
Synergism between the Synthetic Antibacterial and Antibiofilm Peptide (SAAP)-148 and Halicin

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Bactericidal Activities of SAAP-148 and Halicin
2.2. Anti-Biofilm Activities of SAAP-148 and Halicin
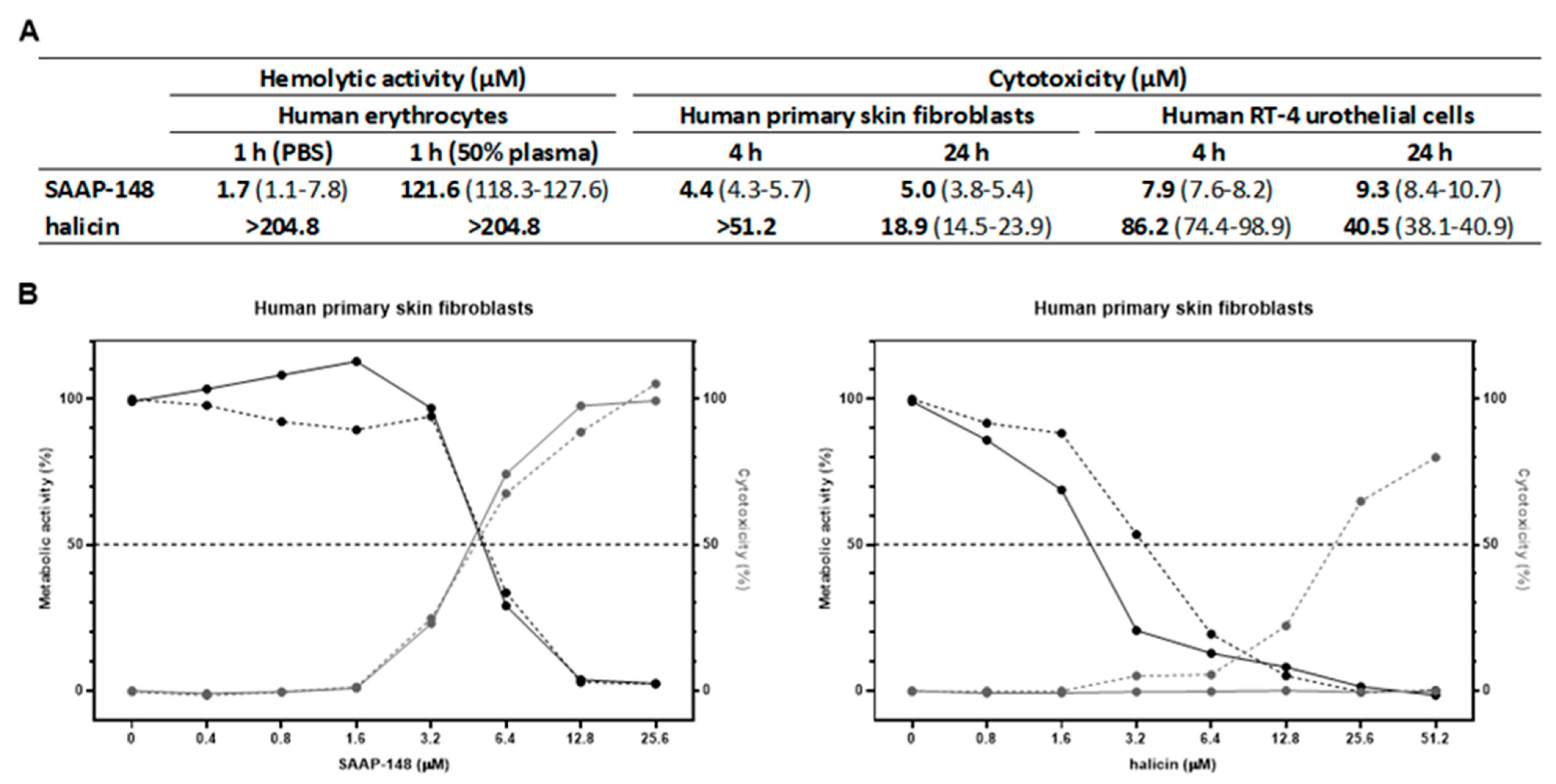
2.3. Hemolytic and Cytotoxic Activities of SAAP-148 and Halicin
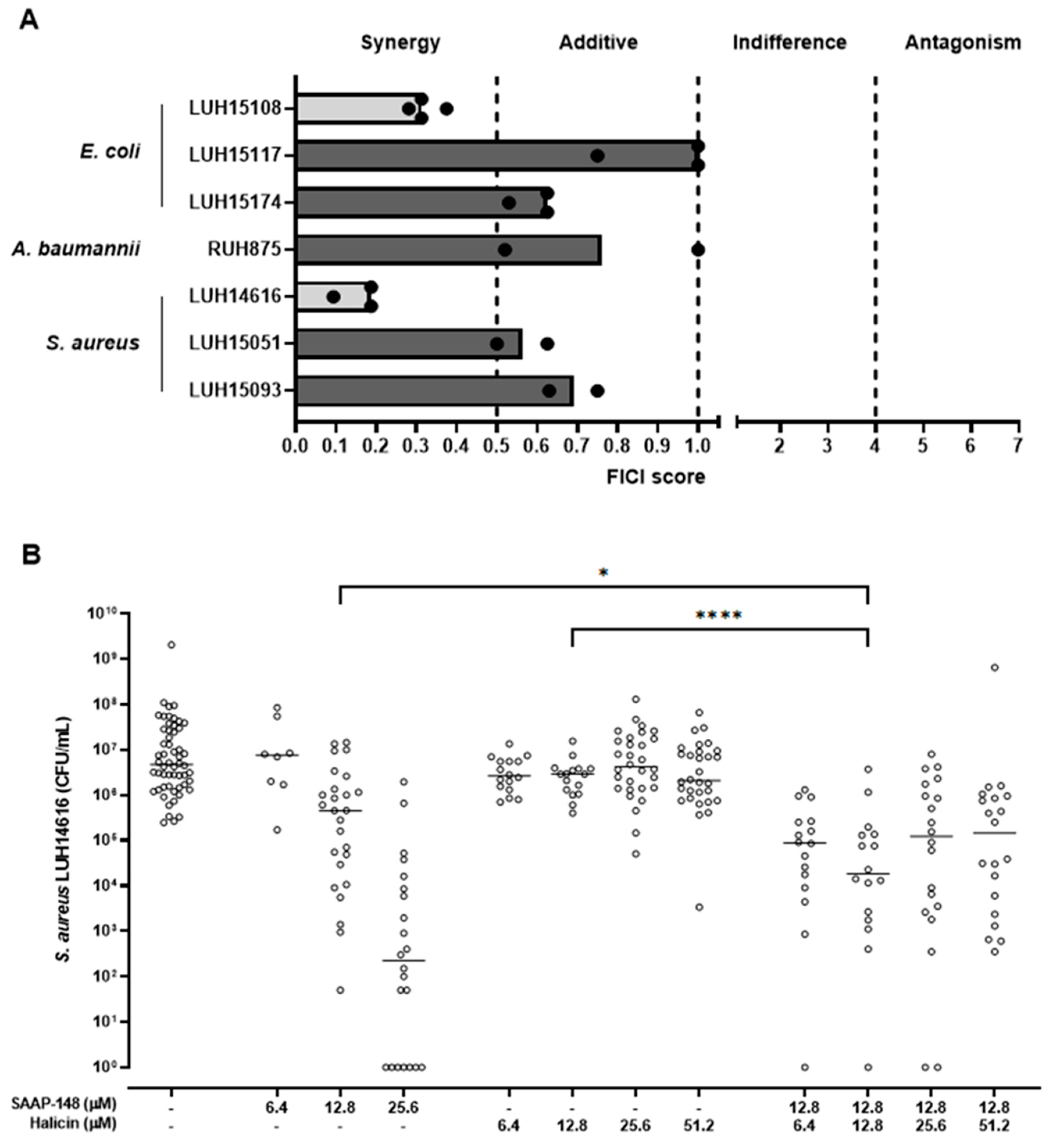
2.4. Favourable Interactions between SAAP-148 and Halicin towards Strains of AMR E. coli and S. aureus
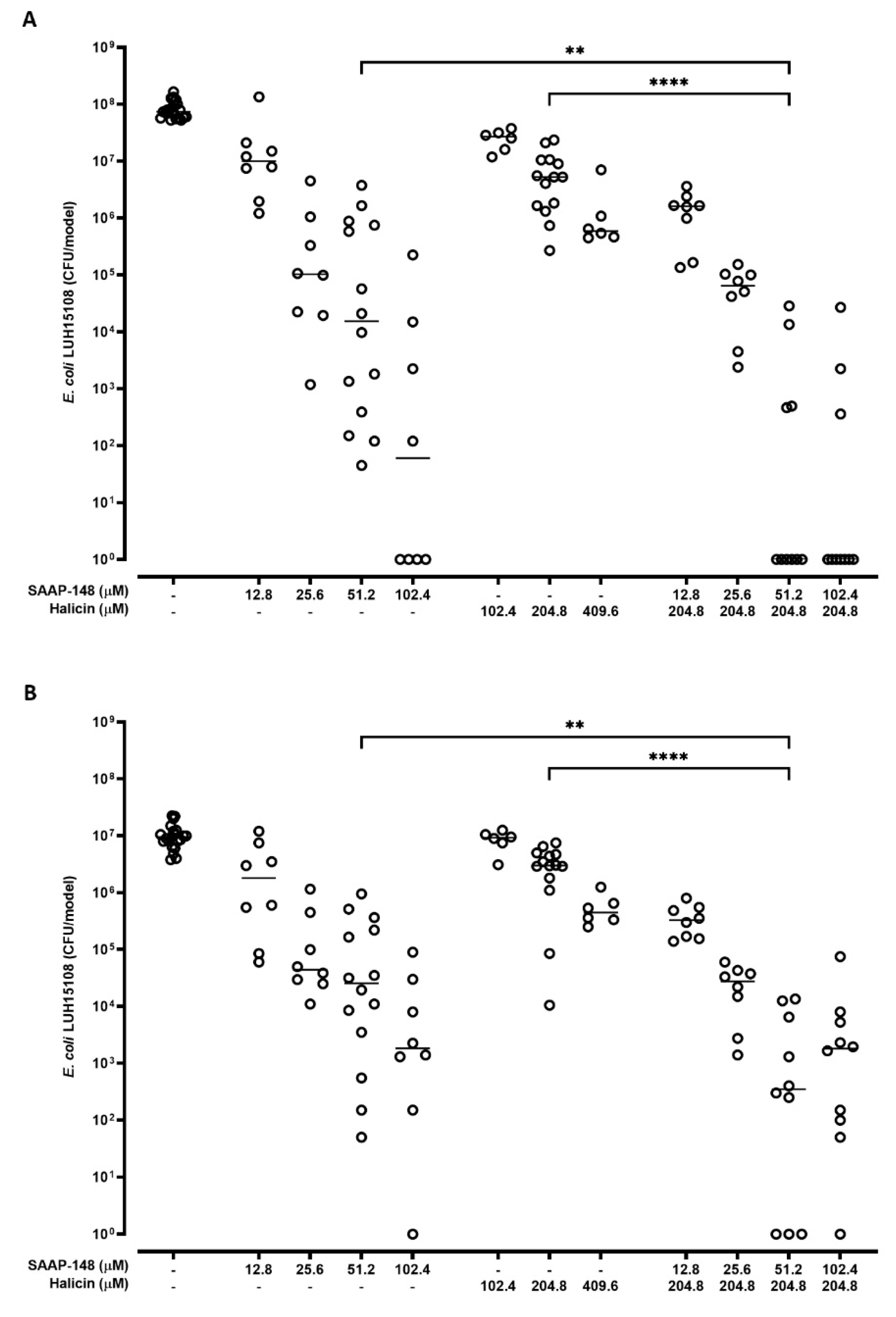
2.5. Effect of Combinations of SAAP-148 and Halicin on AMR E. coli in a 3D Human Bladder Model
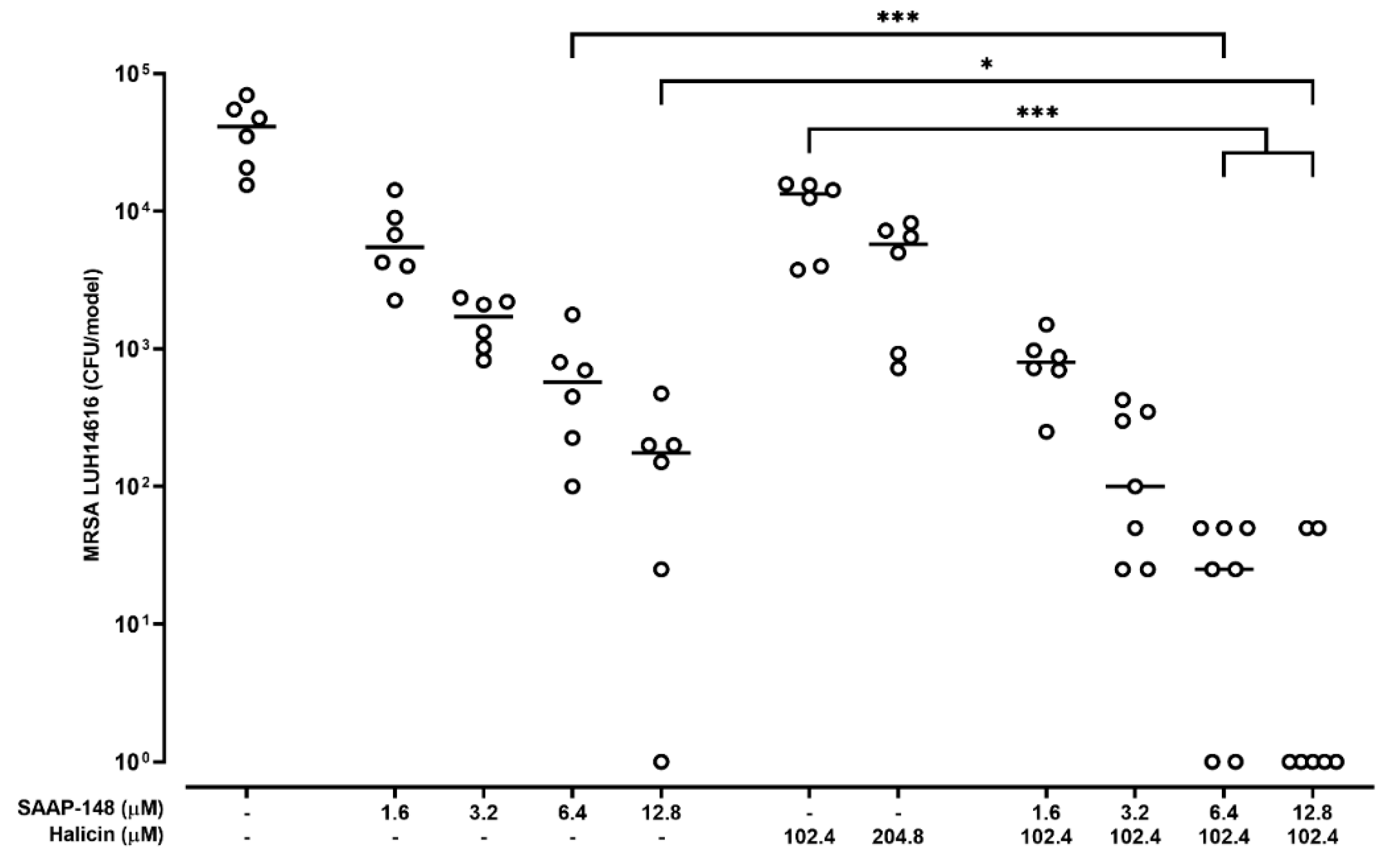
2.6. Effect of Combinations of SAAP-148 and Halicin on AMR S. aureus in a 3D Human Epidermal Model
3. Discussion
4. Materials and Methods
4.1. Antibacterial Agents
4.2. Bacteria
4.3. In Vitro Killing Assay
4.4. Anti-Biofilm Assay
4.5. Hemolysis Assay
4.6. Assays for Cytotoxicity Using Human Primary Skin Fibroblasts and Human RT-4 Urothelial Cells
4.7. Checkerboard Assay
4.8. Biofilm Model on Elastomer Disks
4.9. 3D Bladder Infection Model
4.10. 3D Human Epidermal Infection Model
4.11. Statistics
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McPhillie, M.J.; Cain, R.M.; Narramore, S.; Fishwick, C.W.G.; Simmons, K.J. Computational Methods to Identify New Antibacterial Targets. Chem. Biol. Drug Des. 2014, 85, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Zakeri, B.; Lu, T.K. Synthetic Biology of Antimicrobial Discovery. ACS Synth. Biol. 2012, 2, 358–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloosterman, A.M.; Cimermancic, P.; Elsayed, S.S.; Du, C.; Hadjithomas, M.; Donia, M.S.; Fischbach, M.A.; van Wezel, G.P.; Medema, M.H. Expansion of RiPP biosynthetic space through integration of pan-genomics and machine learning uncovers a novel class of lanthipeptides. PLoS Biol. 2020, 18, e3001026. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C. Mechanisms of Antimicrobial Resistance in Bacteria. Am. J. Med. 2006, 119, S3–S10. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Gerdes, K.; Semsey, S. Microbiology: Pumping persisters. Nature 2016, 534, 41–42. [Google Scholar] [CrossRef] [Green Version]
- De Breij, A.; Riool, M.; Cordfunke, R.A.; Malanovic, N.; de Boer, L.; Koning, R.I.; Ravensbergen, E.; Franken, M.; van der Heijde, T.; Boekema, B.K.; et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci. Transl. Med. 2018, 10, eaan4044. [Google Scholar] [CrossRef] [Green Version]
- Scheper, H.; Wubbolts, J.M.; Verhagen, J.A.M.; de Visser, A.W.; van der Wal, R.J.P.; Visser, L.G.; de Boer, M.G.J.; Nibbering, P.H. SAAP-148 Eradicates MRSA Persisters Within Mature Biofilm Models Simulating Prosthetic Joint Infection. Front. Microbiol. 2021, 12, 99. [Google Scholar] [CrossRef]
- Peek, N.F.A.W.; Nell, M.J.; Brand, R.; Jansen-Werkhoven, T.; van Hoogdalem, E.J.; Verrijk, R.; Vonk, M.J.; Wafelman, A.R.; Valentijn, A.R.P.M.; Frijns, J.H.M.; et al. Ototopical drops containing a novel antibacterial synthetic peptide: Safety and efficacy in adults with chronic suppurative otitis media. PLoS ONE 2020, 15, e0231573. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, M.H.; Orozco, R.Q.; Rezende, S.B.; Rodrigues, G.; Oshiro, K.G.; Cândido, E.S.; Franco, O.L. Computer-Aided Design of Antimicrobial Peptides: Are We Generating Effective Drug Candidates? Front. Microbiol. 2019, 10, 3097. [Google Scholar] [CrossRef]
- Torres, M.T.; de la Fuente-Nunez, C. Toward computer-made artificial antibiotics. Curr. Opin. Microbiol. 2019, 51, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Stokes, J.M.; Yang, K.; Swanson, K.; Jin, W.; Cubillos-Ruiz, A.; Donghia, N.M.; MacNair, C.R.; French, S.; Carfrae, L.A.; Bloom-Ackermann, Z.; et al. A Deep Learning Approach to Antibiotic Discovery. Cell 2020, 181, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Aburayan, W.S.; Booq, R.Y.; BinSaleh, N.S.; Alfassam, H.A.; Bakr, A.A.; Bukhary, H.A.; Alyamani, E.J.; Tawfik, E.A. The Delivery of the Novel Drug ‘Halicin’ Using Electrospun Fibers for the Treatment of Pressure Ulcer against Pathogenic Bacteria. Pharmaceutics 2020, 12, 1189. [Google Scholar] [CrossRef] [PubMed]
- Nussbaumer-Pröll, A.; Zeitlinger, M. Use of Supplemented or Human Material to Simulate PD Behavior of Antibiotics at the Target Site In Vitro. Pharmaceutics 2020, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of Antimicrobial Peptides against Bacterial Biofilms. Materials 2018, 11, 2468. [Google Scholar] [CrossRef] [Green Version]
- Teirlinck, E.; Samal, S.K.; Coenye, T.; Braeckmans, K. Penetrating the bacterial biofilm: Challenges for antimicrobial treatment. In Functionalized Nanomaterials for the Management of Microbial Infection; Elsevier: Amsterdam, The Netherlands, 2017; pp. 49–76. [Google Scholar]
- Stewart, P.S. Diffusion in biofilms. J. Bacteriol. 2003, 185, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Cerca, N.; Gomes, F.; Pereira, S.; Teixeira, P.; Oliveira, R. Confocal laser scanning microscopy analysis of S. epidermidis biofilms exposed to farnesol, vancomycin and rifampicin. BMC Res. Notes 2012, 5, 244. [Google Scholar] [CrossRef] [Green Version]
- Sans-Serramitjana, E.; Jorba, M.; Pedraz, J.L.; Vinuesa, T.; Viñas, M. Determination of the spatiotemporal dependence of Pseudomonas aeruginosa biofilm viability after treatment with NLC-colistin. Int. J. Nanomed. 2017, 12, 4409–4413. [Google Scholar] [CrossRef] [Green Version]
- Pinto, S.N.; Dias, S.A.; Cruz, A.F.; Mil-Homens, D.; Fernandes, F.; Valle, J.; Andreu, D.; Prieto, M.; Castanho, M.A.; Coutinho, A.; et al. The mechanism of action of pepR, a viral-derived peptide, against Staphylococcus aureus biofilms. J. Antimicrob. Chemother. 2019, 74, 2617–2625. [Google Scholar] [CrossRef]
- D’Arpa, P.; Karna, S.L.R.; Chen, T.; Leung, K.P. Pseudomonas aeruginosa transcriptome adaptations from colonization to biofilm infection of skin wounds. Sci. Rep. 2021, 11, 20632. [Google Scholar] [CrossRef]
- Nourbakhsh, F.; Namvar, A.E. Detection of genes involved in biofilm formation in Staphylococcus aureus isolates. GMS Hyg. Infect. Control 2016, 11, Doc07. [Google Scholar] [PubMed]
- Qin, H.; Lo, N.W.; Loo, J.F.; Lin, X.; Yim, A.K.; Tsui, S.K.; Lau, T.C.; Ip, M.; Chan, T.F. Comparative transcriptomics of multidrug-resistant Acinetobacter baumannii in response to antibiotic treatments. Sci. Rep. 2018, 8, 3515. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Gómez, J.M.; Molina-Santiago, C.; Udaondo, Z.; Garitaonaindia, M.T.; Krell, T.; Ramos, J.L.; Daddaoua, A. Full Transcriptomic Response of Pseudomonas aeruginosa to an Inulin-Derived Fructooligosaccharide. Front. Microbiol. 2020, 11, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skindersoe, M.E.; Alhede, M.; Phipps, R.; Yang, L.; Jensen, P.O.; Rasmussen, T.B.; Bjarnsholt, T.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M. Effects of Antibiotics on Quorum Sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2008, 52, 3648–3663. [Google Scholar] [CrossRef] [Green Version]
- De, S.K.; Stebbins, J.L.; Chen, L.H.; Riel-Mehan, M.; Machleidt, T.; Dahl, R.; Yuan, H.; Emdadi, A.; Barile, E.; Chen, V.; et al. Design, Synthesis, and Structure−Activity Relationship of Substrate Competitive, Selective, and In Vivo Active Triazole and Thiadiazole Inhibitors of the c-Jun N-Terminal Kinase. J. Med. Chem. 2009, 52, 1943–1952. [Google Scholar] [CrossRef] [Green Version]
- Munoz, L. Non-kinase targets of protein kinase inhibitors. Nat. Rev. Drug Discov. 2017, 16, 424–440. [Google Scholar] [CrossRef]
- Worthington, R.J.; Richards, J.J.; Melander, C. Small molecule control of bacterial biofilms. Org. Biomol. Chem. 2012, 10, 7457–7474. [Google Scholar] [CrossRef] [Green Version]
- Wilke, K.E.; Carlson, E.E. All Signals Lost. Sci. Transl. Med. 2013, 5, 203ps12. [Google Scholar] [CrossRef] [Green Version]
- Van Esbroeck, A.C.M.; Janssen, A.P.A.; Cognetta, A.B., III; Ogasawara, D.; Shpak, G.; van der Kroeg, M.; Kantae, V.; Baggelaar, M.P.; de Vrij, F.M.S.; Deng, H.; et al. Activity-based protein profiling reveals off-target proteins of the FAAH inhibitor BIA 10-2474. Science 2017, 356, 1084–1087. [Google Scholar] [CrossRef] [Green Version]
- Drouet, M.; Chai, F.; Barthélémy, C.; Lebuffe, G.; Debaene, B.; Décaudin, B.; Odou, P. Influence of Vancomycin Infusion Methods on Endothelial Cell Toxicity. Antimicrob. Agents Chemother. 2015, 59, 930–934. [Google Scholar] [CrossRef] [Green Version]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel, B.D.; Watts, J.L.; Håkansson, J.; Hansen, P.R.; Svenson, J. Correlation between hemolytic activity, cytotoxicity and systemic in vivo toxicity of synthetic antimicrobial peptides. Sci. Rep. 2020, 10, 13206. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Javadov, S. Inhibition of JNK Aggravates the Recovery of Rat Hearts after Global Ischemia: The Role of Mitochondrial JNK. PLoS ONE 2014, 9, e113526. [Google Scholar] [CrossRef] [Green Version]
- Van Gent, M.E.; Ali, M.; Nibbering, P.H.; Kłodzińska, S.N. Current Advances in Lipid and Polymeric Antimicrobial Peptide Delivery Systems and Coatings for the Prevention and Treatment of Bacterial Infections. Pharmaceutics 2021, 13, 1840. [Google Scholar] [CrossRef] [PubMed]
- Makowski, M.; Silva, C.; Pais do Amaral, C.; Gonçalves, S.; Santos, N.C. Advances in Lipid and Metal Nanoparticles for Antimicrobial Peptide Delivery. Pharmaceutics 2019, 11, 588. [Google Scholar] [CrossRef] [Green Version]
- Thapa, R.K.; Diep, D.B.; Tønnesen, H.H. Nanomedicine-based antimicrobial peptide delivery for bacterial infections: Recent advances and future prospects. J. Pharm. Investig. 2021, 51, 377–398. [Google Scholar] [CrossRef]
- Duong, L.; Gross, S.P.; Siryaporn, A. Developing Antimicrobial Synergy with AMPs. Front. Med. Technol. 2021, 3, 9. [Google Scholar] [CrossRef]
- Sheard, D.E.; O’Brien-Simpson, N.M.; Wade, J.D.; Separovic, F. Combating bacterial resistance by combination of antibiotics with antimicrobial peptides. Pure Appl. Chem. 2019, 91, 199–209. [Google Scholar] [CrossRef]
- Koppen, B.C.; Mulder, P.P.G.; de Boer, L.; Riool, M.; Drijfhout, J.W.; Zaat, S.A.J. Synergistic microbicidal effect of cationic antimicrobial peptides and teicoplanin against planktonic and biofilm-encased Staphylococcus aureus. Int. J. Antimicrob. Agents 2019, 53, 143–151. [Google Scholar] [CrossRef]
- Li, S.; She, P.; Zhou, L.; Zeng, X.; Xu, L.; Liu, Y.; Chen, L.; Wu, Y. High-Throughput Identification of Antibacterials against Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 3109. [Google Scholar] [CrossRef]
- Huang, S.-M.; Wu, C.S.; Chiu, M.H.; Wu, C.H.; Chang, Y.T.; Chen, G.S.; Lan, C.C.E. High glucose environment induces M1 macrophage polarization that impairs keratinocyte migration via TNF-α: An important mechanism to delay the diabetic wound healing. J. Dermatol. Sci. 2019, 96, 159–167. [Google Scholar] [CrossRef]
- Wu, Y.; Quan, Y.; Liu, Y.; Liu, K.; Li, H.; Jiang, Z.; Zhang, T.; Lei, H.; Radek, K.A.; Li, D.; et al. Hyperglycaemia inhibits REG3A expression to exacerbate TLR3-mediated skin inflammation in diabetes. Nat. Commun. 2016, 7, 13393. [Google Scholar] [CrossRef] [PubMed]
- Pietropaolo, A.; Jones, P.; Moors, M.; Birch, B.; Somani, B.K. Use and Effectiveness of Antimicrobial Intravesical Treatment for Prophylaxis and Treatment of Recurrent Urinary Tract Infections (UTIs): A Systematic Review. Curr. Urol. Rep. 2018, 19, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Miao, Y.; Abraham, S.N. The multiple antibacterial activities of the bladder epithelium. Ann. Transl. Med. 2017, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Cal, P.M.; Matos, M.J.; Bernardes, G.J. Trends in therapeutic drug conjugates for bacterial diseases: A patent review. Expert Opin. Ther. Pat. 2017, 27, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, H.S.; Duinkerken, G.; Benckhuijsen, W.E.; Amons, R.; de Vries, R.R.P.; Roep, B.O.; Drijfhout, J.W. The identification of CD4+ T cell epitopes with dedicated synthetic peptide libraries. Proc. Natl. Acad. Sci. USA 1997, 94, 10313–10318. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente-Nunez, C.; Korolik, V.; Bains, M.; Nguyen, U.; Breidenstein, E.B.M.; Horsman, S.; Lewenza, S.; Burrows, L.; Hancock, R.E.W. Inhibition of bacterial biofilm formation and swarming motility by a small synthetic cationic peptide. Antimicrob. Agents Chemother. 2012, 56, 2696–2704. [Google Scholar] [CrossRef] [Green Version]
- Doern, C.D. When Does 2 Plus 2 Equal 5? A Review of Antimicrobial Synergy Testing. J. Clin. Microbiol. 2014, 52, 4124–4128. [Google Scholar] [CrossRef] [Green Version]
- Smits, J.P.H.; Niehues, H.; Rikken, G.; van Vlijmen-Willems, I.M.J.J.; van de Zande, G.W.H.J.F.; Zeeuwen, P.L.J.M.; Schalkwijk, J.; van den Bogaard, E.H. Immortalized N/TERT keratinocytes as an alternative cell source in 3D human epidermal models. Sci. Rep. 2017, 7, 11838. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | LC99.9 SAAP-148 (µM) | LC99.9 Halicin (µM) | |||||
|---|---|---|---|---|---|---|---|---|
| PBS | 50% Urine/Plasma | PBS | 50% Urine/Plasma | |||||
| E. coli | LUH15174 | 0.8 | 1.6 | (0.8–1.6) | 12.8 | (6.4–51.2) | 51.2 | (6.4–>51.2) |
| A. baumannii | RUH875 | 1.6 | 6.4 | 25.6 | (25.6–51.2) | >102.4 | ||
| S. aureus | LUH14616 | 1.6 | 12.8 | 102.4 | (102.4–>102.4) | >204.8 | ||
| Species | Strain | BIC99 SAAP-148 (µM) | BIC99 Halicin (µM) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 24 h Biofilm | 7 Days Biofilm | 24 h Biofilm | 7 Days Biofilm | ||||||
| 4 h | 24 h | 4 h | 24 h | 4 h | 24 h | 4 h | 24 h | ||
| E. coli | LUH15174 | 204.8 | 68.3 | 204.8 | 68.3 | 204.8 | 68.3 | 204.8 | ≤22.8 |
| (68.3–204.8) | (68.3–204.8) | (204.8–>204.8) | (≤22.8–68.3) | - | (68.3–204.8) | - | (≤22.8–68.3) | ||
| A. baumannii | RUH875 | 68.3 | 204.8 | 68.3 | 68.3 | 204.8 | >204.8 | 204.8 | 68.3 |
| - | - | - | - | - | (204.8–>204.8) | - | (68.3–204.8) | ||
| S. aureus | LUH14616 | 204.8 | 204.8 | 68.3 | 68.3 | >204.8 | 68.3 | >204.8 | 68.3 |
| (68.3–204.8) | - | - | - | - | (68.3–204.8) | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Gent, M.E.; van der Reijden, T.J.K.; Lennard, P.R.; de Visser, A.W.; Schonkeren-Ravensbergen, B.; Dolezal, N.; Cordfunke, R.A.; Drijfhout, J.W.; Nibbering, P.H. Synergism between the Synthetic Antibacterial and Antibiofilm Peptide (SAAP)-148 and Halicin. Antibiotics 2022, 11, 673. https://doi.org/10.3390/antibiotics11050673
van Gent ME, van der Reijden TJK, Lennard PR, de Visser AW, Schonkeren-Ravensbergen B, Dolezal N, Cordfunke RA, Drijfhout JW, Nibbering PH. Synergism between the Synthetic Antibacterial and Antibiofilm Peptide (SAAP)-148 and Halicin. Antibiotics. 2022; 11(5):673. https://doi.org/10.3390/antibiotics11050673
Chicago/Turabian Stylevan Gent, Miriam E., Tanny J. K. van der Reijden, Patrick R. Lennard, Adriëtte W. de Visser, Bep Schonkeren-Ravensbergen, Natasja Dolezal, Robert A. Cordfunke, Jan Wouter Drijfhout, and Peter H. Nibbering. 2022. "Synergism between the Synthetic Antibacterial and Antibiofilm Peptide (SAAP)-148 and Halicin" Antibiotics 11, no. 5: 673. https://doi.org/10.3390/antibiotics11050673