

Antimicrobial Resistance Profiles and Co-Existence of Multiple Antimicrobial Resistance Genes in mcr-Harbouring Colistin-Resistant Enterobacteriaceae Isolates Recovered from Poultry and Poultry Meats in Malaysia
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Colistin Resistance in the Isolates
2.2. Phenotypic Detection of Antimicrobial Resistance
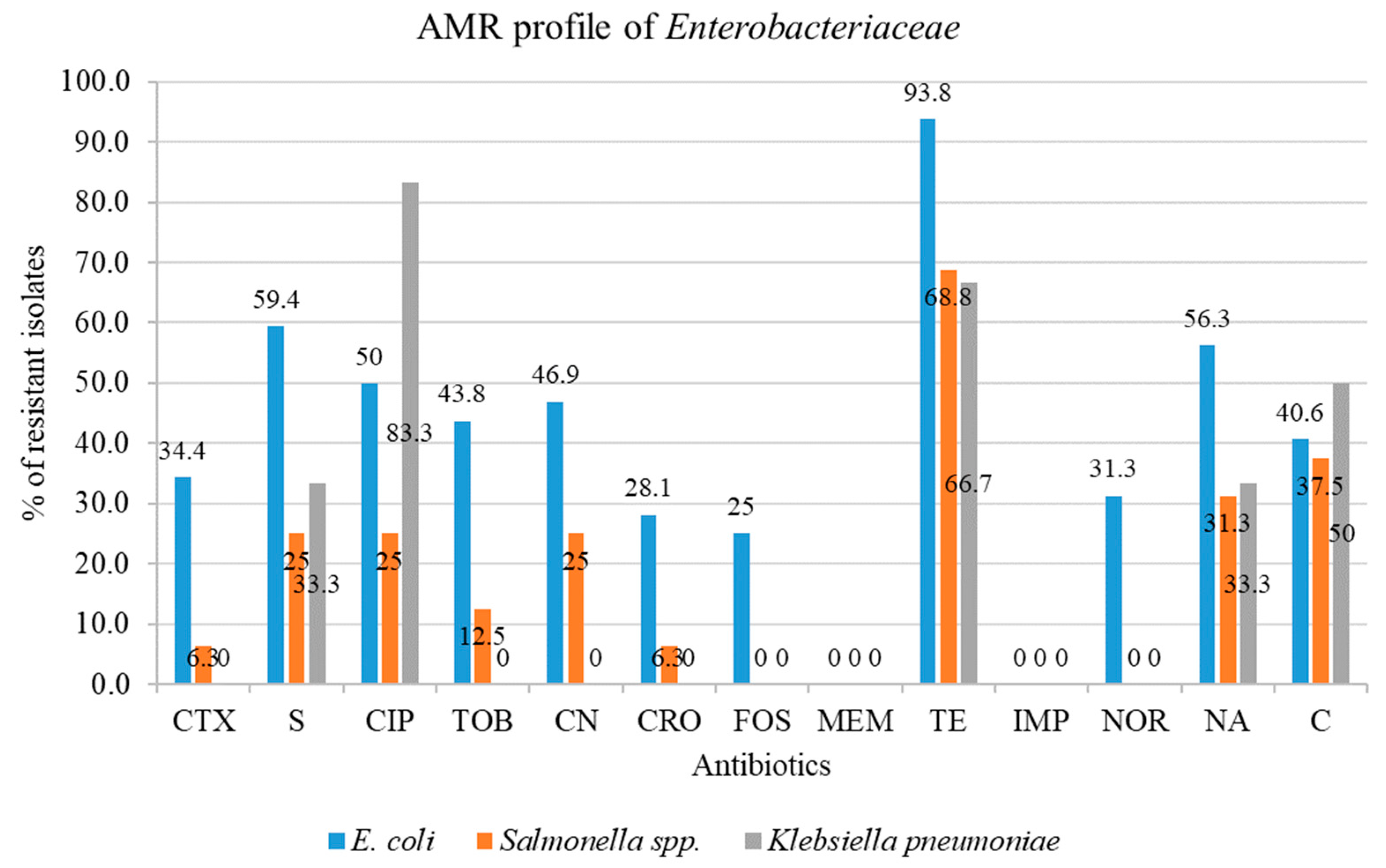
2.2.1. Rate of Antimicrobial Resistance to Enterobacteriaceae
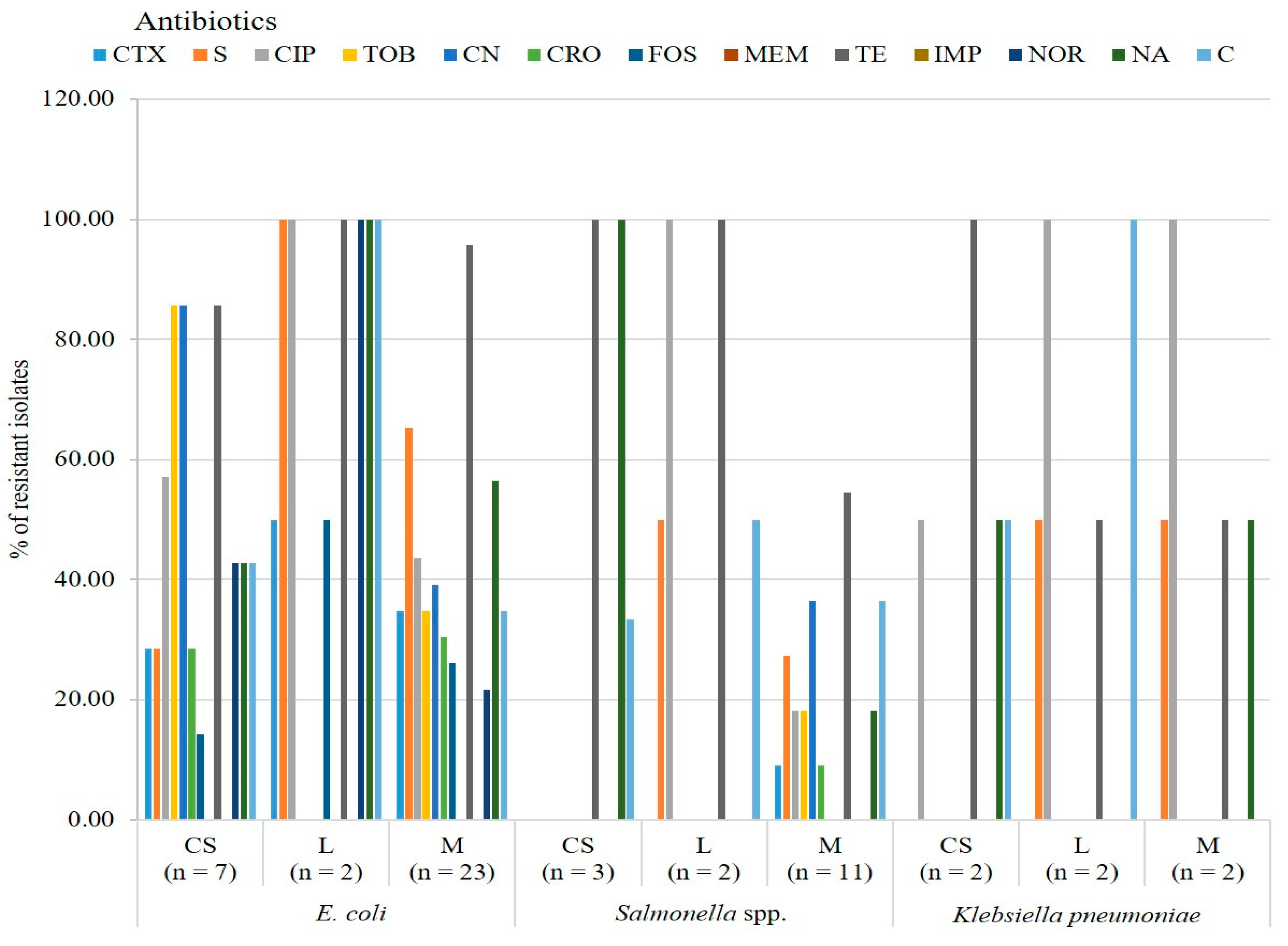
2.2.2. Antimicrobial Resistance Profile
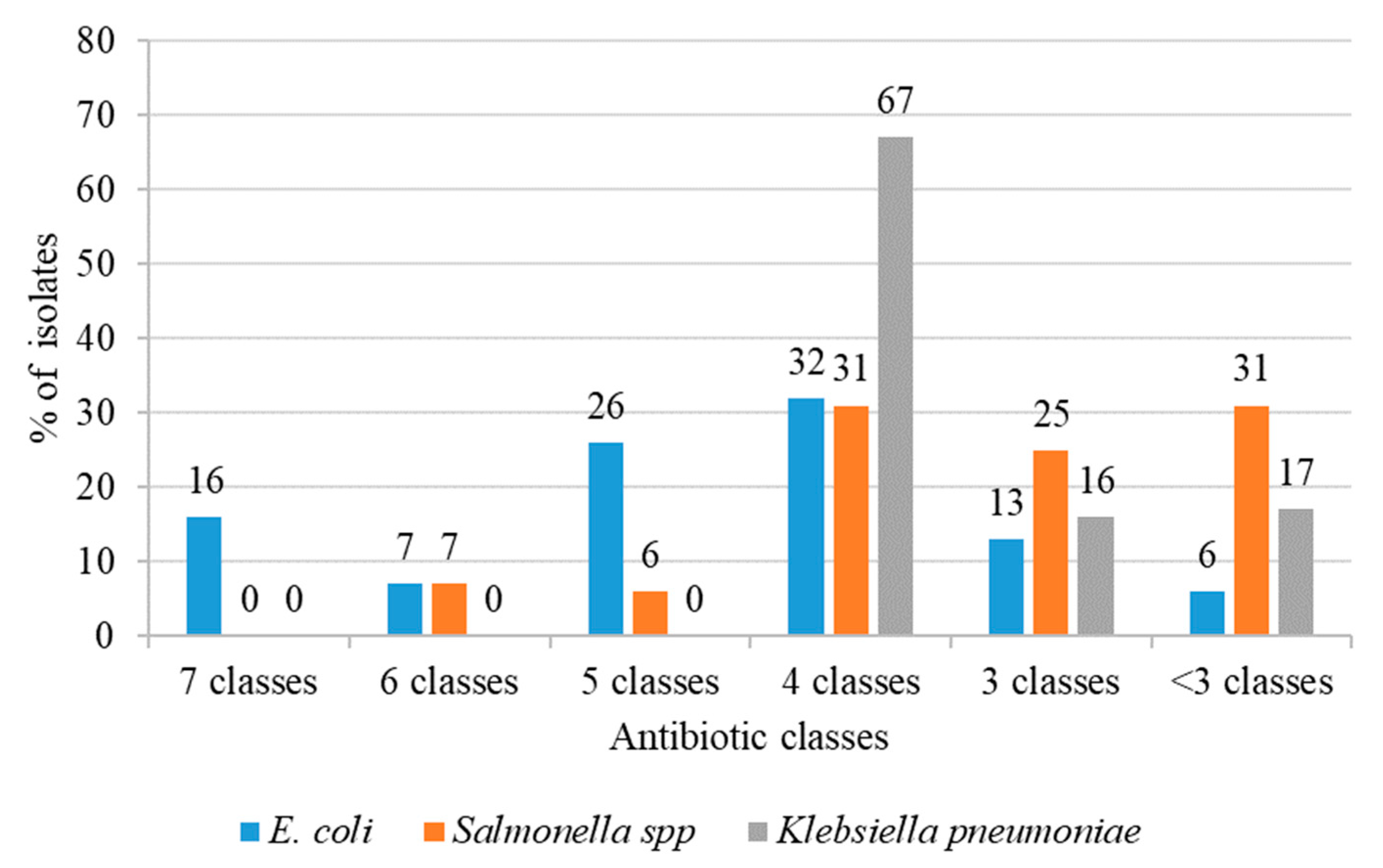
2.2.3. Multiple Antibiotic Resistance Index (MARI)
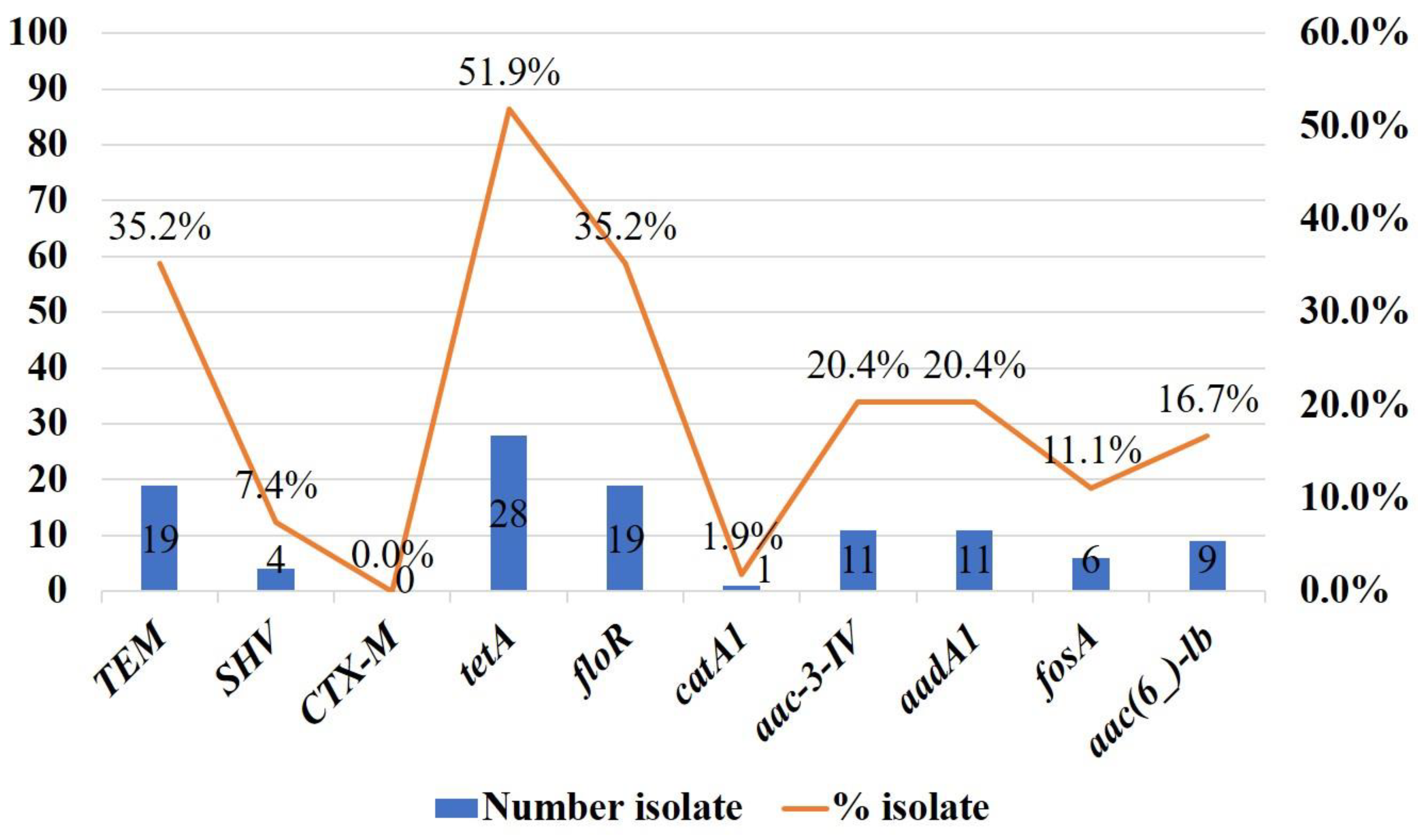
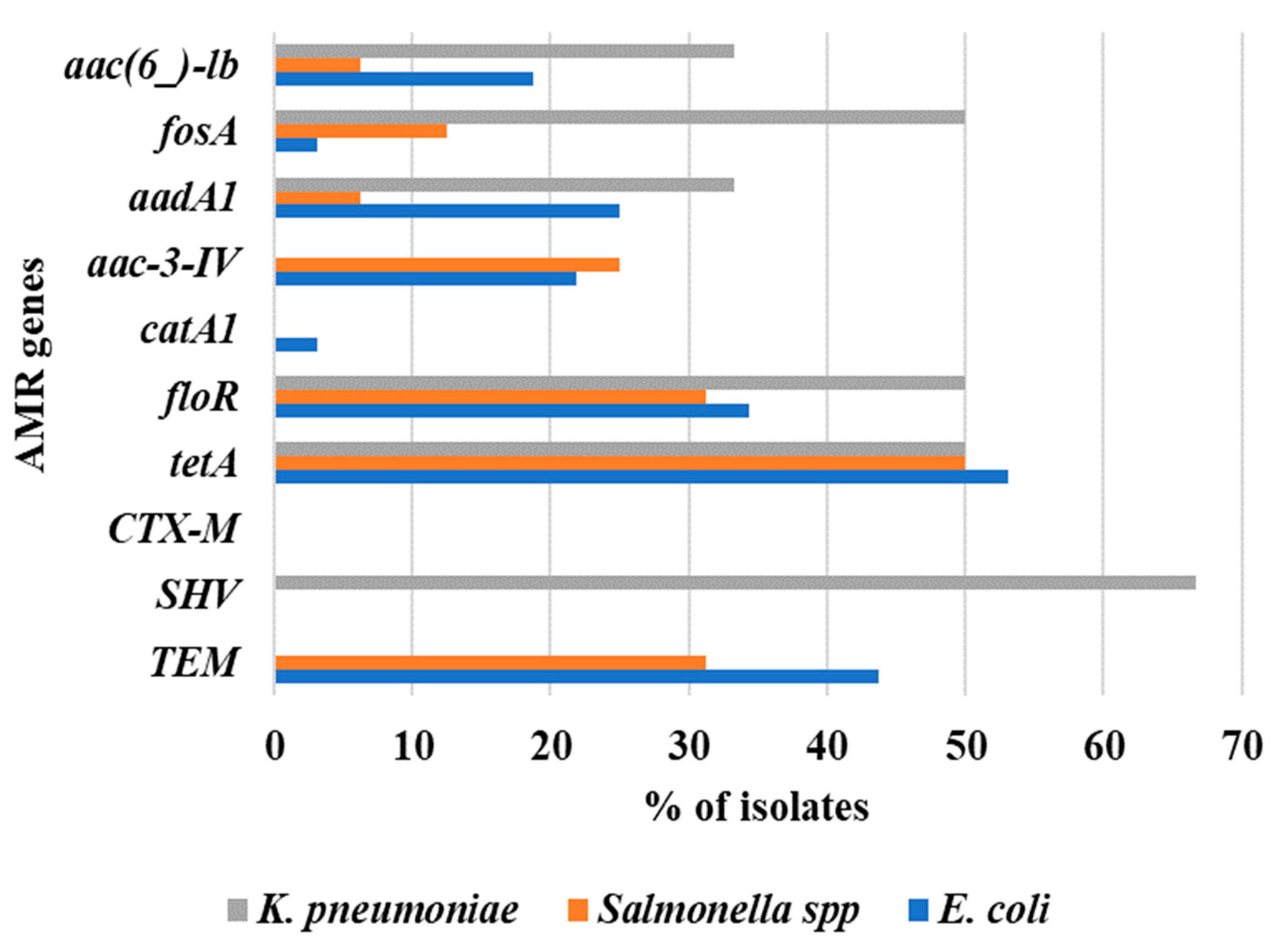
2.3. Other Antibiotic Resistance Genes in Colistin-Resistant Enterobacteriaceae
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Study Design
4.2. Phenotypic Antimicrobial Resistance Testing
4.2.1. Rates of Antimicrobial Resistance (AMR)
4.2.2. Multidrug Resistance (MDR) Patterns
4.2.3. The Multiple Antibiotic Resistance Index (MARI)
4.3. Detection of Antimicrobial Resistance Genes
4.3.1. DNA Extraction
4.3.2. Polymerase Chain Reaction
4.3.3. Agarose Gel electrophoresis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Tilahun, M.; Kassa, Y.; Gedefie, A.; Belete, M.A. Emerging Carbapenem-Resistant Enterobacteriaceae Infection, Its Epidemiology and Novel Treatment Options: A Review. Infect. Drug Resist. 2021, 14, 4363–4374. [Google Scholar] [CrossRef] [PubMed]
- Kayastha, K.; Dhungel, B.; Karki, S.; Adhikari, B.; Banjara, M.R.; Rijal, K.R.; Ghimire, P. Extended-Spectrum β-Lactamase-Producing Escherichia coli and Klebsiella Species in Pediatric Patients Visiting International Friendship Children’s Hospital, Kathmandu, Nepal. Infect. Dis. Res. Treat. 2020, 13, 1178633720909798. [Google Scholar] [CrossRef] [Green Version]
- Sharahi, J.Y.; Hashemi, A.; Ardebili, A.; Davoudabadi, S. Molecular characteristics of antibiotic-resistant Escherichia coli and Klebsiella pneumoniae strains isolated from hospitalized patients in Tehran, Iran. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 32. [Google Scholar] [CrossRef] [PubMed]
- Moubareck, C.A. Polymyxins and Bacterial Membranes: A Review of Antibacterial Activity and Mechanisms of Resistance. Membranes 2020, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Qin, W.; Lin, J.; Fang, S.; Qiu, J. Antibacterial mechanisms of polymyxin and bacterial resistance. BioMed Res. Int. 2015, 2015, 679109. [Google Scholar] [CrossRef]
- Dandachi, I.; Sokhn, E.S.; Dahdouh, E.A.; Azar, E.; El-Bazzal, B.; Rolain, J.-M.; Daoud, Z. Prevalence and Characterization of Multi-Drug-Resistant Gram-Negative Bacilli Isolated From Lebanese Poultry: A Nationwide Study. Front. Microbiol. 2018, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Muktan, B.; Thapa Shrestha, U.; Dhungel, B.; Mishra, B.C.; Shrestha, N.; Adhikari, N.; Banjara, M.R.; Adhikari, B.; Rijal, K.R.; Ghimire, P. Plasmid mediated colistin resistant mcr-1 and co-existence of OXA-48 among Escherichia coli from clinical and poultry isolates: First report from Nepal. Gut Pathog. 2020, 12, 44. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Lenhard, J.R.; Bulman, Z.P.; Tsuji, B.T.; Kaye, K.S. Shifting Gears: The Future of Polymyxin Antibiotics. Antibiotics 2019, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. MBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hou, M.; Xu, Y.; Srinivas, S.; Huang, M.; Liu, L.; Feng, Y. Action and mechanism of the colistin resistance enzyme MCR-4. Commun. Biol. 2019, 2, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [Green Version]
- Abatcha, M.G.; Effarizah, M.E.; Rusul, G. Prevalence, antimicrobial resistance, resistance genes and class 1 integrons of Salmonella serovars in leafy vegetables, chicken carcasses and related processing environments in Malaysian fresh food markets. Food Control 2018, 91, 170–180. [Google Scholar] [CrossRef]
- Aliyu, A.B.; Saleha, A.A.; Jalila, A.; Zunita, Z. Risk factors and spatial distribution of extended spectrum β-lactamase-producing- Escherichia coli at retail poultry meat markets in Malaysia: A cross-sectional study. BMC Public Health 2016, 16, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thung, T.Y.; Mahyudin, N.A.; Basri, D.F.; Radzi, C.W.J.W.M.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance of Salmonella Enteritidis and Salmonella Typhimurium in raw chicken meat at retail markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.; Hassan, L.; Sharif, Z.; Ahmad, N.; Ali, R.M.; Husin, S.A.; Hazis, N.H.b.A.; Sohaimi, N.F.M.; Bakar, S.A.; Garba, B. Analysis of Salmonella enterica serovar Enteritidis isolates from chickens and chicken meat products in Malaysia using PFGE, and MLST. BMC Vet. Res. 2020, 16, 393. [Google Scholar] [CrossRef]
- Chuah, L.O.; Shamila Syuhada, A.K.; Mohamad Suhaimi, I.; Farah Hanim, T.; Rusul, G. Genetic relatedness, antimicrobial resistance and biofilm formation of Salmonella isolated from naturally contaminated poultry and their processing environment in northern Malaysia. Food Res. Int. 2018, 105, 743–751. [Google Scholar] [CrossRef]
- Yu, C.Y.; Ang, G.Y.; Chin, P.S.; Ngeow, Y.F.; Yin, W.-F.; Chan, K.-G. Emergence of mcr-1-mediated colistin resistance in Escherichia coli in Malaysia. Int. J. Antimicrob. Agents 2016, 47, 504–505. [Google Scholar] [CrossRef]
- Aklilu, E.; Raman, K. MCR-1 Gene Encoded Colistin-Resistant Escherichia coli in Raw Chicken Meat and Bean Sprouts in Malaysia. Int. J. Microbiol. 2020, 2020, 8853582. [Google Scholar] [CrossRef]
- Mobasseri, G.; Teh, C.S.J.; Ooi, P.T.; Thong, K.L. The emergence of colistin-resistant Klebsiella pneumoniae strains from swine in Malaysia. J. Glob. Antimicrob. Resist. 2019, 17, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.R.; Thummeepak, R.; Paudel, S.; Acharya, M.; Pradhan, S.; Banjara, M.R.; Leungtongkam, U.; Sitthisak, S. Molecular Characterization of Colistin-Resistant Escherichia coli Isolated from Chickens: First Report from Nepal. Microb. Drug Resist. 2019, 25, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef] [PubMed]
- Adebowale, O.; Makanjuola, M.; Bankole, N.; Olasoju, M.; Alamu, A.; Kperegbeyi, E.; Oladejo, O.; Fasanmi, O.; Adeyemo, O.; Fasina, F.O. Multi-Drug Resistant Escherichia coli, Biosecurity and Anti-Microbial Use in Live Bird Markets, Abeokuta, Nigeria. Antibiotics 2022, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, C.; Zhang, R.; Chen, Y.; Shen, Y.; Hu, F.; Liu, D.; Lu, J.; Guo, Y.; Xia, X.; et al. Changes in colistin resistance and mcr-1 abundance in Escherichia coli of animal and human origins following the ban of colistin-positive additives in China: An epidemiological comparative study. Lancet Infect. Dis. 2020, 20, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Mead, A.; Billon-Lotz, C.; Olsen, R.; Swift, B.; Richez, P.; Stabler, R.; Pelligand, L. Epidemiological Prevalence of Phenotypical Resistances and Mobilised Colistin Resistance in Avian Commensal and Pathogenic E. coli from Denmark, France, The Netherlands, and the UK. Antibiotics 2022, 11, 631. [Google Scholar] [CrossRef]
- Johura, F.T.; Tasnim, J.; Barman, I.; Biswas, S.R.; Jubyda, F.T.; Sultana, M.; George, C.M.; Camilli, A.; Seed, K.D.; Ahmed, N.; et al. Colistin-resistant Escherichia coli carrying mcr-1 in food, water, hand rinse, and healthy human gut in Bangladesh. Gut Pathog. 2020, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Quan, J.; Li, X.; Chen, Y.; Jiang, Y.; Zhou, Z.; Zhang, H.; Sun, L.; Ruan, Z.; Feng, Y.; Akova, M.; et al. Prevalence of mcr-1 in Escherichia coli and Klebsiella pneumoniae recovered from bloodstream infections in China: A multicentre longitudinal study. Lancet Infect. Dis. 2017, 17, 400–410. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Quan, J.; Zhu, J.; Lu, J.; Wang, Y. Prevalence and molecular characteristics of mcr-1 gene in Salmonella typhimurium in a tertiary hospital of Zhejiang Province. Infect. Drug Resist. 2018, 12, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Sia, C.M.; Greig, D.R.; Day, M.; Hartman, H.; Painset, A.; Doumith, M.; Meunier, D.; Jenkins, C.; Chattaway, M.A.; Hopkins, K.L.; et al. The characterization of mobile colistin resistance (mcr) genes among 33,000 Salmonella enterica genomes from routine public health surveillance in England. Microb. Genom. 2020, 6, e000331. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-Y.; Wang, S.-F.; Lin, J.-E.; Griffith, B.T.S.; Lian, S.-H.; Hong, Z.-D.; Lin, L.; Lu, P.-L.; Tseng, S.-P. Contributions of insertion sequences conferring colistin resistance in Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2020, 55, 105894. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, S.; Crawford, M.A.; Lascols, C.; Timme, R.E.; Anderson, K.; Hodge, D.R.; Fisher, D.J.; Pillai, S.P.; Morse, S.A.; Khan, E.; et al. Resistome of carbapenem- and colistin-resistant Klebsiella pneumoniae clinical isolates. PLoS ONE 2018, 13, e0198526. [Google Scholar] [CrossRef] [Green Version]
- Gessew, G.T.; Desta, A.F.; Adamu, E. High burden of multidrug resistant bacteria detected in Little Akaki River. Comp. Immunol. Microbiol. Infect. Dis. 2022, 80, 101723. [Google Scholar] [CrossRef]
- Elshebrawy, H.A.; Abdel-Naeem, H.H.S.; Mahros, M.A.; Elsayed, H.; Imre, K.; Herman, V.; Morar, A.; Sallam, K.I. Multidrug-resistant Salmonella enterica serovars isolated from frozen chicken carcasses. LWT 2022, 164, 113647. [Google Scholar] [CrossRef]
- Adegoke, A.A.; Madu, C.E.; Aiyegoro, O.A.; Stenström, T.A.; Okoh, A.I. Antibiogram and beta-lactamase genes among cefotaxime resistant E. coli from wastewater treatment plant. Antimicrob. Resist. Infect. Control 2020, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Dandachi, I.; Chabou, S.; Daoud, Z.; Rolain, J.M. Prevalence and emergence of extended-spectrum cephalosporin-, carbapenem- and colistin-resistant gram negative bacteria of animal origin in the Mediterranean basin. Front. Microbiol. 2018, 9, 2299. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Guo, S.; Seow, K.L.G.; Ming, G.O.H.; Schlundt, J. Characterization of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolates from Jurong Lake, Singapore with Whole-Genome-Sequencing. Int. J. Environ. Res. Public Health 2021, 18, 937. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22, 30589. [Google Scholar] [CrossRef] [Green Version]
- Pragasam, A.K.; Shankar, C.; Veeraraghavan, B.; Biswas, I.; Nabarro, L.E.B.; Inbanathan, F.Y.; George, B.; Verghese, S. Molecular Mechanisms of Colistin Resistance in Klebsiella pneumoniae Causing Bacteremia from India—A First Report. Front. Microbiol. 2017, 7, 2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manges, A.R.; Smith, S.P.; Lau, B.J.; Nuval, C.J.; Eisenberg, J.N.S.; Dietrich, P.S.; Riley, L.W. Retail meat consumption and the acquisition of antimicrobial resistant Escherichia coli causing urinary tract infections: A case-control study. Foodborne Pathog. Dis. 2007, 4, 419–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falgenhauer, L.; Imirzalioglu, C.; Oppong, K.; Akenten, C.W.; Hogan, B.; Krumkamp, R.; Poppert, S.; Levermann, V.; Schwengers, O.; Sarpong, N.; et al. Detection and characterization of ESBL-producing Escherichia coli from humans and poultry in Ghana. Front. Microbiol. 2019, 10, 3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabala, R.F.; Usui, M.; Tamura, Y.; Abd-Elghany, S.M.; Sallam, K.I.; Elgazzar, M.M. Prevalence of colistin-resistant Escherichia coli harbouring mcr-1 in raw beef and ready-to-eat beef products in Egypt. Food Control 2021, 119, 107436. [Google Scholar] [CrossRef]
- Prasertsee, T.; Chokesajjawatee, N.; Santiyanont, P.; Chuammitri, P.; Deeudom, M.; Tadee, P.; Patchanee, P. Quantification and rep-PCR characterization of Salmonella spp. in retail meats and hospital patients in Northern Thailand. Zoonoses Public Health 2019, 66, 301–309. [Google Scholar] [CrossRef]
- Ghafur, A.; Shankar, C.; GnanaSoundari, P.; Venkatesan, M.; Mani, D.; Thirunarayanan, M.A.; Veeraraghavan, B. Detection of chromosomal and plasmid-mediated mechanisms of colistin resistance in Escherichia coli and Klebsiella pneumoniae from Indian food samples. J. Glob. Antimicrob. Resist. 2019, 16, 48–52. [Google Scholar] [CrossRef]
- Sharma, J.; Kumar, D.; Hussain, S.; Pathak, A.; Shukla, M.; Kumar, V.P.; Anisha, P.N.N.; Rautela, R.; Upadhyay, A.K.K.; Singh, S.P.P.; et al. Prevalence, antimicrobial resistance and virulence genes characterization of nontyphoidal Salmonella isolated from retail chicken meat shops in Northern India. Food Control 2019, 102, 104–111. [Google Scholar] [CrossRef]
- Moawad, A.A.; Hotzel, H.; Neubauer, H.; Ehricht, R.; Monecke, S.; Tomaso, H.; Hafez, H.M.; Roesler, U.; El-Adawy, H. Antimicrobial resistance in Enterobacteriaceae from healthy broilers in Egypt: Emergence of colistin-resistant and extended-spectrum β-lactamase-producing Escherichia coli. Gut Pathog. 2018, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Rahn, K.; De Grandis, S.A.A.; Clarke, R.C.C.; McEwen, S.A.A.; Galán, J.E.E.; Ginocchio, C.; Curtiss, R.; Gyles, C.L.L. Amplification of an invA gene sequence of Salmonella typhimurium by polymerase chain reaction as a specific method of detection of Salmonella. Mol. Cell. Probes 1992, 6, 271–279. [Google Scholar] [CrossRef]
- Ranjbar, R.; Izadi, M.; Hafshejani, T.T.; Khamesipour, F. Molecular detection and antimicrobial resistance of Klebsiella pneumoniae from house flies (Musca domestica) in kitchens, farms, hospitals and slaughterhouses. J. Infect. Public Health 2016, 9, 499–505. [Google Scholar] [CrossRef] [Green Version]
- CLSI. M100S: Performance Standards for Antimicrobial Susceptibility Testing an Informational Supplement for Global Application Developed through the Clinical and Laboratory Standards Institute Consensus Process, 26th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Hudzicki, J. Kirby-Bauer disk diffusion susceptibility test protocol. Am. Soc. Microbiol. 2009, 66, 208. [Google Scholar]
- Papadopoulos, D.; Papadopoulos, T.; Papageorgiou, K.; Sergelidis, D.; Adamopoulou, M.; Kritas, S.K.; Petridou, E.; Papadopoulos, D. Antimicrobial resistance rates in commensal Escherichia coli isolates from healthy pigs in Greek swine farms. J. Hell. Vet. Med. Soc. 2021, 72, 2909–2916. [Google Scholar] [CrossRef]
- Sweeney, M.T.; Lubbers, B.V.; Schwarz, S.; Watts, J.L. Applying definitions for multidrug resistance, extensive drug resistance and pandrug resistance to clinically significant livestock and companion animal bacterial pathogens. J. Antimicrob. Chemother. 2018, 73, 1460–1463. [Google Scholar] [CrossRef]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adzitey, F.; Rusul, G.; Huda, N. Prevalence and antibiotic resistance of Salmonella serovars in ducks, duck rearing and processing environments in Penang, Malaysia. Food Res. Int. 2012, 45, 947–952. [Google Scholar] [CrossRef]
- Titilawo, Y.; Sibanda, T.; Obi, L.; Okoh, A. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of faecal contamination of water. Environ. Sci. Pollut. Res. 2015, 22, 10969–10980. [Google Scholar] [CrossRef] [PubMed]
- Nzima, B.; Adegoke, A.A.; Ofon, U.A.; Al-Dahmoshi, H.O.M.; Saki, M.; Ndubuisi-Nnaji, U.U.; Inyang, C.U. Resistotyping and extended-spectrum beta-lactamase genes among Escherichia coli from wastewater treatment plants and recipient surface water for reuse in South Africa. New Microbes New Infect. 2020, 38, 100803. [Google Scholar] [CrossRef]
- Rahman, M.M.; Husna, A.; Elshabrawy, H.A.; Alam, J.; Runa, N.Y.; Badruzzaman, A.T.M.; Banu, N.A.; Al Mamun, M.; Paul, B.; Das, S.; et al. Isolation and molecular characterization of multidrug-resistant Escherichia coli from chicken meat. Sci. Rep. 2020, 10, 21999. [Google Scholar] [CrossRef]
- Shivakumaraswamy, S.K.; Vittal, V.K.D.R.; Sannejal, A.D.; Mundanda, D.M.; Chakraborty, J.R.M.R.A.; Karunasagar, I. Phenotypic & genotypic study of antimicrobial profile of bacteria isolates from environmental samples. Indian J. Med. Res. 2019, 149, 232–239. [Google Scholar]
- Ma, M.; Wang, H.; Yu, Y.; Zhang, D.; Liu, S. Detection of Antimicrobial Resistance Genes of Pathogenic Salmonella from Swine with DNA Microarray. J. Vet. Diagn. Investig. 2007, 19, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.H.; Hsueh, P.R.; Jacoby, G.A.; Hooper, D.C. Risk factors and clinical characteristics of patients with qnr-positive Klebsiella pneumoniae bacteraemia. J. Antimicrob. Chemother. 2013, 68, 2907–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Robicsek, A.; Jacoby, G.A.; Sahm, D.; Hooper, D.C. Prevalence in the United States of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrob. Agents Chemother. 2006, 50, 3953–3955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Ceballos, D.L.; Burbano-Rosero, E.M.; Mondragon, E.I. Characterization of antibiotic-resistant Escherichia coli associated with urinary tract infections in Southern Colombia. Univ. Sci. 2020, 25, 463–488. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | No. of Antibiotics (Class) | Multidrug Profile | No. of Isolates (%) | Prevalence of MDR% |
|---|---|---|---|---|
| mcr carriage Enterobacteriaceae (n = 7) | 1 (1) | Any one of the tested antibiotics | 0 | 100 |
| 2 (2) | Combination of any two antibiotics | 0 | ||
| 5 (4) | TOB, CN, TE, C, CL | 1 (14.28) | ||
| 6 (5) | TOB, CN, NA, CIP, NOR, CL | 1 (14.28) | ||
| 7 (6) | S, TE, NA, CIP, NOR, C, CL | 2 (28.57) | ||
| 10 (6) | TOB, CN, S, TE, NA, CIP, CTX, CRO, C, CL | |||
| 9 (7) | S, TE, NA, CIP, NOR, CTX, FOS, C, CL | 3 (42.86) | ||
| 11 (7) | TOB, CN, S, TE, NA, CIP, CTX, CRO, FOS, C, CL | |||
| 11 (7) | TOB, CN, S, TE, CIP, NOR, CTX, CRO, FOS, C, CL |
| Sources | Antibiotic Resistance Genes Profile (%) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| blaTEM | p-Value | blaSHV | p-Value | blaCTX-M | p-Value | tetA | p-Value | floR | p-Value | catA1 | p-Value | aac-3-IV | p-Value | aadA1 | p-Value | fosA | p-Value | aac(6′)-Ib | p-Value | |
| Cloacal swab (n = 12) | 5 (41.7) | 0.868 | 1 (8.3) | 0.03 | 0 | nc | 9 (75) | 0.025 | 4 (33.3) | 0.03 | 0 | 0.775 | 4 (33.3) | 0.45 | 4 (33.3) | 0.002 | 2 (16.7) | 0.105 | 4 (33.3) | 0.067 |
| Litter (n = 6) | 2 (33.3) | 2 (33.3) | 0 | 5 (83.3) | 5 (83.3) | 0 | 1 (16.7) | 4 (66.7) | 2 (33.3) | 2 (33.3) | ||||||||||
| Meat (n = 36) | 12 (33.3) | 1 (2.8) | 0 | 14 (38.9) | 10 (27.8) | 1 (2.8) | 11 (20.4) | 3 (8.3) | 2 (5.6) | 3 (8.3) | ||||||||||
| Total (n = 54) | 19 (35.2) | 4 (7.4) | 0 | 28 (51.9) | 19 (35.2) | 1 (1.9) | 11 (20.4) | 6 (11.1) | 9 (16.7) | 9 (16.7) | ||||||||||
| Isolates | Strains | ARG Patterns | No. of ARG | No. of Isolates (%) |
|---|---|---|---|---|
| E. coli | E297 | blaTEM, tetA, floR, aac-3-IV, aadA1, fosA, aac(6_)-lb | 7 | 1 (3.13) |
| E. coli | E48 | blaTEM, tetA, floR, aac-3-IV, aadA1, aac(6_)-lb | 6 | 1 (3.13) |
| E. coli | E172 | tetA, floR, aac-3-IV, aadA1, aac(6_)-lb | 5 | 1 (3.13) |
| E. coli | E49 | blaTEM, tetA, floR, aadA1, aac(6_)-lb | 5 | 1 (3.13) |
| E. coli | E278 | blaTEM, floR, catA1, aac-3-IV, aac(6_)-lb | 5 | 1 (3.13) |
| E. coli | E275 | blaTEM, tetA, floR, aac-3-IV | 4 | 1 (3.13) |
| E. coli |  | |||
| E. coli | blaTEM, tetA, floR, aadA1 | 4 | 4 (12.5) | |
| E. coli | ||||
| E. coli | E446 | |||
| E. coli | E331 | blaTEM, tetA, floR, aac(6_)-lb | 4 | 1 (3.13) |
| E. coli |  | blaTEM, tetA, aac-3-IV | 3 | 2 (6.25) |
| E. coli | ||||
| E. coli |  | blaTEM, tetA | 2 | 2 (6.25) |
| E. coli | ||||
| E. coli |  | tetA | 1 | 2 (6.25) |
| E. coli | ||||
| E. coli | - | - | 0 | 15 (46.87) |
| Salmonella spp. | S283 | blaTEM, tetA, floR, aac-3-IV, fosA, aac(6_)-lb | 6 | 1 (6.25) |
| Salmonella spp. | S242 | blaTEM, tetA, floR, aadA1, fosA | 5 | 1 (6.25) |
| Salmonella spp. |  | |||
| Salmonella spp. | blaTEM, tetA, floR, aac-3-IV | 4 | 3 (18.75) | |
| Salmonella spp. | ||||
| Salmonella spp. |  | |||
| Salmonella spp. | tetA | 1 | 3 (18.75) | |
| Salmonella spp. | ||||
| Salmonella spp. | - | - | 0 | 8 (50) |
| K. pneumoniae |  | blaSHV, tetA, floR, aadA1, fosA, aac(6_)-lb | 6 | 2 (33.3) |
| K. pneumoniae | ||||
| K. pneumoniae | K55 | blaSHV, tetA, floR | 3 | 1 (16.67) |
| K. pneumoniae | K402 | blaSHV, fosA | 2 | 1 (16.67) |
| K. pneumoniae | - | - | 0 | 2 (33.3) |
| Sources | Strains | Col-R Gene | Other ARG Patterns | No. of ARG | No. of Isolates (%) |
|---|---|---|---|---|---|
| Cloacal swab | E13 | mcr-1 | blaTEM, tetA, aac-3-IV | 3 | 1 (14.3) |
| E48 | mcr-1 | blaTEM, tetA, floR, aac-3-IV, aadA1, aac(6_)-lb | 6 | 1 (14.3) | |
| E297 | mcr-1 | blaTEM, tetA, floR, aac-3-IV, aadA1, fosA, aac(6_)-lb | 7 | 1 (14.3) | |
| Litter | E172 | mcr-1 | tetA, floR, aac-3-IV, aadA1, aac(6_)-lb | 5 | 1 (14.3) |
| Meat | E278 | mcr-1 | blaTEM, floR, catA1, aac-3-IV, aac(6_)-lb | 5 | 1 (14.3) |
| E331 | mcr-1 | blaTEM, tetA, floR, aac(6_)-lb | 4 | 1 (14.3) | |
| S283 | mcr-5 | blaTEM, tetA, floR, aac-3-IV, fosA, aac(6_)-lb | 6 | 1 (14.3) |
| Isolates | Sources | Number of Isolates |
|---|---|---|
| E. coli (n = 32) | Meat | 23 |
| CS | 7 | |
| Litter | 2 | |
| Salmonella spp. (n = 16) | Meat | 11 |
| CS | 3 | |
| Litter | 2 | |
| K. pneumoniae (n = 6) | Meat | 2 |
| CS | 2 | |
| Litter | 2 | |
| Total | 54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karim, M.R.; Zakaria, Z.; Hassan, L.; Mohd Faiz, N.; Ahmad, N.I. Antimicrobial Resistance Profiles and Co-Existence of Multiple Antimicrobial Resistance Genes in mcr-Harbouring Colistin-Resistant Enterobacteriaceae Isolates Recovered from Poultry and Poultry Meats in Malaysia. Antibiotics 2023, 12, 1060. https://doi.org/10.3390/antibiotics12061060
Karim MR, Zakaria Z, Hassan L, Mohd Faiz N, Ahmad NI. Antimicrobial Resistance Profiles and Co-Existence of Multiple Antimicrobial Resistance Genes in mcr-Harbouring Colistin-Resistant Enterobacteriaceae Isolates Recovered from Poultry and Poultry Meats in Malaysia. Antibiotics. 2023; 12(6):1060. https://doi.org/10.3390/antibiotics12061060
Chicago/Turabian StyleKarim, Md. Rezaul, Zunita Zakaria, Latiffah Hassan, Nik Mohd Faiz, and Nur Indah Ahmad. 2023. "Antimicrobial Resistance Profiles and Co-Existence of Multiple Antimicrobial Resistance Genes in mcr-Harbouring Colistin-Resistant Enterobacteriaceae Isolates Recovered from Poultry and Poultry Meats in Malaysia" Antibiotics 12, no. 6: 1060. https://doi.org/10.3390/antibiotics12061060