, S.; Gerstein, H.C.; Anand, S.S. Metabolic Syndrome and Risk of Acute Myocardial Infarction: A Case-Control Study of 26,903 Subjects From 52 Countries. J. Am. Coll. Cardiol. 2010, 55, 2390–2398. [Google Scholar] [CrossRef] [Green Version]
- Kassi, E.; Pervanidou, P.; Kaltsas, G.; Chrousos, G. Metabolic syndrome: Definitions and controversies. BMC Med. 2011, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Daghri, N.M.; Clerici, M.; Al-Attas, O.; Forni, D.; Alokail, M.S.; Alkharfy, K.M.; Sabico, S.; Mohammed, A.K.; Cagliani, R.; Sironi, M. A nonsense polymorphism (R392X) in TLR5 protects from obesity but predisposes to diabetes. J. Immunol. 2013, 190, 3716–3720. [Google Scholar] [CrossRef]
- Al-Daghri, N.M.; Al-Attas, O.S.; Alokail, M.S.; Alkharfy, K.M.; Yousef, M.; Sabico, S.L.; Chrousos, G.P. Diabetes mellitus type 2 and other chronic non-communicable diseases in the central region, Saudi Arabia (riyadh cohort 2): A decade of an epidemic. BMC Med. 2011, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Al-Rubeaan, K.; Bawazeer, N.; Al Farsi, Y.; Youssef, A.M.; Al-Yahya, A.A.; AlQumaidi, H.; Al-Malki, B.M.; Naji, K.A.; Al-Shehri, K.; Al Rumaih, F.I. Prevalence of metabolic syndrome in Saudi Arabia—A cross sectional study. BMC Endocr. Disord. 2018, 18, 1–9. [Google Scholar] [CrossRef]
- Lumeng, C.N. Innate immune activation in obesity. Mol. Asp. Med. 2013, 34, 12–29. [Google Scholar] [CrossRef] [Green Version]
- Tornatore, L.; Thotakura, A.K.; Bennett, J.; Moretti, M.; Franzoso, G. The nuclear factor kappa B signaling pathway: Integrating metabolism with inflammation. Trends Cell Biol. 2012, 22, 557–566. [Google Scholar] [CrossRef]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nat. Cell Biol. 2012, 482, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malagón, M.M.; Díaz-Ruiz, A.; Guzmán-Ruiz, R.; Jiménez-Gómez, Y.; Moreno, N.R.; García-Navarro, S.; Vázquez-Martínez, R.; Peinado, J.R. Adipobiology for novel therapeutic approaches in metabolic syndrome. Curr. Vasc. Pharmacol. 2014, 11, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Kaser, A. Gut microbiome, obesity, and metabolic dysfunction. J. Clin. Investig. 2011, 121, 2126–2132. [Google Scholar] [CrossRef]
- Toubal, A.; Treuter, E.; Clément, K.; Venteclef, N. Genomic and epigenomic regulation of adipose tissue inflammation in obesity. Trends Endocrinol. Metab. 2013, 24, 625–634. [Google Scholar] [CrossRef]
- Kraja, A.T.; Chasman, D.I.; North, K.E.; Reiner, A.P.; Yanek, L.R.; Kilpeläinen, T.O.; Smith, J.A.; Dehghan, A.; Dupuis, J.; Johnson, A.D.; et al. Pleiotropic genes for metabolic syndrome and inflammation. Mol. Genet. Metab. 2014, 112, 317–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sell, H.; Habich, C.; Eckel, J. Adaptive immunity in obesity and insulin resistance. Nat. Rev. Endocrinol. 2012, 8, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R.A. Inflammasomes in health and disease. Nat. Cell Biol. 2012, 481, 278–286. [Google Scholar] [CrossRef]
- Ronkina, N.; Shushakova, N.; Tiedje, C.; Yakovleva, T.; Tollenaere, M.A.X.; Scott, A.; Batth, T.S.; Olsen, J.V.; Helmke, A.; Bekker-Jensen, S.H.; et al. The Role of TTP Phosphorylation in the Regulation of Inflammatory Cytokine Production by MK2/3. J. Immunol. 2019, 203, 2291–2300. [Google Scholar] [CrossRef]
- Rappl, P.; Brüne, B.; Schmid, T. Role of Tristetraprolin in the Resolution of Inflammation. Biology 2021, 10, 66. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Yang, S.S.; Vo, M.-T.; Cho, W.J.; Lee, B.J.; Leem, S.-H.; Lee, S.-H.; Cha, H.J.; Park, J.W. Tristetraprolin down-regulates IL-23 expression in colon cancer cells. Mol. Cells 2013, 36, 571–576. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Stoecklin, G.; Van Way, S.; Hinkovska-Galcheva, V.; Guo, R.-F.; Anderson, P.; Shanley, T.P. Tristetraprolin (TTP)-14-3-3 Complex Formation Protects TTP from Dephosphorylation by Protein Phosphatase 2a and Stabilizes Tumor Necrosis Factor-α mRNA. J. Biol. Chem. 2007, 282, 3766–3777. [Google Scholar] [CrossRef] [Green Version]
- Dolicka, D.; Sobolewski, C.; De Sousa, M.C.; Gjorgjieva, M.; Foti, M. mRNA Post-Transcriptional Regulation by AU-Rich Element-Binding Proteins in Liver Inflammation and Cancer. Int. J. Mol. Sci. 2020, 21, 6648. [Google Scholar] [CrossRef]
- Qiu, L.-Q.; Lai, W.S.; Bradbury, A.; Zeldin, D.; Blackshear, P.J. Tristetraprolin (TTP) coordinately regulates primary and secondary cellular responses to proinflammatory stimuli. J. Leukoc. Biol. 2015, 97, 723–736. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.H.; Yoon, N.A.; Vo, M.-T.; Kim, C.W.; Woo, J.M.; Cha, H.J.; Cho, Y.W.; Lee, B.J.; Cho, W.J.; Park, J.W. Tristetraprolin down-regulates IL-17 through mRNA destabilization. FEBS Lett. 2011, 586, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.-G.; Amar, M.J.; Remaley, A.T.; Kwon, J.; Blackshear, P.J.; Wang, P.-Y.; Hwang, P.M. Zinc Finger Protein Tristetraprolin Interacts with CCL3 mRNA and Regulates Tissue Inflammation. J. Immunol. 2011, 187, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Tiedje, C.; Ronkina, N.; Tehrani, M.; Dhamija, S.; Laass, K.; Holtmann, H.; Kotlyarov, A.; Gaestel, M. The p38/MK2-Driven Exchange between Tristetraprolin and HuR Regulates AU–Rich Element–Dependent Translation. PLoS Genet. 2012, 8, e1002977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Chen, X.; Liu, Q.; Zhang, S.; Hu, W. Translation repression via modulation of the cytoplasmic poly(A)-binding protein in the inflammatory response. eLife 2017, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Schott, J.; Reitter, S.; Philipp, J.; Haneke, K.; Schäfer, H.; Stoecklin, G. Translational Regulation of Specific mRNAs Controls Feedback Inhibition and Survival during Macrophage Activation. PLoS Genet. 2014, 10, e1004368. [Google Scholar] [CrossRef]
- Al-Daghri, N.M.; Al-Attas, O.S.; Wani, K.; Sabico, S.; Alokail, M.S. Serum Uric Acid to Creatinine Ratio and Risk of Metabolic Syndrome in Saudi Type 2 Diabetic Patients. Sci. Rep. 2017, 7, 12104. [Google Scholar] [CrossRef] [Green Version]
- Al-Attas, O.; Al-Daghri, N.; Alokail, M.; Abd-Alrahman, S.; Vinodson, B.; Sabico, S. Metabolic Benefits of Six-month Thiamine Supplementation in Patients with and Without Diabetes Mellitus Type 2. Clin. Med. Insights Endocrinol. Diabetes 2014, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Patial, S.; Curtis, A.D.; Lai, W.S.; Stumpo, D.J.; Hill, G.D.; Flake, G.P.; Mannie, M.D.; Blackshear, P.J. Enhanced stability of tristetraprolin mRNA protects mice against immune-mediated inflammatory pathologies. Proc. Natl. Acad. Sci. USA 2016, 113, 1865–1870. [Google Scholar] [CrossRef] [Green Version]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Caracciolo, V.; Young, J.; Gonzales, D.; Ni, Y.; Flowers, S.J.; Summer, R.; Waldman, S.A.; Kim, J.; Jung, D.Y.; Noh, H.L.; et al. Myeloid-specific deletion of Zfp36 protects against insulin resistance and fatty liver in diet-induced obese mice. Am. J. Physiol. Metab. 2018, 315, E676–E693. [Google Scholar] [CrossRef]
- Clement, S.L.; Scheckel, C.; Stoecklin, G.; Lykke-Andersen, J. Phosphorylation of Tristetraprolin by MK2 Impairs AU-Rich Element mRNA Decay by Preventing Deadenylase Recruitment. Mol. Cell. Biol. 2010, 31, 256–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vohl, M.-C.; Sladek, R.; Robitaille, J.; Gurd, S.; Marceau, P.; Richard, D.; Hudson, T.J.; Tchernof, A. A Survey of Genes Differentially Expressed in Subcutaneous and Visceral Adipose Tissue in Men*. Obes. Res. 2004, 12, 1217–1222. [Google Scholar] [CrossRef]
- Bouchard, L.; Vohl, M.-C.; Deshaies, Y.; Rhéaume, C.; Daris, M.; Tchernof, A. Visceral adipose tissue zinc finger protein 36 mRNA levels are correlated with insulin, insulin resistance index, and adiponectinemia in women. Eur. J. Endocrinol. 2007, 157, 451–457. [Google Scholar] [CrossRef]
- Lai, W.S.; Stumpo, D.J.; Blackshear, P.J. Rapid insulin-stimulated accumulation of an mRNA encoding a proline-rich protein. J. Biol. Chem. 1990, 265, 16556–16563. [Google Scholar] [CrossRef]
- Cao, H.; Urban, J.F., Jr.; Anderson, R.A. Insulin Increases Tristetraprolin and Decreases VEGF Gene Expression in Mouse 3T3–L1 Adipocytes. Obesity 2008, 16, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Keijer, J.; Bunschoten, A.; Bouwman, F.; Renes, J.; Mariman, E. Insulin modulates the secretion of proteins from mature 3T3-L1 adipocytes: A role for transcriptional regulation of processing. Diabetology 2006, 49, 2453–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H. Expression, Purification, and Biochemical Characterization of the Antiinflammatory Tristetraprolin: A Zinc-Dependent mRNA Binding Protein Affected by Posttranslational Modifications. Biochemestry 2004, 43, 13724–13738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carballo, E.; Cao, H.; Lai, W.S.; Kennington, E.A.; Campbell, D.; Blackshear, P.J. Decreased Sensitivity of Tristetraprolin-deficient Cells to p38 Inhibitors Suggests the Involvement of Tristetraprolin in the p38 Signaling Pathway. J. Biol. Chem. 2001, 276, 42580–42587. [Google Scholar] [CrossRef] [Green Version]
- Hitti, E.; Iakovleva, T.; Brook, M.; Deppenmeier, S.; Gruber, A.D.; Radzioch, D.; Clark, A.; Blackshear, P.J.; Kotlyarov, A.; Gaestel, M. Mitogen-Activated Protein Kinase-Activated Protein Kinase 2 Regulates Tumor Necrosis Factor mRNA Stability and Translation Mainly by Altering Tristetraprolin Expression, Stability, and Binding to Adenine/Uridine-Rich Element. Mol. Cell. Biol. 2006, 26, 2399–2407. [Google Scholar] [CrossRef] [Green Version]
- Marchese, F.; Aubareda, A.; Tudor, C.; Saklatvala, J.; Clark, A.; Dean, J.L.E. MAPKAP Kinase 2 Blocks Tristetraprolin-directed mRNA Decay by Inhibiting CAF1 Deadenylase Recruitment. J. Biol. Chem. 2010, 285, 27590–27600. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Taylor, W.R.; Joseph, G.; Caracciolo, V.; Gonzales, D.M.; Sidell, N.; Seli, E.; Blackshear, P.J.; Kallen, C.B. mRNA-Binding Protein ZFP36 Is Expressed in Atherosclerotic Lesions and Reduces Inflammation in Aortic Endothelial Cells. Arter. Thromb. Vasc. Biol. 2013, 33, 1212–1220. [Google Scholar] [CrossRef] [Green Version]
- Brooks, S.A.; Connolly, J.E.; Diegel, R.J.; Fava, R.A.; Rigby, W.F.C. Analysis of the function, expression, and subcellular distribution of human tristetraprolin. Arthritis Rheum. 2002, 46, 1362–1370. [Google Scholar] [CrossRef]
- Suswam, E.A.; Shacka, J.J.; Walker, K.; Lu, L.; Li, X.; Si, Y.; Zhang, X.; Zheng, L.; Nabors, L.; Cao, H.; et al. Mutant tristetraprolin: A potent inhibitor of malignant glioma cell growth. J. Neuro-Oncol. 2013, 113, 195–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deepa, S.S.; Dong, L.Q. APPL1: Role in adiponectin signaling and beyond. Am. J. Physiol. Metab. 2009, 296, E22–E36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, C.; Mertz, V.A.; Heigenhauser, G.J.F.; Dyck, D.J. The Stimulatory Effect of Globular Adiponectin on Insulin-Stimulated Glucose Uptake and Fatty Acid Oxidation Is Impaired in Skeletal Muscle from Obese Subjects. Diabetes 2005, 54, 3154–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pun, N.T.; Park, P.-H. Adiponectin inhibits inflammatory cytokines production by Beclin-1 phosphorylation and B-cell lymphoma 2 mRNA destabilization: Role for autophagy induction. Br. J. Pharmacol. 2018, 175, 1066–1084. [Google Scholar] [CrossRef] [Green Version]
- Pun, N.T.; Khakurel, A.; Shrestha, A.; Kim, S.H.; Park, P.-H. Critical role of tristetraprolin and AU-rich element RNA-binding protein 1 in the suppression of cancer cell growth by globular adiponectin. FEBS Open Bio 2018, 8, 1964–1976. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameters | All | Control | MetS | p-Value | Adjusted for Age |
|---|---|---|---|---|---|
| N | 200 (94/106) | 100 (50/50) | 100 (44/56) | - | - |
| Age (years) | 38.6 ± 8.3 | 35.4 ± 7.9 | 41.9 ± 7.2 | - | - |
| TTP (pg/mL) | 242.9 (124.7–330.9) | 150.9 (68.2–280.9) | 288.1 (229.3–386.8) | <0.001 | <0.001 |
| Leptin (pg/mL) | 145.3 (44.6–609) | 111.4 (56.1–399) | 159.7 (33.9–945.6) | 0.59 | 0.78 |
| TNF-α (pg/mL) | 1.1 (0.5–1.7) | 0.5 (0.2–1.1) | 1.5 (0.9–1.9) | <0.001 | <0.001 |
| IL-1β (pg/mL) | 0.96 (0.5–1.6) | 0.5 (0.4–0.9) | 1.4 (1.0–3.5) | <0.001 | <0.001 |
| CRP (ng/mL) | 3242 (961–6009) | 1961 (573.6–4387.9) | 4452 (1727.9–6191) | <0.001 | <0.001 |
| Adiponectin (ng/mL) | 10,872 (5173.9–20,176) | 12,904 (4974–22,843) | 10,045 (5439–16,574) | 0.18 | 0.66 |
| Insulin (µIU/mL) | 10.9 (5.2–23.1) | 8.2 (4.4–14.8) | 17.2 (7.1–43.8) | <0.001 | 0.001 |
| HOMA-IR | 2.8 (1.2–6.8) | 1.8 (0.9–3.9) | 5.2 (1.8–11.3) | <0.001 | - |
| HOMA-B | 35.3 (14.8–81.2) | 28.9 (14.2–53.8) | 50.8 (18.3–141.1) | 0.005 | - |
| Parameters | Males | Females | ||||
|---|---|---|---|---|---|---|
| N | Control | MetS | p-Value | Control | MetS | p-Value |
| 50 | 44 | 50 | 56 | |||
| Age (years) | 35.2 ± 7.8 | 41.7 ± 7.1 | <0.001 | 35.6 ± 8.2 | 42.3 ± 7.4 | <0.001 |
| TTP (pg/mL) | 95.3 (43.4–187.6) | 286 (221–372.6) | <0.001 | 231.2 (127.9–315.7) | 288.9 (237.8–397.1) | 0.007 |
| Leptin (pg/mL) | 149.9 (91.9–349.8) | 119.6 (25.8–648.6) | 0.75 | 93.8 (54.3–409.6) | 176.8 (42.1–1706) | 0.26 |
| TNF-α (pg/mL) | 0.21 (0.1–0.24) | 1.2 (0.8–1.6) | <0.001 | 0.6 (0.4–1.2) | 1.5 (0.9–2.0) | <0.001 |
| IL-1β (pg/mL) | 0.43 (0.21–0.54) | 1.3 (1.1–2.7) | <0.001 | 0.8 (0.4–1.4) | 1.6 (1.0–4.7) | <0.001 |
| CRP (ng/mL) | 1854.3 (702.5–4114) | 4570 (1890–6147) | 0.001 | 2194 (517.3–5119) | 4378 (1466–6222) | 0.004 |
| Adiponectin (ng/mL) | 14,892 (7313–20,878) | 10,504 (5811–17,376) | 0.07 | 9235 (2280–24,569) | 9721 (4481–14,499) | 0.91 |
| Insulin (µIU/mL) | 11.6 (6.5–18.9) | 17.2 (5.6–52.0) | 0.18 | 5.7 (3.6–10.9) | 17.2 (7.2–35.9) | <0.001 |
| HOMA-IR | 2.8 (1.6–4.2) | 6.2 (1.5–15.6) | 0.037 | 1.3 (0.9–2.8) | 4.6 (2.2–9.6) | <0.001 |
| HOMA-B | 42.2 (26.9–68.3) | 56.1 (17.8–159.9) | 0.37 | 17.6 (11.4–38.2) | 50.8 (18.3–130.5) | 0.003 |
| Overall | Male | Female | ||||
|---|---|---|---|---|---|---|
| B ± SE | p-Value | B ± SE | p-Value | B ± SE | p-Value | |
| Male | −0.13 ± 0.05 | 0.006 | ||||
| Central Obesity | 0.08 ± 0.05 | 0.14 | 0.17 ± 0.08 | 0.044 | 0.00 ± 0.07 | 0.96 |
| Hypertension | 0.02 ± 0.05 | 0.64 | 0.01 ± 0.08 | 0.89 | 0.06 ± 0.07 | 0.40 |
| High Blood Glucose | 0.10 ± 0.05 | 0.051 | 0.14 ± 0.08 | 0.11 | 0.06 ± 0.07 | 0.38 |
| Low HDL-C | 0.13 ± 0.05 | 0.011 | 0.14 ± 0.08 | 0.09 | 0.10 ± 0.07 | 0.14 |
| High Triglycerides | 0.11 ± 0.05 | 0.037 | 0.21 ± 0.09 | 0.016 | 0.04 ± 0.07 | 0.58 |
| Adjusted R-Square | 16.4 | 25.7 | 0.00 | |||
| Model p-value | <0.001 | <0.001 | 0.450 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Daghri, N.M.; Al-Shuwaie, A.Y.A.; Alghamdi, A.; Amer, O.E.; Khattak, M.N.K.; Ansari, M.G.A.; Alnaami, A.M.; Sabico, S. Tristetraprolin, Inflammation, and Metabolic Syndrome in Arab Adults: A Case Control Study. Biology 2021, 10, 550. https://doi.org/10.3390/biology10060550
Al-Daghri NM, Al-Shuwaie AYA, Alghamdi A, Amer OE, Khattak MNK, Ansari MGA, Alnaami AM, Sabico S. Tristetraprolin, Inflammation, and Metabolic Syndrome in Arab Adults: A Case Control Study. Biology. 2021; 10(6):550. https://doi.org/10.3390/biology10060550
Chicago/Turabian StyleAl-Daghri, Nasser M., Albatul Y.A. Al-Shuwaie, Amani Alghamdi, Osama E. Amer, Malak N.K. Khattak, Mohammed G.A. Ansari, Abdullah M. Alnaami, and Shaun Sabico. 2021. "Tristetraprolin, Inflammation, and Metabolic Syndrome in Arab Adults: A Case Control Study" Biology 10, no. 6: 550. https://doi.org/10.3390/biology10060550