
The Research Status, Potential Hazards and Toxicological Mechanisms of Fluoroquinolone Antibiotics in the Environment

Abstract
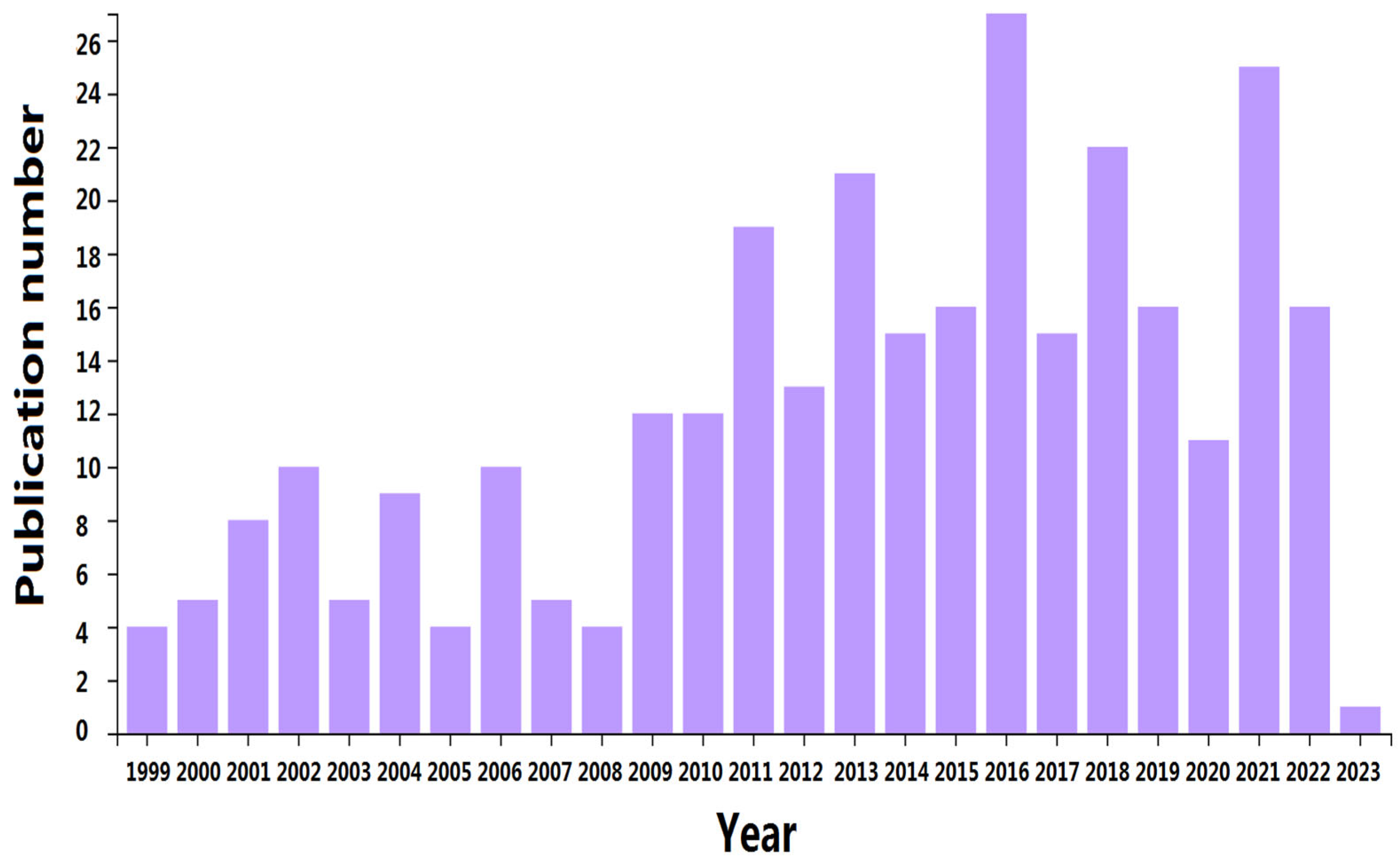
:1. Introduction
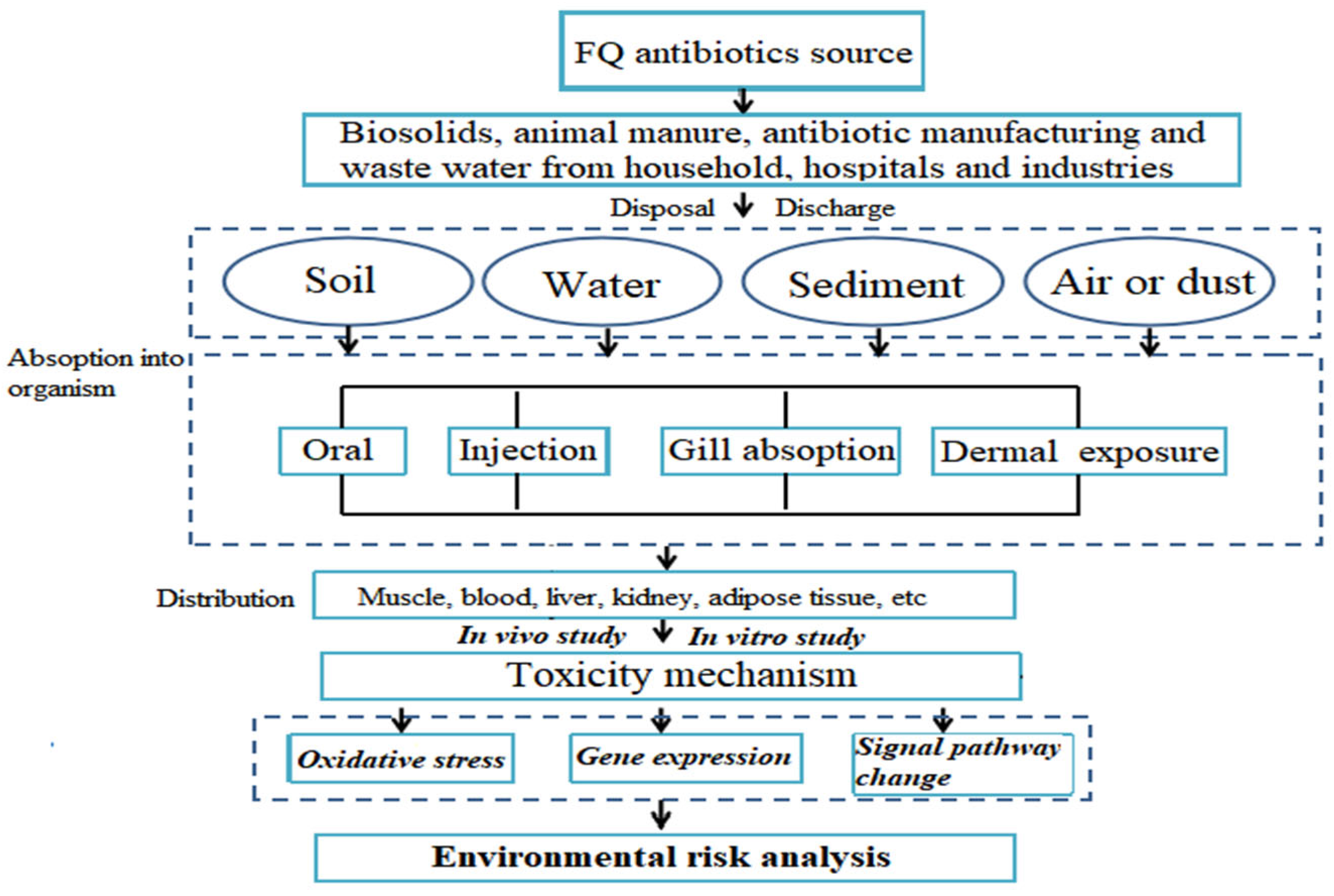
2. Source and Fate of FQ Antibiotics in the Environment
3. Toxicity Effect of FQ Antibiotics on Organisms
3.1. In Vitro Studies on FQ Antibiotics
3.2. In Vivo Studies on FQ Antibiotics
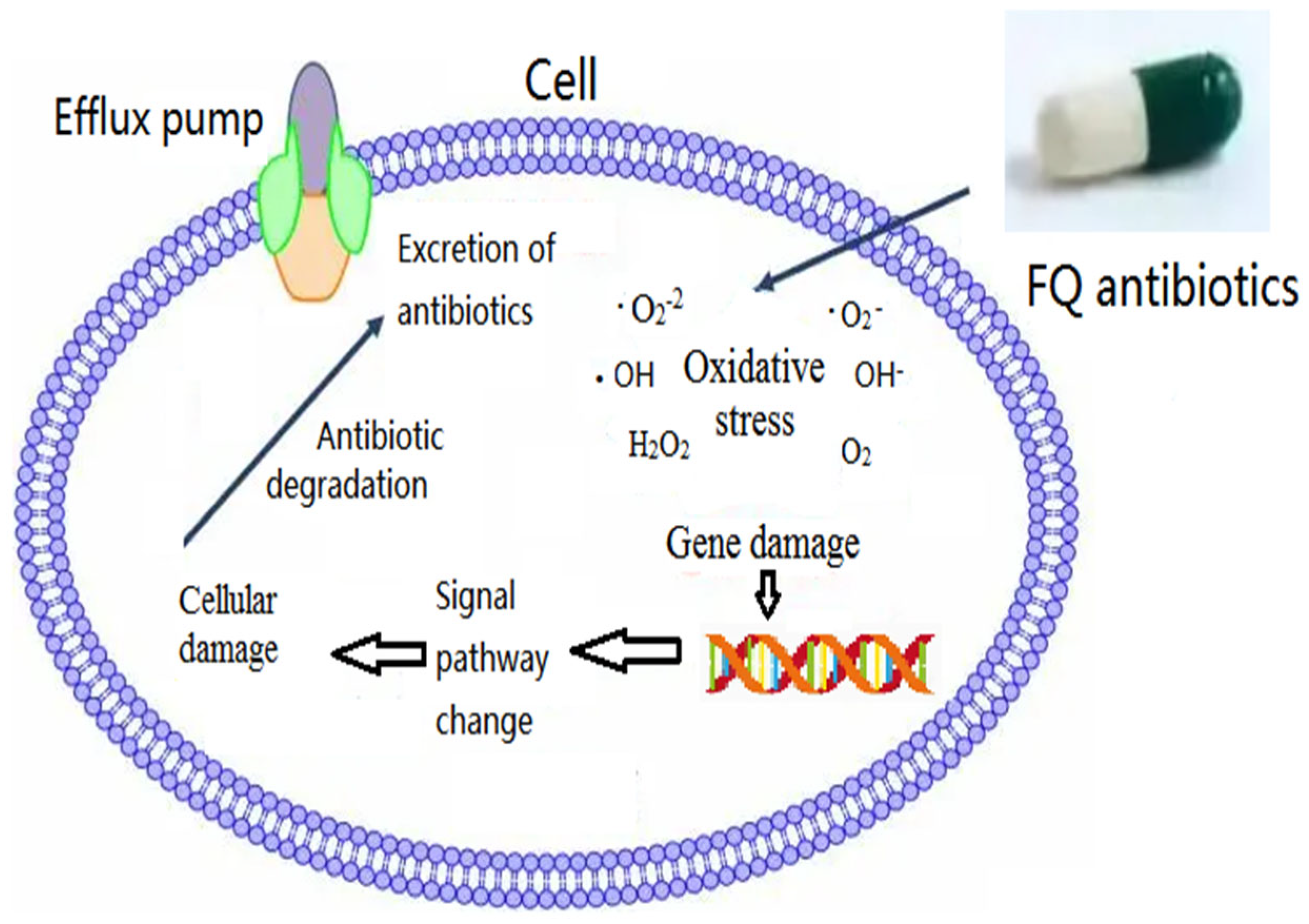
4. Environmental Toxicology Mechanisms of FQs
4.1. Oxidative Stress-Related Mechanism
4.2. Nonoxidative Stress-Related Mechanism
5. Toxicity Evaluation of Mixing Residual FQ Antibiotics and Other Pollutants

{kind=link}
{kind=link}
{kind=link}
| Pollution | Species | Toxic Effect | References |
|---|---|---|---|
| Enrofloxacin combined with two antibiotics (ciprofloxacin and florfenicol) | Human hepatocytes | Synergistic cytotoxicity | [136] |
| Enrofloxacin combined with carbendazim | Zebrafish | Reduced the hatching rate and increased the hatching malformation and lethality rates | [137] |
| Ciprofloxacin combined with ZnO nanoparticle | Firmicutes, Aminicenantes, Chloroflexi, and Parcubacteria | Additive toxic effects on methanogenesis and the degradation of proteins and carbohydrates | [138] |
| Carbon nanotube combined with two antibiotics (pentachlorophenol and ciprofloxacin) | Bacteria | Additive and synergistic toxicities | [139] |
| Polystyrene nanoplastics combined with ciprofloxacin | Corbicula fluminea | Reduced the toxic effects of ciprofloxacin on digestive glands and enhanced the inhibition rate of Corbicula fluminea siphoning | [140] |
| Enrofloxacin combined with copper | Bacteria | Comprehensive toxicity to soil enzyme activity (urease, sucrase, phosphatase, and Rubisco) | [141] |
| Norfloxacin combined with sulfamethazine | Bacteria | Reduced the methane production rate | [143] |
| Tetracycline combined with two antibiotics (ciprofloxacin and norfloxacin) | Plant | Reduced plant root elongation | [144] |
6. Conclusions
- Formulate an ecological toxicology evaluation system and guidelines to analyze the biotoxicity of FQ antibiotics at different concentration levels in multiple environments. Such a system will complete the prudent use guidelines.
- The standardized research methodology of FQ antibiotics is important for guiding enterprises in the design and production of drugs with high environmental biocompatibility.
- More information on the toxicity mechanism of FQ antibiotics is needed, which is essential for the government to control the discharge of pollutants into the environment and formulate policy coordination.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.L.; Wong, M.H. Pharmaceuticals and personal care products (PPCPs): A review on environmental contamination in China. Environ. Int. 2013, 59, 208–224. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, T.; Bilal, M.; Nabeel, F.; Adeel, M.; Iqbal, H.M.N. Environmentally related contaminants of high concern: Potential sources and analytical modalities for detection, quantifification, and treatment. Environ. Int. 2019, 122, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Sommet, A.; Benevent, J.; Rousseau, V.; Chebane, L.; Douros, A.; Montastruc, J.L.; Montastruc, F. What Fluoroquinolones Have the Highest Risk of Aortic Aneurysm? A Case/Non-case Study in VigiBase (R). J. Gen. Intern. Med. 2019, 34, 502–503. [Google Scholar] [CrossRef] [Green Version]
- Janecko, N.; Pokludova, L.; Blahova, J.; Svobodova, Z.; Literakyk, I. Implications of fluoroquinolone contamination for the aquatic environment—A review. Environ. Toxicol. Chem. 2016, 35, 2647–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, A.A.; Belden, J.B.; Lydy, M.J. Toxicity of fluoroquino lone antibiotics to aquatic organisms. Environ. Toxicol. Chem. 2005, 24, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Mascaretti, O.A. Bacteria versus Antibacterial Agents: An Integrated Approach; American Society of Microbiology: Washington, DC, USA, 2003. Available online: https://www.cabdirect.org/cabdirect/abstract/20043133239 (accessed on 27 February 2016).
- Rutgersson, C.; Fick, J.; Marathe, N.; Kristiansson, E.; Janzon, A.; Angelin, M.; Johansson, A.; Shouche, Y.; Flach, C.-F.; Larsson, D.G.J. Fluoroquinolones and qnr genes in sediment, water, soil, and human fecal flora in an environment polluted by manufacturing discharges. Environ. Sci. Technol. 2014, 48, 7825–7832. [Google Scholar] [CrossRef]
- Zhao, L.; Dong, Y.H.; Wang, H. Residues of veterinary antibiotics in manures from feedlot livestock in eight provinces of China. Sci. Total Environ. 2010, 408, 1069–1075. [Google Scholar] [CrossRef]
- Leung, H.W.; Minh, T.B.; Murphy, M.B.; Lam, J.C.W.; So, M.K.; Martin, M.; Lam, P.K.S.; Richardson, B.J. Distribution, fate and risk assessment of antibiotics in sewage treatment plants in Hong Kong, South China. Environ. Int. 2012, 42, 1–9. [Google Scholar] [CrossRef]
- Cabello, F.C.; Godfrey, H.P.; Buschmann, A.H.; Dölz, H.J. Aquaculture as yet another environmental gateway to the development and globalization of antimicrobial resistance. Lancet Infect. Dis. 2016, 16, e127–e133. [Google Scholar] [CrossRef]
- Rosal, R.; Rodríguez, A.; Perdigón-Melón, J.A.; Petre, A.; García-Calvo, E.; Gómez, M.J.; Agüera, A.; Fernández-Alba, A.R. Occurrence of emerging pollutants in urban wastewater and their removal through biological treatment followed by ozonation. Water Res. 2010, 44, 578–588. [Google Scholar] [CrossRef]
- Zhou, L.J.; Ying, G.G.; Zhao, J.L.; Yang, J.F.; Wang, L.; Yang, B.; Liu, S. Trends in the occurrence of human and veterinary antibiotics in the sediments of the Yellow River, Hai River and Liao River in northern China. Environ. Pollut. 2011, 159, 1877–1885. [Google Scholar] [CrossRef] [PubMed]
- Zarei-Baygi, A.; Harb, M.; Wang, P.; Stadler, L.B.; Smith, A.L. Evaluating antibiotic resistance gene correlations with antibiotic exposure conditions in anaerobic membrane bioreactors. Environ. Sci. Technol. 2019, 53, 3599–3609. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, G.; Patring, J.; Kreuger, J.; Norrgren, L.; Oskarsson, A. Toxicity of 15 veterinary pharmaceuticals in zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2013, 126, 30–41. [Google Scholar] [CrossRef]
- Nie, X.; Gu, J.; Lu, J.; Pan, W.; Yang, Y. Effects of norfloxacin and butylated hydroxyanisole on the freshwater microalga Scenedesmus obliquus. Ecotoxicology 2009, 18, 677–684. [Google Scholar] [CrossRef]
- Kumar, R.R.; Lee, J.T.; Cho, J.Y. Fate, occurrence, and toxicity of veterinary antibiotics in environment. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 701–709. [Google Scholar] [CrossRef]
- Gao, L.; Shi, Y.; Li, W.; Niu, H.; Liu, J.; Cai, Y. Occurrence of antibiotics in eight sewage treatment plants in Beijing, China. Chemosphere 2012, 86, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Chung, K.Y.; Lee, H.C.; Lee, J.K.; Bark, K.M. Ionization and divalent cation complexation of quinolone antibiotics in aqueous solution. Bull. Korean Chem. Soc. 2000, 21, 849–854. [Google Scholar]
- Kümmerer, K. Antibiotics in the aquatic environment—A review–part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Conkle, J.L.; Lattao, C.; White, J.R.; Cook, R.L. Competitive sorption and desorption behavior for three fluoroquinolone antibiotics in a wastewater treatment wetland soil. Chemosphere 2010, 80, 1353–1359. [Google Scholar] [CrossRef]
- Golet, E.M.; Strehler, A.; Alder, A.C.; Giger, W. Determination of fluoroquinolone antibacterial agents in sewage sludge and sludgetreated soil using accelerated solvent extraction followed by solid-phase extraction. Anal. Chem. 2002, 74, 5455–5462. [Google Scholar] [CrossRef]
- Youngquist, C.P.; Liu, J.; Orfe, L.H.; Jones, S.S.; Call, D.R. Ciprofloxacin residues in municipal biosolid compost do not selectively enrich populations of resistant bacteria. Appl. Environ. Microbiol. 2014, 80, 7521–7526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthikeyan, K.G.; Meyer, M.T. Occurrence of antibiotics in wastewater treatment facilities in Wisconsin, USA. Sci. Total Environ. 2006, 361, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Gibs, J.; Heckathorn, H.A.; Meyer, M.T.; Klapinski, F.R.; Alebus, M.; Lippincott, R.L. Occurrence and partitioning of antibiotic compounds found in the water column and bottom sediments from a stream receiving two wastewater treatment plant effluents in Northern New Jersey, 2008. Sci. Total Environ. 2013, 458–460, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Government of Canada. Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS) Annual Report; Chapter 2: Antimicrobial Resistance; Public Health Agency of Canada: Guelph, ON, Canada, 2014. [Google Scholar]
- American Association of Swine Veterinarians. News Archive: Baytril FDA Approved for Control of E. coli in Swine. 2015. Available online: https://www.aasv.org/news/story.php? id¼7798 (accessed on 7 April 2016).
- Campagnolo, E.R.; Johnson, K.R.; Karpati, A.; Rubin, C.S.; Kolpin, D.W.; Meyer, M.T.; Esteban, J.E.; Currier, R.W.; Smith, K.; Thu, K.M.; et al. Antimicrobial residues in animal waste and water resources proximal to large-scale swine and poultry feeding operations. Sci. Total Environ. 2002, 299, 89–95. [Google Scholar] [CrossRef]
- Heberer, T. Occurrence, fate, and removal of pharmaceutical residues in the aquatic environment: A review of recent research data. Toxicol. Lett. 2002, 131, 5–17. [Google Scholar] [CrossRef]
- Zhou, J.L.; Kang, Y. Matrix effect in high-performance liquid hromatography-tandem mass spectrometry analysis of antibiotics in environmental water samples. J. Sep. Sci. 2013, 36, 564–571. [Google Scholar] [CrossRef]
- Wu, Y.B.; Wang, Z.S.; Liao, X.D.; Chen, Z. Study on the excretion of enrofloxacin in chicken and its degradation in chicken feces. Acta Vet. Zootech. Sin. 2005, 36, 1069. [Google Scholar]
- Janusch, F.; Scherz, G.; Mohring, S.A.; Hamscher, G. Determination of fluoroquinolones in chicken feces–A new liquid–liquid extraction method combined with LC–MS/MS. Environ. Toxicol. Pharmacol. 2014, 38, 792–799. [Google Scholar] [CrossRef]
- Mutiyar, P.K.; Mittal, A.K. Risk assessment of antibiotic residues in different water matrices in India: Key issues and challenges. Environ. Sci. Pollut. Res. 2014, 21, 7723–7736. [Google Scholar] [CrossRef]
- Kummerer, K. The presence of pharmaceuticals in the environment due to human use—Present knowledge and future challenges. J. Environ. Manag. 2009, 90, 2354–2366. [Google Scholar] [CrossRef] [PubMed]
- Thuy, H.T.T.; Nga, L.P.; Loan, T.T.C. Antibiotic contaminants in coastal wetlands from Vietnamese shrimp farming. Environ. Sci. Pollut. Res. 2011, 18, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Emara, Y.; Lehmann, A.; Siegert, M.W.; Finkbeiner, M. Modeling pharmaceutical emissions and their toxicity-related effects in life cycle assessment (LCA): A review. Integr. Environ. Assess. Manag. 2019, 15, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Khan, A.; Wahid, A.; Butt, Z.; Farhan, M.; Ahmad, F. Role of hospital eflfluents in the contribution of antibiotics and antibiotic resistant bacteria to the aquatic environment. Pak. J. Nutr. 2012, 11, 1177–1182. [Google Scholar] [CrossRef] [Green Version]
- Verlicchi, P.; Aukidy, M.A.; Galletti, A.; Petrovic, M.; Barcel, D. Hospital efflfluent: Investigation of the concentrations and distribution of pharmaceuticals and environmental risk assessment. Sci. Total Environ. 2012, 430, 109–118. [Google Scholar] [CrossRef]
- Hartmann, A.; Alder, A.C.; Koller, T.; Widmer, R.M. Identifification of flfluoroquinolone antibiotics as the main source of umuC genotoxicity in native hospital wastewater. Environ. Toxicol. Chem. 1998, 17, 377–382. [Google Scholar] [CrossRef]
- Kookana, R.S.; Williams, M.; Boxall, A.B.A.; Larsson, D.G.J.; Gaw, S.; Choi, K.; Yamamoto, H.; Thatikonda, S.; Zhu, Y.G.; Carriquiriborde, P. Potential ecological footprints of active pharmaceutical ingredients: An examination of risk factors in low-, middle- and high-income countries. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369, 20130586. [Google Scholar] [CrossRef]
- Fick, J.; Soderstrom, H.; Lindberg, R.H.; Phan, C.; Tysklind, M.; Larsson, D.G.J. Contamination of surface, ground, and drinking water from pharmaceutical production. Environ. Toxicol. Chem. 2009, 28, 2522–2527. [Google Scholar] [CrossRef]
- Gao, P.; Mao, D.; Luo, Y.; Wang, L.; Xu, B.; Xu, L. Occurrence of sulfonamide and tetracycline-resistant bacteria and resistance genes in aquaculture environment. Water Res. 2012, 46, 2355–2364. [Google Scholar] [CrossRef]
- Lindberg, R.H.; Olofsson, U.; Rendahl, P.; Johansson, M.I.; Tysklind, M.; Andersson, B.A. Behavior of fluoroquinolones and trimethoprim during mechanical, chemical, and active sludge treatment of sewage water and digestion of sludge. Environ. Sci. Technol. 2006, 40, 1042–1048. [Google Scholar] [CrossRef]
- Jjemba, P.K. Excretion and ecotoxicity of pharmaceutical and personal care products in the environment. Ecotoxicol. Environ. Saf. 2006, 63, 113–130. [Google Scholar] [CrossRef]
- Zhang, T.; Li, B. Occurrence, transformation, and fate of antibiotics in municipal wastewater treatment plants. Crit Rev Environ. Sci. Technol. 2011, 41, 951–998. [Google Scholar] [CrossRef]
- Grenni, P.; Ancona, V.; Barra Caracciolo, A. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 136, 25–39. [Google Scholar] [CrossRef]
- Zarfl, C.; Klasmeier, J.; Matthies, M. A conceptual model describing the fate of sulfadiazine and its metabolites observed in manure-amended soils. Chemosphere 2009, 77, 720–726. [Google Scholar] [CrossRef]
- Förster, M.; Laabs, V.; Lamshöft, M.; Groeneweg, J.; Zühlke, S.; Spiteller, M.; Krauss, M.; Kaupenjohann, M.; Amelung, W. Sequestration of manure-applied sulfadiazine residues in soils. Environ. Sci. Technol. 2009, 43, 1824–1830. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Wagil, M.; Kumirska, J.; Stolte, S.; Puckowski, A.; Maszkowska, J.; Stepnowski, P.; Białk-Bielinska, A. Development of sensitive and reliable LC-MS/MS methods for the determination of three fluoroquinolones in water and fifish tissue samples and preliminary environmental risk assessment of their presence in two rivers in northern Poland. Sci. Total Environ. 2014, 493, 1006–1013. [Google Scholar] [CrossRef]
- Rico, A.; Dimitrov, M.R.; Van Wijngaarden, R.P.A.; Satapornvanit, K.; Smidt, H.; Van den Brink, P.J. Effects of the antibiotic enrofloxacin on the ecology of tropical eutrophic freshwater microcosms. Aquat. Toxicol. 2014, 147, 92–104. [Google Scholar] [CrossRef]
- Karnjanapiboonwong, A.; Suski, J.G.; Shah, A.A.; Cai, Q.; Morse, A.N.; Anderson, T.A. Occurrence of PPCPs at a wastewater treatment plant and in soil and groundwater at a land application site. Water Air Soil Pollut. 2011, 216, 257–273. [Google Scholar] [CrossRef]
- Rico, A.; Oliveira, R.; McDonough, S.; Matser, A.; Khatikarn, J.; Satapornvanit, K.; Nogueira, A.J.A.; Soares, A.M.V.M.; Domingues, I.; Van den Brink, P.J. Use, fate and ecological risks of antibiotics applied in tilapia cage farming in Thailand. Environ. Pollut. Barking Essex. 2014, 191, 8–16. [Google Scholar] [CrossRef]
- Chee-Sanford, J.C.; Mackie, R.I.; Koike, S.; Krapac, I.G.; Lin, Y.-F.; Yannarell, A.C.; Maxwell, S.; Aminov, R.I. Fate and transport of antibiotic residues and antibiotic resistance genes following land application of manure waste. J. Environ. Qual. 2009, 38, 1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.H.; Jho, E.H.; Jeong, S.; Choi, S.; Kal, Y.; Cha, S. Photodegradation of tetracycline and sulfathiazole individually and in mixtures. Food Chem. Toxicol. 2018, 116, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Tsurumaki, Y.; Takei, M.; Hosaka, M.; Oomori, Y. In vitro method for prediction of the phototoxic potentials of fluoroquinolones. Toxicol. Vitr. 2001, 15, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, M.; Berger, J.; Möller, J.-G.; Döring, G. Moxifloxacin and Ciprofloxacin Protect Human Respiratory Epithelial Cells against Streptococcus pneumoniae, Staphylococcus aureus, Pseudomonas aeruginosa, and Haemophilus influenzae in Vitro. Infection 2005, 33, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, T.; Goto, K.; Watanabe, K.; Hashimoto, K.; Suzuki, T.; Kishino, H.; Fujimoto, K.; Mori, K. Fluoroquinolones suppress gluconeogenesis by inhibiting fructose 1,6-bisphosphatase in primary monkey hepatocytes. Toxicol. Vitr. 2020, 65, 104786. [Google Scholar] [CrossRef] [PubMed]
- Kempka, G.; Ahr, H.J.; Ruther, W.; Schluter, G. Effects of fluoroquinolones and glucocorticoids on cultivated tendon cells in vitro. Toxicol. Vitr. 1996, 10, 743–754. [Google Scholar] [CrossRef]
- Bezwada, P.; Clark, L.A.; Schneider, S. Intrinsic cytotoxic effects of fluoroquinolones on human corneal keratocytes and endothelial cells. Curr. Med. Res. Opin. 2008, 24, 419–427. [Google Scholar] [CrossRef]
- Matsumoto, S.; Way, W.; Tarlo, K.; Short, B. Comparative toxicity of fluoroquinolone antibiotics on corneal cells in vitro. Cornea 2006, 25, S1–S7. [Google Scholar] [CrossRef]
- Bai, Z.L.; Chen, Q.; Yang, S.D.; Zhang, F.; Wang, H.Y.; Yang, D.L.; Ding, W.Y. Toxic Effects of Levofloxacin on Rat Annulus Fibrosus Cells: An In-vitro Study. Med. Sci. Monit. 2015, 20, 2205–2212. [Google Scholar]
- Kaye, S.; McLean, K.; Foulkes, D.M.; Sloniecka, M.; Byrne, D.; Haneef, A.S.; Winstanley, C.; Berry, N.G.; Fernig, D.G. P-18 Impact of fluoroquinolones and aminoglycosides on P. aeruginosa virulence factor production and cytotoxicity. BMJ Open Ophthalmol. 2022, 7, A6. [Google Scholar]
- Marquez-Lazaro, J.P.; Mora, L.; Mendez-Cuadro, D.; Rodriguez-Cavallo, E.; Toldra, F. In vitro oxidation promoted by sarafloxacin antibiotic residues on myosin and chicken meat proteins. J. Food Compos. Anal. 2022, 111, 104622. [Google Scholar] [CrossRef]
- Silva, F.B.E.; Carrijo-Carvalho, L.C.; Teixeira, A.; de Freitas, D.; Carvalho, F.R.D. Toxicity of Intracameral Injection of Fourth-Generation Fluoroquinolones on the Corneal Endothelium. Cornea 2016, 35, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Salimiaghdam, N.; Singh, L.; Schneider, K.; Nalbandian, A.; Chwa, M.; Atilano, S.R.; Bao, A.; Kenney, M.C. Potential adverse effects of ciprofloxacin and tetracycline on ARPE-19 cell lines. BMJ Open Ophthalmol. 2021, 5, e000458. [Google Scholar] [CrossRef]
- Oum, B.S.; Kim, N.M.; Lee, J.S.; Park, Y.M. Effects of Fluoroquinolone Eye Solutions without Preservatives on Human Corneal Epithelial Cells in vitro. Ophthalmic Res. 2014, 51, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Segreti, J.; Jones, R.N.; Bertino, J.S., Jr. Challenges in Assessing Microbial Susceptibility and Predicting Clinical Response to Newer-Generation Fluoroquinolones. J. Ocul. Pharmacol. Ther. 2012, 28, 1. [Google Scholar] [CrossRef] [PubMed]
- Maurin, M.; Bryskier, A.; Raoult, D. Antibiotic Susceptibilities of Parachlamydia acanthamoeba in Amoebae. Antimicrob. Agents Chemother. 2002, 46, 3065–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, M.C.S.; Junior, I.N.; De Souz, M.V.N. In vitro activity of ciprofloxacin, ofloxacin, levofloxacin, sparfloxacin and gatifloxacin against multidrug-resistant Mycobacterium tuberculosis in Rio de Janeiro Brazil. Médecine Et Mal. Infect. 2007, 37, 295–296. [Google Scholar] [CrossRef]
- Frean, J.; Klugman, K.P.; Arntzen, L.; Bukofzer, S. Susceptibility of Bacillus anthracis to eleven antimicrobial agents including novel fluoroquinolones and a ketolide. J. Antimicrob. Chemother. 2003, 52, 297–299. [Google Scholar] [CrossRef] [Green Version]
- Andoh, M.; Naganawa, T.; Yamaguchi, T.; Fukushi, H.; Hirai, K. In vitro susceptibility to tetracycline and fluoroquinolones of Japanese isolates of Coxiella burnetii. Microbiol. Immunol. 2004, 48, 661–664. [Google Scholar] [CrossRef]
- Murray, P.R.; Bratcher, J.L.; Niles, A.C.; Hampton, C.M. In vitro activity of nine fluoroquinolone antibiotics against 200 strains of enterococci. Diagn. Microbiol. Infect. Dis. 1993, 16, 83–85. [Google Scholar] [CrossRef]
- Trujillano-Martin, I.; Garcia-Sanchez, E.; Martinez, I.M.; Fresnadillo, M.J.; Garcia-Sanchez, J.E.; Garcia-Rodriguez, J.A. In vitro activities of six new fluoroquinolones against Brucella melitensis. Antimicrob. Agents Chemother. 1999, 43, 194–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, S.H.; Ian, M.; Dean, E. A comparison of the bactericidal activity of quinolone antibiotics in a Mycobacterium fortuitum model. J. Med. Microbiol. 2001, 50, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koss, M.J.; Eder, M.; Blumenkranz, M.S.; Klauss, V.; Ta, C.N.; de Kaspar, H.M. The effectiveness of the new fluoroquinolones against the normal bacterial flora of the conjunctiva. Ophthalmologe 2007, 104, 21–27. [Google Scholar] [CrossRef]
- Mammeri, H.; Poirel, L.; Nordmann, P. Bactericidal activity of fluoroquinolones against plasmid-mediated QnrA-producing Escherichia coli. Clin. Microbiol. Infect. 2005, 11, 1048–1049. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, M.; Maeda, S.; Deguchi, T. In vitro activity of fluoroquinolones against Mycoplasma genitalium and their bacteriological efficacy for treatment of M-genitalium-positive nongonococcal urethritis in men. Clin. Infect. Dis. 2005, 41, 1357–1359. [Google Scholar] [CrossRef] [Green Version]
- Muller, E.; Hom, S. Efficacy of enrofloxacin and marbofloxacin on bacteria isolated from dogs and cats—In vitro data of antibiotic resistance. Prakt. Tierarzt 2007, 90, 512. [Google Scholar]
- Steward, J.; Lever, M.S.; Russell, P.; Beedham, R.J.; Stagg, A.J.; Taylor, R.R.; Brooks, T.J.G. Efficacy of the latest fluoroquinolones against experimental Yersinia pestis. Int. J. Antimicrob. Agents 2004, 24, 609–612. [Google Scholar] [CrossRef]
- Rizk, M.A.; AbouLaila, M.; El-Sayed, S.A.S.; Guswanto, A.; Yokoyama, N.; Igarashi, I. Inhibitory effects of fluoroquinolone antibiotics on Babesia divergens and Babesia microti, blood parasites of veterinary and zoonotic importance. Infect. Drug Resist. 2018, 11, 1605–1615. [Google Scholar] [CrossRef] [Green Version]
- Maitre, T.; Petitjean, G.; Chauffour, A.; Bernard, C.; El Helali, N.; Jarlier, V.; Reibel, F.; Chavanet, P.; Aubry, A.; Veziris, N. Are moxifloxacin and levofloxacin equally effective to treat XDR tuberculosis? J. Antimicrob. Chemother. 2017, 72, 2326–2333. [Google Scholar] [CrossRef] [Green Version]
- Shadoud, L.; Almahmoud, I.; Jarraud, S.; Etienne, J.; Larrat, S.; Schwebel, C.; Timsit, J.F.; Schneider, D.; Maurin, M. Hidden Selection of Bacterial Resistance to Fluoroquinolones In Vivo: The Case of Legionella pneumophila and Humans. Ebiomedicine 2015, 2, 1179–1185. [Google Scholar] [CrossRef] [Green Version]
- Quinn, T.; Bolla, J.-M.; Pagès, J.-M.; Fanning, S. Antibiotic-resistant Campylobacter: Could efflux pump inhibitors control infection? J. Antimicrob. Chemother. 2007, 59, 1230–1236. [Google Scholar] [CrossRef] [PubMed]
- Regmi, N.L.; Abd El-Aty, A.M.; Kubota, R.; Shah, S.S.; Shimoda, M. Lack of inhibitory effects of several fluoroquinolones on cytochrome P-450 3A activities at clinical dosage in dogs. J. Vet. Pharmacol. Ther. 2007, 30, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Li, J.P.; Fan, Q.Y.; Zuo, J.; Xue, B.Q.; Zhang, X.L.; Wei, Y.; Sun, L.Y.; Grenier, D.; Yi, L.; Hou, X.G. Paeoniflorin combined with norfloxacin ameliorates drug-resistant Streptococcus suis infection. J. Antimicrob. Chemother. 2022, 77, 3275–3282. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Ahmad, S.; Ali, N.; Rahman, S.U.; Hussain, H.; Alghamdi, S.; Almehmadi, M.; Dablool, A.S.; Bannunah, A.M.; Bukhari, S.H. Insulinotropic Potential of Moxifloxacin and Gemifloxacin: An In Vivo Rabbits Model Study Followed by Randomized Phase I Clinical Trial. Antibiotics 2022, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Hao, G.; Qu, Y.C.; Chen, L.; Hua, W.Y.; Zong, S.L.; Wang, M.; Su, C.J.; Zhang, Q.Y.; Du, Z.Y. Comparative effect of ciprofloxacin and moxifloxacin on the modulation of bile acid profiles and gut microbiota in rats. Braz. J. Pharm. Sci. 2022, 58, e191086. [Google Scholar] [CrossRef]
- Ellerbrock, R.E.; Canisso, I.F.; Larsen, R.J.; Garrett, K.S.; Stewart, M.C.; Herzog, K.K.; Kersh, M.E.; Moshage, S.G.; Podico, G.; Lima, F.S. Fluoroquinolone exposure in utero did not affect articular cartilage of resulting foals. Equine Vet. J. 2020, 53, 385–396. [Google Scholar] [CrossRef]
- Tang, Y.J.; Xu, J.Q.; Chen, L.; Qiu, J.L.; Liu, Y.; Ouyang, G.F. Rapid in vivo determination of fluoroquinolones in cultured puffer fish (Takifugu obscurus) muscle by solid-phase microextraction coupled with liquid chromatography-tandem mass spectrometry. Talanta 2017, 175, 550–556. [Google Scholar] [CrossRef]
- Lin, J.N.; Lai, C.H.; Huang, Y.H.; Yang, C.H. Antimicrobial Effects of Minocycline, Tigecycline, Ciprofloxacin, and Levofloxacin against Elizabethkingia anophelis Using In Vitro Time-Kill Assays and In Vivo Zebrafish Animal Models. Antibiotics 2021, 10, 285. [Google Scholar] [CrossRef]
- Du, J.; Liu, Q.H.; Fu, L. Metabolic and transcriptional disruption of American shad (Alosa sapidissima) by enrofloxacin in commercial aquaculture. Environ. Sci. Pollut. Res. 2021, 29, 2052–2062. [Google Scholar] [CrossRef]
- Xiao, C.Q.; Han, Y.; Liu, Y.; Zhang, J.; Hu, C.Q. Relationship Between Fluoroquinolone Structure and Neurotoxicity Revealed by Zebrafish Neurobehavior. Chem. Res. Toxicol. 2018, 31, 238–250. [Google Scholar] [CrossRef]
- Stroman, D.W.; Dajcs, J.J.; Cupp, G.A.; Schlech, B.A. In vitro and in vivo potency of moxifloxacin and moxifloxacin ophthalmic solution 0.5%, a new topical fluoroquinolone. Surv. Ophthalmol. 2005, 50, S16–S31. [Google Scholar] [CrossRef] [PubMed]
- Riaz, L.; Mahmood, T.; Coyne, M.S.; Khalid, A.; Rashid, A.; Hayat, M.T.; Gulzar, A.; Amjad, M. Physiological and antioxidant response of wheat (Triticum aestivum) seedlings to fluoroquinolone antibiotics. Chemosphere 2017, 17, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.K.; Yang, Y.T.; Zhao, C.X.; Huang, X.R.; Chen, H.M.; Zhao, W.L.; Yang, X.R.; Zhu, Y.G.; Liu, H.J. ROS as a key player in quinolone antibiotic stress on Arabidopsis thaliana: From the perspective of photosystem function, oxidative stress and phyllosphere microbiome. Sci. Total Environ. 2022, 848, 157821. [Google Scholar] [CrossRef]
- Gomes, M.P.; Brito, J.C.M.; Carvalho, M.M.L.; da Cunha, C.M.R.R.; Garcia, Q.S.; Figueredo, C.C. Responses of the nitrogen-fixing aquatic fern Azolla to water contaminated with ciprofloxacin: Impacts on biofertilization. Environ. Pollut. 2018, 232, 293–299. [Google Scholar] [CrossRef]
- Hillis, D.G.; Fletcher, J.; Solomon, K.R.; Sibley, P.K. Effects of Ten Antibiotics on Seed Germination and Root Elongation in Three Plant Species. Arch. Environ. Contam. Toxicol. 2011, 60, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Rydzyński, D.; Piotrowicz-Cieślak, A.I.; Grajek, H.; Michalczyk, D.J. Instability of chlorophyll in yellow lupin seedlings grown in soil contaminated with ciprofloxacin and tetracycline. Chemosphere 2017, 184, 62–73. [Google Scholar] [CrossRef]
- Hu, Y.; Habibul, N.; Meng, F.-L.; Sheng, G.-P. Chemical speciation of ciprofloxacin in aqueous solution regulates its phytotoxicity and uptake by rice (Oryza sativa L.). Sci. Total Environ. 2021, 771, 144787. [Google Scholar] [CrossRef] [PubMed]
- Opriş, O.; Copaciu, F.; Soran, M.L.; Ristoiu, D.; Niinemets, Ü.; Copolovici, L. Influence of nine antibiotics on key secondary metabolites and physiological characteristics in Triticum aestivum: Leaf volatiles as a promising new tool to assess toxicity. Ecotoxicol. 2013, 87, 70–79. [Google Scholar] [CrossRef]
- Sayre, L.M.; Moreira, P.I.; Smith, M.A.; Perry, G. Metal ions and oxidative protein modification in neurological disease. Ann. Ist. Super Sanita 2005, 41, 143–164. [Google Scholar]
- Cheng, G.Y.; Hao, H.H.; Dai, M.H.; Liu, Z.L.; Yuan, Z.H. Antibacterial action of quinolones: From target to network. Eur. J. Med. Chem. 2013, 66, 555–562. [Google Scholar] [CrossRef]
- Chetelat, A.A.; Albertini, S.; Gocke, E. The photomutagenicity of fluoroquinolones in tests for gene mutation, chromosomal aberration, gene conversion and DNA breakage (Comet assay). Mutagenesis 1996, 996, 497–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, R.S.; Agrawal, N.; Misra, R.B.; Farooq, M.; Hans, R.K. Radiation-induced in vitro phototoxic potential of some fluoroquinolones. Drug Chem. Toxicol. 2006, 29, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Spratt, T.E.; Schultz, S.S.; Levy, D.E.; Chen, D.; Schluter, G.; Williams, G.M. Different mechanisms for the photoinduced production of oxidative DNA damage by fluoroquinolones differing in photostability. Chem. Res. Toxicol. 1999, 12, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Gurbay, A.; Gonthier, B.; Signorini-Allibe, N.; Barret, L.; Favier, A.; Hincal, F. Ciprofloxacin-induced DNA damage in primary culture of rat astrocytes and protection by Vitamin E. NeuroToxicology 2006, 27, 6–10. [Google Scholar] [CrossRef]
- Pan, X.R.; Qin, P.F.; Liu, R.T.; Li, J.F.; Zhang, F.C. Molecular mechanism on two fluoroquinolones inducing oxidative stress: Evidence from copper/zinc superoxide dismutase. RSC Adv. 2016, 6, 91141–91149. [Google Scholar] [CrossRef]
- Beberok, A.; Buszman, E.; Zdybel, M.; Pilawa, B.; Wrześniok, D. EPR examination of free radical properties of DOPA-melanin complexes with ciprofloxacin, lomefloxacin, norfloxacin and sparfloxacin. Chem. Phys. Lett. 2010, 497, 115–122. [Google Scholar] [CrossRef]
- Trisciuoglio, D.; Krasnowska, E.; Maggi, A.; Pozzi, R.; Parasassi, T.; Sapora, O. Phototoxic effect of fluoroquinolones on two human cell lines. Toxicol. Vitr. 2002, 16, 449–456. [Google Scholar] [CrossRef]
- Huang, J.H.; Liao, L.Z.; Wang, G.; Du, Z.; Wu, Z.X. Reproductive toxicity of enrofloxacin in Caenorhabditis elegans involves oxidative stress-induced cell apoptosis. J. Environ. Sci. 2023, 127, 726–737. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Wang, L.F.; Zhuang, H.; Li, X.X.; Gao, X.J.; An, Z.H.; Liu, X.D.; Yang, H.; Wei, W.Z.; Zhang, X.J. Excessive use of enrofloxacin leads to growth inhibition of juvenile giant freshwater prawn Macrobrachium rosenbergii. Ecotoxicol. Environ. Saf. 2019, 169, 344–352. [Google Scholar] [CrossRef]
- Liang, X.M.; Wang, F.; Li, K.; Nie, X.; Fang, H.S. Effects of norfloxacin nicotinate on the early life stage of zebrafish (Danio rerio): Developmental toxicity, oxidative stress and immunotoxicity. Fish Shellfish Immunol. 2020, 96, 262–269. [Google Scholar] [CrossRef]
- Luo, T.L.; Chen, J.W.; Li, X.; Zhang, S.; Yao, H.; Peijnenburg, W.J.G.M. Effects of lomefloxacin on survival, growth and reproduction of Daphnia magna under simulated sunlight radiation. Ecotoxicol. Environ. Saf. 2018, 166, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.; Veiga, V.; Frankenbach, S.; Serôdio, J.; Pinto, G. Evaluation of physiological changes induced by the fluoroquinolone antibiotic ciprofloxacin in the freshwater macrophyte species Lemna minor and Lemna gibba. Environ. Toxicol. Pharmacol. 2019, 72, 103242. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Liu, Y.Y.; Liu, C.; Zou, H.Y.; Wang, Y.; Li, W.T.; Bi, D.R.; He, Q.G. Laboratory detection of haemophilus parasuis with decreased susceptibility to nalidixic acid and enrofloxacin due to GyrA and ParC mutations. J. Anim. Vet. Adv. 2011, 10, 2870–2873. [Google Scholar]
- Bai, H.; Du, J.F.; Hu, M.; Qi, J.; Cai, Y.N.; Niu, W.W.; Liu, Y.Q. Analysis of mechanisms of resistance and tolerance of Escherichia coli to enrofloxacin. Ann. Microbiol. 2012, 62, 293–298. [Google Scholar] [CrossRef]
- Sheng, Z.G.; Cao, X.J.; Peng, S.; Wang, C.; Li, Q.; Wang, Y.; Liu, M.F. Ofloxacin induces apoptosis in microencapsulated juvenile rabbit chondrocytes by caspase-8-dependent mitochondrial pathway. Toxicol. Appl. Pharmacol. 2008, 226, 119–127. [Google Scholar] [CrossRef]
- Soto, S.M.; Ruiz, J.; Mendoza, M.C.; Vila, J. In vitro fluoroquinolone-resistant mutants of Salmonella enterica serotype Enteritidis: Analysis of mechanisms involved in resistance. Int. J. Antimicrob. Agents 2003, 22, 537–540. [Google Scholar] [CrossRef]
- Fief, C.A.; Hoang, K.G.; Phipps, S.D.; Wallace, J.L.; Deweese, J.E. Examining the Impact of Antimicrobial Fluoroquinolones on Human DNA Topoisomerase II alpha and II beta. Acs Omega 2019, 4, 4049–4055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramon, M.S.; Canton, E.; Peman, J.; Pastor, A.; Martinez, J.P. Mechanisms of action of quinolones against staphylococci and relationship with their in vitro bactericidal activity. Chemotherapy 1999, 45, 175–182. [Google Scholar] [CrossRef]
- Shen, R.; Yu, Y.; Lan, R.; Yu, R.; Yuan, Z.; Xia, Z. The cardiovascular toxicity induced by high doses of gatifloxacin and ciprofloxacin in zebrafish. Environ. Pollut. 2019, 254, 112861. [Google Scholar] [CrossRef]
- Niu, Z.G.; Xu, W.A.; Na, J.; Lv, Z.; Zhang, Y. How long-term exposure of environmentally relevant antibiotics may stimulate the growth of Prorocentrum lima: A probable positive factor for red tides. Environ. Pollut. 2019, 255, 1. [Google Scholar] [CrossRef]
- Hong, J.; Huang, X.H.; Wang, Z.; Luo, X.; Huang, S.; Zheng, Z. Combined toxic effects of enrofloxacin and microplastics on submerged plants and epiphytic biofilms in high nitrogen and phosphorus waters. Chemosphere 2022, 308, 136099. [Google Scholar] [CrossRef] [PubMed]
- Kocak, E.; Ozkul, C. Comparative Proteomic Analysis of Escherichia coli Under Ofloxacin Stress. Turk. J. Pharm. Sci. 2021, 18, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Volff, J.; Vandewiele, D.; Decaris, B. Stimulation of genetic instability and associated large genomic rearrangements in streptomyces-ambofaciens by 3 fluoroquinolones. Antimicrob. Agents Chemother. 1994, 38, 1984–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahiya, S.; Kapil, A.; Lodha, R.; Kumar, R.; Das, B.K.; Sood, S.; Kabra, S.K. Induction of resistant mutants of Salmonella enterica serotype Typhi under ciprofloxacin selective pressure. Indian J. Med. Res. 2014, 139, 746–753. [Google Scholar]
- Mandell, L.A.; Ball, P.; Tillotson, G. Antimicrobial safety and tolerability: Differences and dilemmas. Clin. Infect. Dis. 2001, 32, S72–S79. [Google Scholar] [CrossRef]
- Jiang, T.Y.; Kustermann, S.; Wu, X.Q.; Zihlmann, C.; Zhang, M.F.; Mao, Y.; Wu, W.W. Mitochondrial dysfunction is underlying fluoroquinolone toxicity: An integrated mitochondrial toxicity assessment. Mol. Cell. Toxicol. 2022, 19, 333–342. [Google Scholar] [CrossRef]
- Dalhoff, A. Selective toxicity of antibacterial agents-still a valid concept or do we miss chances and ignore risks? Infection 2021, 49, 29–56. [Google Scholar] [CrossRef]
- Sendzik, J.; Shakibaei, M.; Schafer-Korting, M.; Stahlmann, R. Fluoroquinolones cause changes in extracellular matrix, signalling proteins, metalloproteinases and caspase-3 in cultured human tendon cells. Toxicology 2005, 212, 24–36. [Google Scholar] [CrossRef]
- Metterlein, T.; Schuster, F.; Tadda, L.; Hager, M.; Muldoon, S.; Capacchione, J.; Roewer, N.; Anetseder, M. Fluoroquinolones influence the intracellular calcium handling in individuals susceptible to malignant hyperthermia. Muscle Nerve 2011, 44, 208–212. [Google Scholar] [CrossRef]
- Zhan, L.; Zhang, J.Q.; Zhao, B.; Li, X.; Zhang, X.; Hu, R.; Elken, E.M.; Kong, L.; Gao, Y.H. Genomic and Transcriptomic Analysis of Bovine Pasteurella multocida Serogroup A Strain Reveals Insights into Virulence Attenuation. Front. Vet. Sci. 2021, 8, 765495. [Google Scholar] [CrossRef]
- Guo, X.W.; Li, W.Y.; An, R.; Huang, M.; Yu, Z.G. Composite ammonium glycyrrhizin has hepatoprotective effects in chicken hepatocytes with lipopolysaccharide/enrofloxacin-induced injury. Exp. Ther. Med. 2020, 20, 5. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Lu, Q.R.; Yang, Y.; Martínez, M.-A.; Lopez-Torres, B.; Martínez-Larrañaga, M.-R.; Wang, X.; Anadón, A.; Ares, I. A proposed “steric-like effect” for the slowdown of enrofloxacin antibiotic metabolism by ciprofloxacin, and its mechanism. Chemosphere 2021, 284, 131347. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.H.; Chen, K.X.; Zhao, J.J.; Cheng, L.L. Comparative Study on Synergistic Toxicity of Enrofloxacin Combined with Three Antibiotics on Proliferation of THLE-2 Cell. Antibiotics 2022, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.Q.; Zhang, W.J.; Jia, L.; Luo, S.L.; Liu, Y.; Jin, Y.P.; Li, Y.C.; Yuan, X.Y.; Chen, Y.Q. Antagonistic Effects of Enrofloxacin on Carbendazim-Induced Developmental Toxicity in Zebrafish Embryos. Toxics 2022, 9, 349. [Google Scholar] [CrossRef]
- Zhao, L.; Ji, Y.; Sun, P.Z.; Li, R.Y.; Xiang, F.; Wang, H.Y.; Ruiz-Martinez, J.; Yang, Y.K. Effects of individual and complex ciprofloxacin, fullerene C-60, and ZnO nanoparticles on sludge digestion: Methane production, metabolism, and microbial community. Bioresour. Technol. 2018, 267, 46–53. [Google Scholar] [CrossRef]
- Deng, R.; Yang, K.; Lin, D.H. Pentachlorophenol and ciprofloxacin present dissimilar joint toxicities with carbon nanotubes to Bacillus subtilis. Environ. Pollut. 2021, 270, 116071. [Google Scholar] [CrossRef]
- Guo, X.Y.; Cai, Y.P.; Ma, C.; Han, L.; Yang, Z.F. Combined toxicity of micro/nano scale polystyrene plastics and ciprofloxacin to Corbicula fluminea in freshwater sediments. Sci. Total Environ. 2021, 789, 147887. [Google Scholar] [CrossRef]
- Yang, R.; Wang, J.H.; Zhu, L.S.; Wang, J.; Yang, L.L.; Mao, S.S.; Conkle, J.L.; Chen, Y.Y.; Kim, Y.M. Effects of interaction between enrofloxacin and copper on soil enzyme activity and evaluation of comprehensive toxicity. Chemosphere 2022, 268, 129208. [Google Scholar] [CrossRef]
- Ren, Z.X.; Xu, H.H.; Wang, Y.W.; Li, Y.F.; Han, S.; Ren, J.B. Combined toxicity characteristics and regulation of residual quinolone antibiotics in water environment. Chemosssphere 2021, 263, 128301. [Google Scholar] [CrossRef]
- Zhao, L.; Ji, Y.; Sun, P.Z.; Deng, J.H.; Wang, H.Y.; Yang, Y.K. Effects of individual and combined zinc oxide nanoparticle, norfloxacin, and sulfamethazine contamination on sludge anaerobic digestion. Bioresour. Technol. 2019, 273, 454–461. [Google Scholar] [CrossRef]
- Yang, L.; Feng, Y.X.; Zhang, H.; Yu, X.Z. Estimating the synergistic and antagonistic effects of dual antibiotics on plants through root elongation test. Ecotoxicology 2020, 30, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Bombaywala, S.; Mandpe, A.; Paliya, S.; Kumar, S. Antibiotic resistance in the environment: A critical insight on its occurrence, fate, and eco-toxicity. Environ. Sci. Pollut. Res. 2021, 28, 24889–24916. [Google Scholar] [CrossRef] [PubMed]



| Species | Types | Effect | References |
|---|---|---|---|
| HeLa cells | Norfloxacin, ciprofloxacin, and enoxacin | Phototoxicity, cytotoxicity, and DNA strand-breaking activity | [56] |
| Primary monkey hepatocytes | Moxifloxacin and gatifloxacin | Inhibiting fructose 1,6-bisphosphatase | [58] |
| Tendon cells (human, dog, mini pig, rat, marmoset) | Ciprofloxacin | Cytotoxicity and achilles tendon rupture | [59] |
| Corneal epithelial cell | Moxifloxacin and ciprofloxacin | Upregulation of type III secretion system and cytotoxic effects | [63] |
| Human corneal keratocytes and endothelial cells | Ciprofloxacin, gatifloxacin, levofloxacin, moxifloxacin, and ofloxacin | Cytotoxicity | [60] |
| Human corneal epithelial cells | Ofloxacin, levofloxacin, tosufloxacin, moxifloxacin, and gatifloxacin | Severe cellular morphological damage | [67] |
| Myosin and chicken meat proteins | Ciprofloxacin and sarafloxacin | Effect on meat proteins’ nutritional and technological properties | [61] |
| Streptococcus pneumoniae, Staphylococcus aureus, Staphylococcus epidermidis, and Haemophilus influenza | Besifloxacin, gatifloxacin, and moxifloxacin | Bactericidal activity | [68] |
| Intracellular bacteria of amoebae | Ofloxacin and ciprofloxacin | Antibacterial activity | [69] |
| Mycobacterium tuberculosis | Ciprofloxacin, ofloxacin, levofloxacin, sparfloxacin, and gatifloxacin | Antibacterial activity | [70] |
| Human retinal pigment epithelial cells | Ciprofloxacin and tetracycline | Upregulation of genes related to apoptosis, inflammation, and antioxidant pathways | [66] |
| Species | Types | Effect | References |
|---|---|---|---|
| American shad | Enrofloxacin | Affects many signaling pathways, such as the glycolysis/gluconeogenesis and pentose phosphate pathways | [92] |
| Human | Moxifloxacin | Inhibition of bacterial enzymes needed for bacterial DNA synthesis | [94] |
| Mouse | Enrofloxacin, enoxacin, trovafloxacin, norfloxacin, and ofloxacin | Antibabesial effect | [81] |
| Zebrafish | Levofloxacin | Decreased mortality | [91] |
| Chicken | FQ antibiotics | Decreasing the morbidity and mortality associated with the treatment of antibiotic-resistant Campylobacter | [84] |
| Puffer fish | FQ antibiotics | Residues in dorsal-epaxial muscle of living puffer fish | [90] |
| Dog | Enrofloxacin, ofloxacin, orbifloxacin, and ciprofloxacin | Lack of inhibitory effects of several FQs on cytochrome P-450 3A activities | [85] |
| Rabbits | Moxifloxacin and gemifloxacin | Influence blood glucose levels and serum insulin levels | [87] |
| Rat | Ciprofloxacin and moxifloxacin | Reduced the hydrophobicity index of the bile acid pool, reduced secondary bile acids, and increased taurine-conjugated primary bile acids | [88] |
| Foal | Enrofloxacin | Severe cartilage erosions, synovitis, and joint capsular thickening | [89] |
| Pig | Enrofloxacin | Induced catalase (CAT) and glutathione peroxidase (Gpx) and increased CYP450 content in pig liver microsomes | [86] |
| Wheat | Ciprofloxacin, enrofloxacin, and levofloxacin | Damaged the physiological structure, reduced crop productivity, and decreased growth | [95] |
| Arabidopsis thaliana | Enrofloxacin, norfloxacin, and levofloxacin | Induced oxidative stress and increased reactive oxygen species (ROS) levels and malondialdehyde (MDA) content | [96] |
| Azolla | Ciprofloxacin | Decreased photosynthesis and nitrogenase activity and altered plants’ amino acid profile, with decreases in cell N concentrations | [97] |
| Lettuce, alfalfa, and carrot | Levofloxacin | Phytotoxic | [98] |
| Lupin | Ciprofloxacin | Decreased leaf chlorophyll content | [99] |
| Rice | Ciprofloxacin | Inhibited plant growth, decreased photosynthetic pigment contents, and enhanced antioxidant enzyme activities | [100] |
| Toxic Effect | Mechanisms | References |
|---|---|---|
| In vitro phototoxicity | Formation of reactive oxygen species | [105] |
| DNA damage | Both type I (radical) and type II (singlet oxygen) mechanisms | [106] |
| DNA damage | Oxidative stress | [107] |
| Conformational and functional changes in Cu/Zn-SOD molecule | Cellular oxidative stress | [108] |
| ROS production | Lipid peroxidation chain reaction | [110] |
| Reproductive toxicity and cell apoptosis | Oxidative stress | [111] |
| Increase in the activity of the antioxidant enzyme catalase and decrease in lipid peroxidation levels | Oxidative stress | [115] |
| Bacterial resistance | GyrA mutations | [116] |
| Escherichia coli sensitivity to enrofloxacin | Target gene mutations and efflux pump activity | [117] |
| Apoptosis | Caspase-8-dependent mitochondrial pathway | [118] |
| Increase in minimal inhibitory concentrations to some FQs | AcrAB-like efflux pumps | [119] |
| Damage in the enzymes that ligate cleaved DNA and in the structure of DNA | Catalytic mechanism of topoisomerase IV and DNA gyrase | [120] |
| Bactericidal activity | Cell division as well as bacterial protein and RNA synthesis | [121] |
| Cardiovascular toxicity | Downregulation of genes associated with calcium signaling pathway and cardiac muscle contraction | [122] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, J.; Liu, Q.; Pan, Y.; Xu, S.; Li, H.; Tang, J. The Research Status, Potential Hazards and Toxicological Mechanisms of Fluoroquinolone Antibiotics in the Environment. Antibiotics 2023, 12, 1058. https://doi.org/10.3390/antibiotics12061058
Du J, Liu Q, Pan Y, Xu S, Li H, Tang J. The Research Status, Potential Hazards and Toxicological Mechanisms of Fluoroquinolone Antibiotics in the Environment. Antibiotics. 2023; 12(6):1058. https://doi.org/10.3390/antibiotics12061058
Chicago/Turabian StyleDu, Jia, Qinghua Liu, Ying Pan, Shaodan Xu, Huanxuan Li, and Junhong Tang. 2023. "The Research Status, Potential Hazards and Toxicological Mechanisms of Fluoroquinolone Antibiotics in the Environment" Antibiotics 12, no. 6: 1058. https://doi.org/10.3390/antibiotics12061058