Engineered Biosensors for Diagnosing Multidrug Resistance in Microbial and Malignant Cells

 , and
, and
Abstract
:1. Introduction
2. Multidrug Resistance in Microorganisms and Biosensors
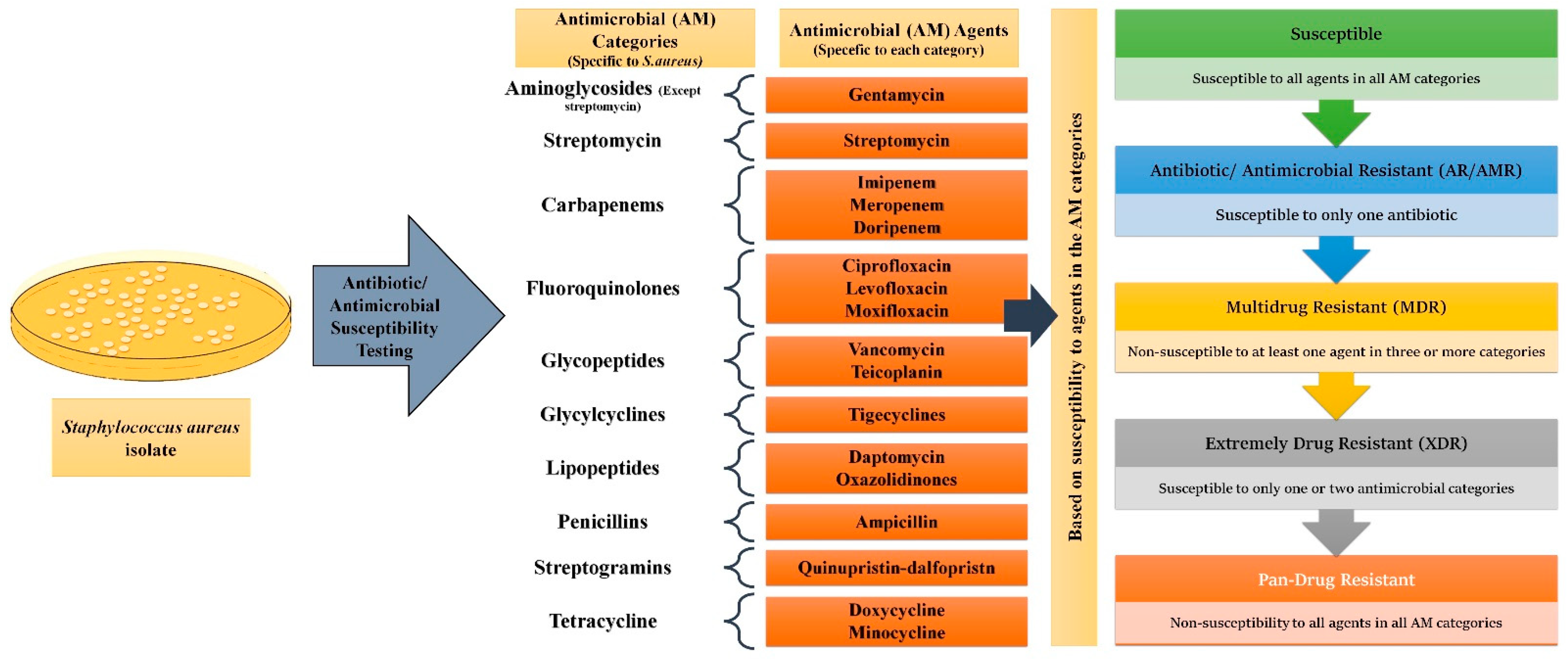
2.1. MDR Glossary and Origin of Concept
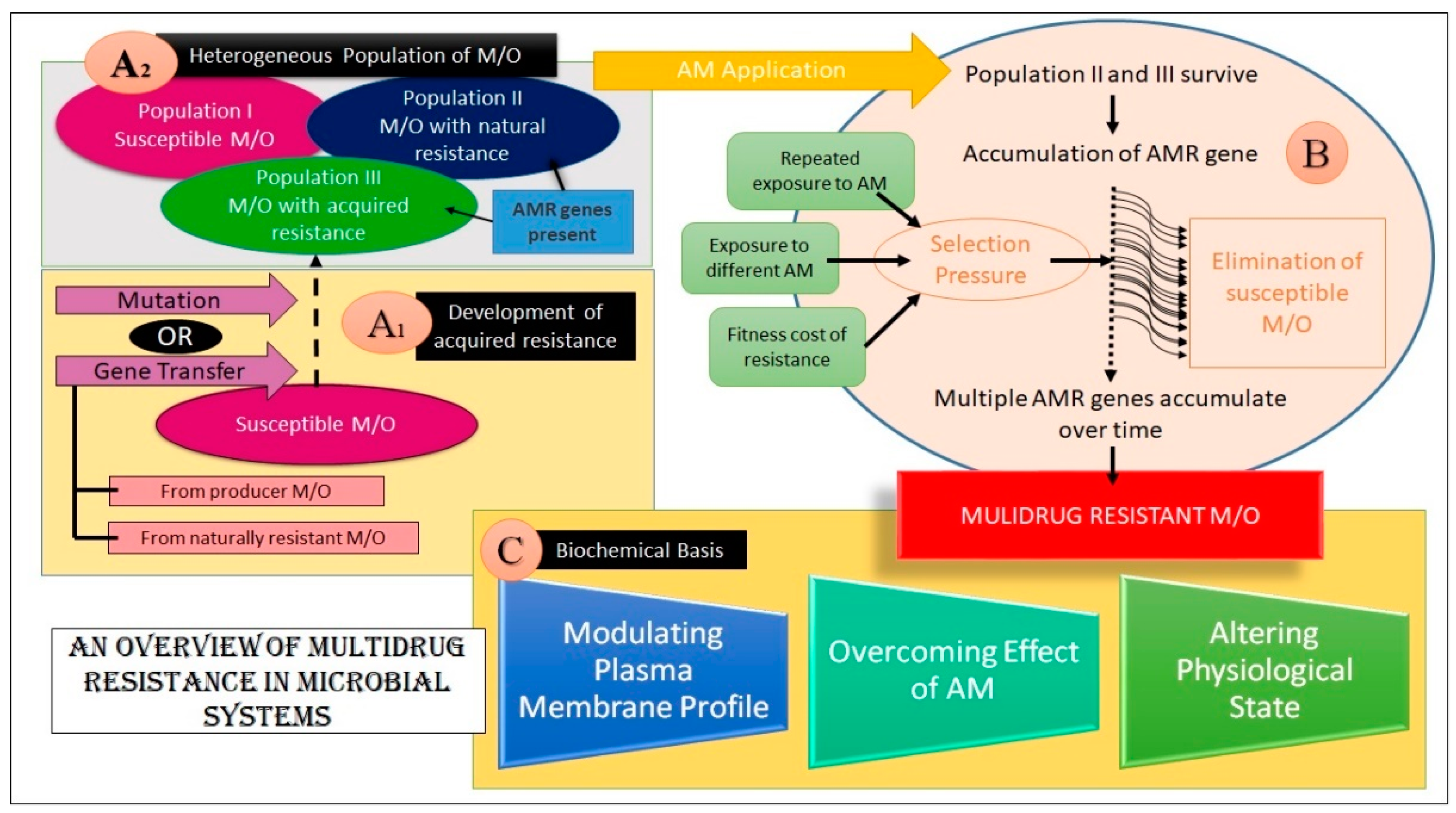
2.2. Foundation of the Emergence of MDR
2.3. Biochemical Basis of MDR in Microbes
2.3.1. Alteration in the PM Profile
2.3.2. Overcoming the Drug Action
2.3.3. Alteration in the Physiological State
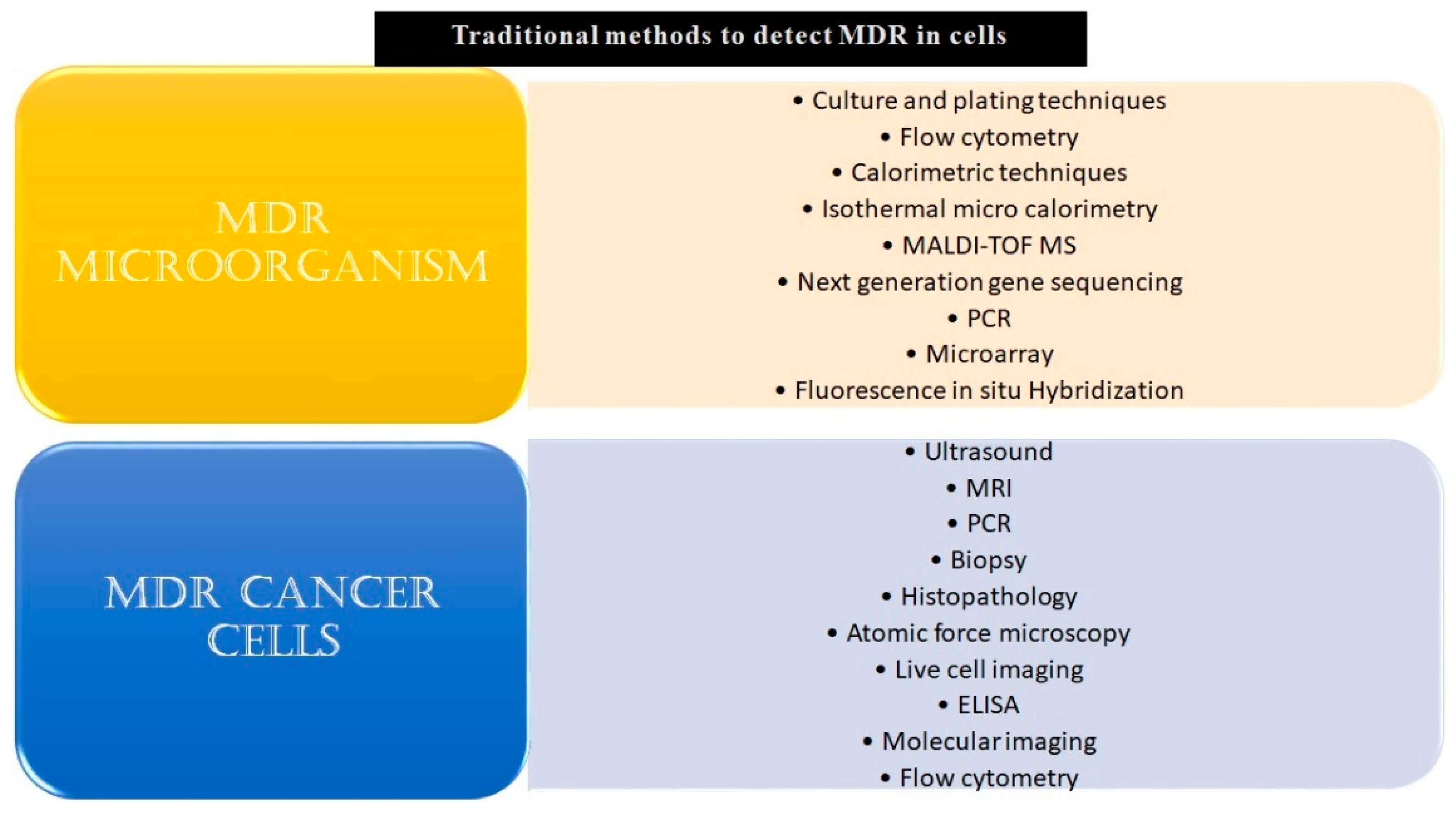
2.4. Use of Biosensors in Detecting MDR Microbes
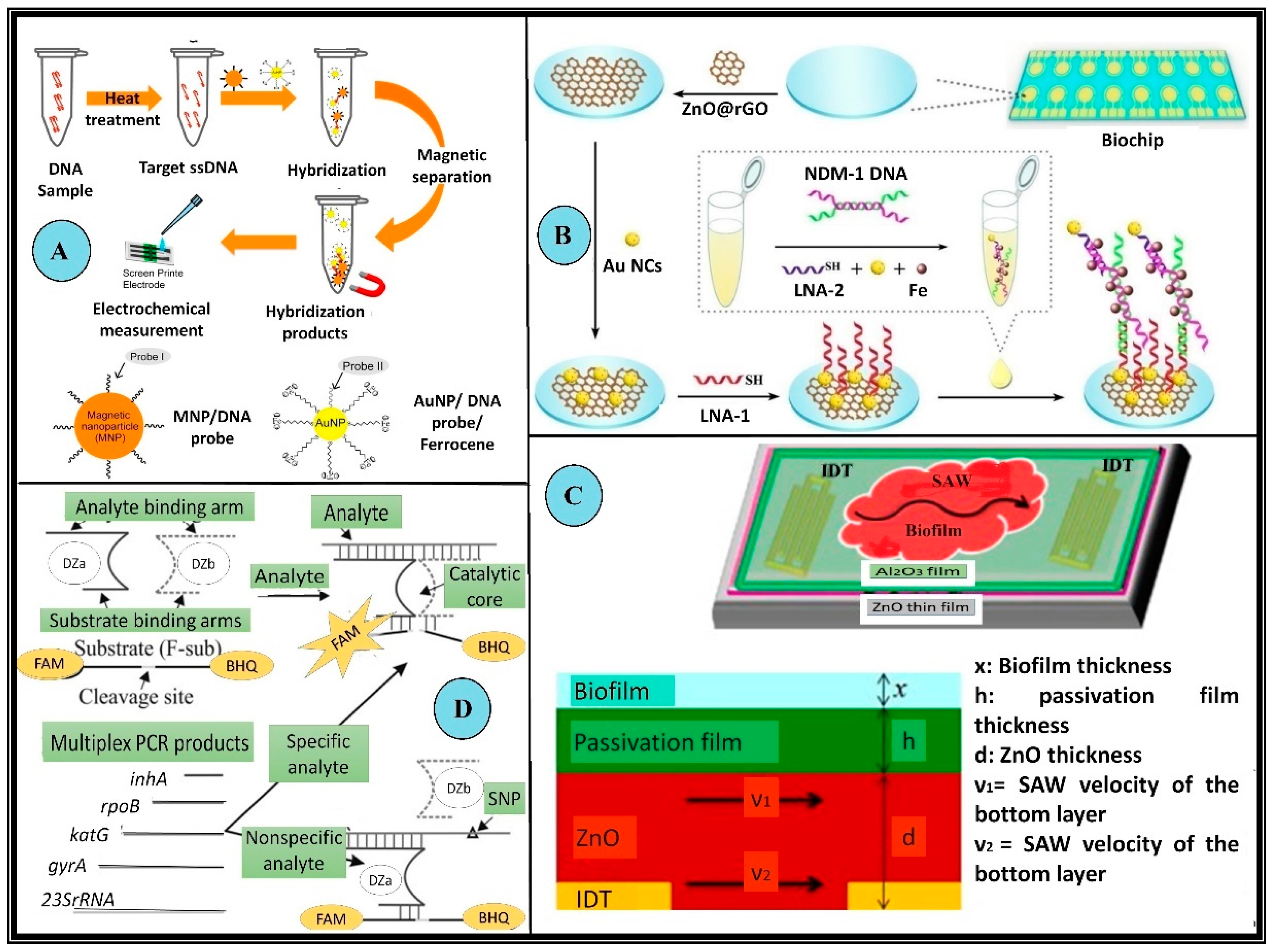
2.4.1. Genotypic Biosensor for MDR Microbes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Form Associated with MDR | Function | Resistance Against | Reference |
|---|---|---|---|---|
| mecA | Normal | Codes for alternative penicillin-binding protein PBP2a | Methicillin, nafcillin, oxacillin, and cephalosporins | [46] |
| rpoB | Mutated | Codes for the β-subunit of RNA polymerase | Rifampicin and isoniazid in multidrug resistant Mycobacterium tuberculosis | [47,48,49] |
| ampR | Normal | Involved in β-lactamase transcription, a transcriptional activator of the lysR family | 3rd generation cephalosporinase | [50,51] |
| katG | Mutated | Codes for the catalase-peroxidase enzyme. | Isoniazid in M. tuberculosis when loss of function of the gene is seen | [52,53] |
| gyrA | Mutated | Codes for GyrA protein or DNA gyrase, a target of quinolones | Quinolones | [54,55] |
| inhA | Mutated | Codes for enoyl-ACP reductase of type II fatty acid synthase. which is crucial for the biosynthesis of mycolic acid (a component of the cell wall of Mycobacterium) | Isoniazid | [56,57] |
| hlyA | Normal | Codes for extracellular hyaluronate lyase Codes for α-hemolysin Shows enhanced virulence | -- | [58,59] |
| YMDD motif in reverse transcriptase | Mutated YMDD motif | Locus/motif present in RNA-dependent DNA polymerase. | Lamivudine in Hepatitis B virus | [60,61] |
| K13 gene | C580Y mutation | Codes for Kelch protein | Artemisinin in Plasmodium falciparum | [62] |
| NDM1 gene(blaNDM-1) | Normal and variants | Codes for New Delhi metallo-β-lactamase-1 (NDM-1) | Resistance to carbapeneme and β lactam antibiotics (except for azetreonam) | [45,63] |
| gliT gene | Normal | Codes for gliotoxin A virulence factor associated with invasive aspergillosis | -- | [64,65] |
2.4.2. Phenotypic Biosensors for MDR Microbes
3. Multidrug Resistance in Malignant Systems and Biosensors
3.1. Foundation and Emergence of MDR in Cancer
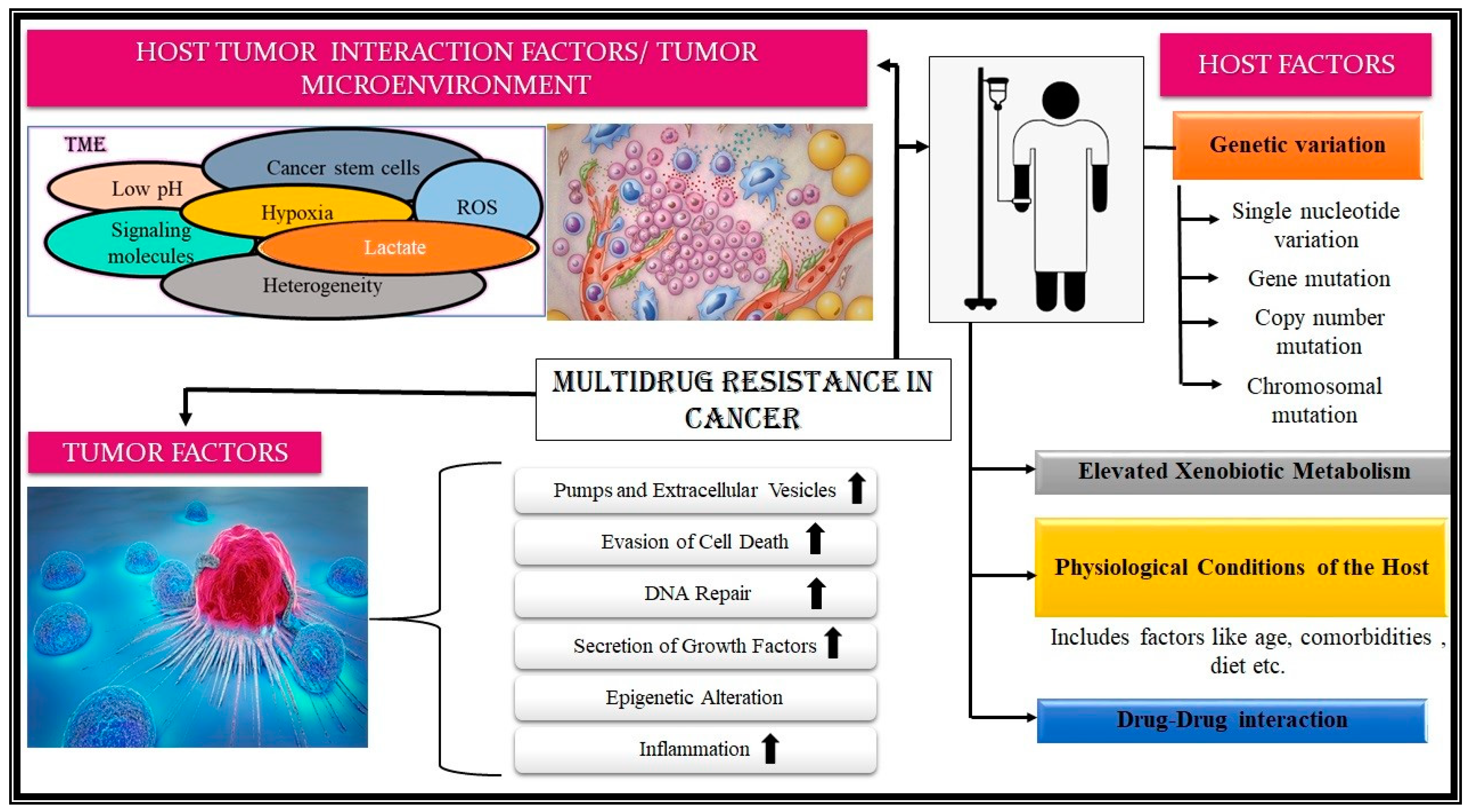
3.2. Biochemical Basis of MDR in Cancer
3.2.1. Host-Associated Factors
3.2.2. Tumor-Associated Factors
3.2.3. Host-Tumor-Interaction Associated Factor

3.3. Use of Biosensors in Detecting MDR in Neoplasms
3.3.1. Genotypic Biosensor for MDR Cancer
3.3.2. Phenotypic Biosensor for MDR Cancer
3.3.3. Drug Pharmacokinetics-Based Biosensors for MDR Cancer
4. Discussion and Future Perspectives
4.1. What Kind of Threat Does MDR Pose?
4.2. What Is the Current Scenario of Diagnosis?
4.3. How can Biosensors Be Used to Address These Problems?
4.4. Possible Roadblocks and Shortcomings
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention (2019). AR Threats Report. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 24 July 2022).
- Barie, P.S. Multidrug-Resistant Organisms and Antibiotic Management. Surg. Clin. N. Am. 2012, 92, 345–391. [Google Scholar] [CrossRef]
- Menkem, E.; Lemalue, B.; Shinwin, S.; Tamunjoh, A.; Fabrice, F. Antibiotic Residues in Food Animals: Public Health Concern. Acta Ecol. Sin. 2019, 39, 411–415. [Google Scholar] [CrossRef]
- Morehead, M.S.; Scarbrough, C. Emergence of Global Antibiotic Resistance. Prim. Care—Clin. Off. Pract. 2018, 45, 467–484. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. In Current Opinion in Microbiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 51, pp. 72–80. [Google Scholar] [CrossRef]
- Wu, D.; Ding, Y.; Yao, K.; Gao, W.; Wang, Y. Antimicrobial Resistance Analysis of Clinical Escherichia Coli Isolates in Neonatal Ward. Front. Pediatr. 2021, 9, 670470. [Google Scholar] [CrossRef]
- Li, Y.J.; Lei, Y.H.; Yao, N.; Wang, C.R.; Hu, N.; Ye, W.C.; Zhang, D.M.; Chen, Z.S. Autophagy and multidrug resistance in cancer. Chin. J. Cancer 2017, 36, 52. [Google Scholar] [CrossRef]
- Ye, Q.; Liu, K.; Shen, Q.; Li, Q.; Hao, J.; Han, F.; Jiang, R.W. Reversal of multidrug resistance in cancer by multi-functional flavonoids. Front. Oncol. 2019, 9, 487. [Google Scholar] [CrossRef]
- Baguley, B.C. Chapter 1 Multidrug Resistance in Cancer; Springer International Publishing: Berlin/Heidelberg, Germany, 2010; Volume 596. [Google Scholar] [CrossRef]
- Kunjachan, S.; Rychlik, B.; Storm, G.; Kiessling, F.; Lammers, T. Multidrug resistance: Physiological principles and nanomedical solutions. In Advanced Drug Delivery Reviews; Elsevier: Amsterdam, The Netherlands, 2013; Volume 65, pp. 1852–1865. [Google Scholar] [CrossRef]
- Mahato, K.; Maurya, P.K.; Chandra, P. Fundamentals and commercial aspects of nanobiosensors in point-of-care clinical diagnostics. 3 Biotech. 2018, 8, 149. [Google Scholar] [CrossRef]
- Cui, F.; Ye, Y.; Ping, J.; Sun, X. Biosensors and Bioelectronics Carbon dots: Current advances in pathogenic bacteria monitoring and prospect applications. Biosens. Bioelectron. 2020, 156, 112085. [Google Scholar] [CrossRef]
- Gupta, S.; Kakkar, V. Recent technological advancements in tuberculosis diagnostics—A review. Biosens. Bioelectron. 2018, 115, 14–29. [Google Scholar] [CrossRef]
- Alafeef, M.; Moitra, P.; Pan, D. Nano-enabled sensing approaches for pathogenic bacterial detection. Biosens. Bioelectron. 2020, 165, 112276. [Google Scholar] [CrossRef]
- Jayanthi, V.S.P.K.S.A.; Das, A.B.; Saxena, U. Recent advances in biosensor development for the detection of cancer biomarkers. Biosens. Bioelectron. 2017, 91, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Kaur, B.; Kumar, S.; Kaushik, B.K. Recent advancements in optical biosensors for cancer detection. Biosens. Bioelectron. 2022, 197, 113805. [Google Scholar] [CrossRef]
- Kumar, A.; Purohit, B.; Maurya, P.K.; Pandey, L.M.; Chandra, P. Engineered nanomaterial assisted signal-amplification strategies for enhancing analytical performance of electrochemical biosensors. Electroanalysis 2019, 31, 1615–1629. [Google Scholar] [CrossRef]
- Andrei, L.; Kasas, S.; Ochoa Garrido, I.; Stanković, T.; Suárez Korsnes, M.; Vaclavikova, R.; Assaraf, Y.G.; Pešić, M. Advanced Technological Tools to Study Multidrug Resistance in Cancer. Drug Resist. Updat. 2020, 48, 100658. [Google Scholar] [CrossRef]
- Zhu, Y.; Chandra, P.; Song, K.M.; Ban, C.; Shim, Y.B. Label-Free Detection of Kanamycin Based on the Aptamer-Functionalized Conducting Polymer/Gold Nanocomposite. Biosens. Bioelectron. 2012, 36, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Noh, H.B.; Shim, Y.B. Cancer Cell Detection Based on the Interaction between an Anticancer Drug and Cell Membrane Components. Chem. Commun. 2013, 49, 1900–1902. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Majdinasab, M.; Khan, R.; Li, Z.; Hayat, A.; Marty, J.L. Nanomaterials in fluorescence-based biosensors: Defining key roles. Nano-Struct. Nano-Objects 2021, 27, 100774. [Google Scholar] [CrossRef]
- Mandal, R.; Baranwal, A.; Srivastava, A.; Chandra, P. Evolving trends in bio/chemical sensor fabrication incorporating bimetallic nanoparticles. In Biosensors and Bioelectronics; Elsevier: Amsterdam, The Netherlands, 2018; Volume 117, pp. 546–561. [Google Scholar] [CrossRef]
- Purohit, B.; Vernekar, P.R.; Shetti, N.P.; Chandra, P. Biosensor Nanoengineering: Design, Operation, and Implementation for Biomolecular Analysis. Sens. Int. 2020, 1, 100040. [Google Scholar] [CrossRef]
- Gupta, N.; Renugopalakrishnan, V.; Liepmann, D.; Paulmurugan, R.; Malhotra, B.D. Cell-Based Biosensors: Recent Trends, Challenges and Future Perspectives. Biosens. Bioelectron. 2019, 141, 111435. [Google Scholar] [CrossRef]
- Mahato, K.; Kumar, A.; Maurya, P.K.; Chandra, P. Shifting Paradigm of Cancer Diagnoses in Clinically Relevant Samples Based on Miniaturized Electrochemical Nanobiosensors and Microfluidic Devices. Biosens. Bioelectron. 2018, 100, 411–428. [Google Scholar] [CrossRef]
- Tech, J.A.B.; Mahato, K.; Prasad, A.; Maurya, P.K.; Chandra, P. Nanobiosensors: Next Generation Point-of-Care Biomedical Devices for Personalized Diagnosis. J. Anal. Bioanal. Tech. 2016, 7, e125. [Google Scholar] [CrossRef]
- Ullah, H.; Ali, S. [Online] Classification of antibacterial agents and their functions. Intech 1989, 32, 137–144. Available online: https://www.intechopen.com/books/advanced-biometric-technologies/liveness-detection-in-biometrics (accessed on 8 January 2022).
- Basak, S.; Singh, P.; Rajurkar, M. Multidrug Resistant and Extensively Drug Resistant Bacteria: A Study. J. Pathog. 2016, 2016, 4065603. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef]
- Ray, S.M.S.; Das, S. Molecular Mechanism of Drug Resistance. In Drug Resistance in Bacteria. Fungi, Malaria, and Cancer; Springer: Cham, Switzerland, 2017; pp. 47–100. [Google Scholar] [CrossRef]
- Jacopin, E.; Lehtinen, S.; Débarre, F.; Blanquart, F. Factors Favouring the Evolution of Multidrug Resistance in Bacteria. J. R Soc. Interface 2020, 17, 20200105. [Google Scholar] [CrossRef]
- Conteduca, D.; Brunetti, G.; Dell’Olio, F.; Armenise, M.N.; Krauss, T.F.; Ciminelli, C. Monitoring of Individual Bacteria Using Electro-Photonic Traps. Biomed. Opt. Express 2019, 10, 3463. [Google Scholar] [CrossRef]
- Chang, H.-H.; Cohen, T.; Grad, Y.H.; Hanage, W.P.; O’Brien, T.F.; Lipsitch, M. Origin and Proliferation of Multiple-Drug Resistance in Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2015, 79, 101–116. [Google Scholar] [CrossRef]
- Lubelski, J.; Konings, W.N.; Driessen, A.J.M. Distribution and Physiology of ABC-Type Transporters Contributing to Multidrug Resistance in Bacteria. Microbiol. Mol. Biol. Rev. 2007, 71, 463–476. [Google Scholar] [CrossRef]
- Tanwar, J.; Das, S.; Fatima, Z.; Hameed, S. Multidrug Resistance: An Emerging Crisis. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 541340. [Google Scholar] [CrossRef]
- Arora, G.; Sajid, A.; Kalia, V.C. Drug Resistance in Bacteria, Fungi, Malaria, and Cancer; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; ISBN 9783319486833. [Google Scholar] [CrossRef]
- Subhadra, B.; Hwan Oh, M.; Hee Choi, C. Quorum Sensing in Acinetobacter: With Special Emphasis on Antibiotic Resistance, Biofilm Formation and Quorum Quenching. AIMS Microbiol. 2016, 2, 27–41. [Google Scholar] [CrossRef]
- Bardbari, A.M.; Arabestani, M.R.; Karami, M.; Keramat, F.; Alikhani, M.Y.; Bagheri, K.P. Correlation between Ability of Biofilm Formation with Their Responsible Genes and MDR Patterns in Clinical and Environmental Acinetobacter Baumannii Isolates. Microb. Pathog. 2017, 108, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.R. The challenge of multidrug resistance: Actual strategies in the development of novel antibacterials. Appl. Microbiol. Biotechnol. 2004, 63, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. In Antimicrobial Resistance and Infection Control; BioMed Central Ltd.: London, UK, 2019; Volume 8, p. 76. [Google Scholar] [CrossRef]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial Persister Cell Formation and Dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Kuwata, N.; Sakamoto, H.; Amano, Y.; Satomura, T.; Suye, S.I. A Smart DNA Sensing System for Detecting Methicillin-Resistant Staphylococcus Aureus Using Modified Nanoparticle Probes. Biosens. Bioelectron. 2015, 67, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hedström, M.; Chen, D.; Fan, X.; Mattiasson, B. A Capacitive DNA Sensor-Based Test for Simple and Sensitive Analysis of Antibiotic Resistance in Field Setting. Biosens. Bioelectron. 2015, 64, 255–259. [Google Scholar] [CrossRef]
- Zhang, L.; Liang, W.; Ran, Q.; Liu, F.; Chen, D.; Xiong, Y.; Jiang, D.; Li, Y.; Tu, D.; Luo, X.; et al. Ultrasensitive Detection of NDM-1 Resistant Bacteria Based on Signal Amplification with Sandwich-Type LNA Electrochemical Biochips. Sens. Actuators B Chem. 2020, 306, 127556. [Google Scholar] [CrossRef]
- Opal, S.M.; Pop-Vicas, A. Molecular mechanisms of antibiotic resistance in bacteria. In Eighth Edi; Elsevier, Inc.: Amsterdam, The Netherlands, 2014; Volume 1. [Google Scholar] [CrossRef]
- Sinha, P.; Srivastava, G.N.; Tripathi, R.; Mishra, M.N.; Anupurba, S. Detection of Mutations in the RpoB Gene of Rifampicin-Resistant Mycobacterium Tuberculosis Strains Inhibiting Wild Type Probe Hybridization in the MTBDR plus Assay by DNA Sequencing Directly from Clinical Specimens. BMC Microbiol. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Makadia, J.S.; Jain, A.; Patra, S.K.; Sherwal, B.L.; Khanna, A. Emerging Trend of Mutation Profile of RpoB Gene in MDR Tuberculosis, North India. Indian J. Clin. Biochem. 2012, 27, 370–374. [Google Scholar] [CrossRef]
- Adékambi, T.; Drancourt, M.; Raoult, D. The RpoB Gene as a Tool for Clinical Microbiologists. Trends Microbiol. 2009, 17, 37–45. [Google Scholar] [CrossRef]
- Lodge, J.; Busby, S.; Piddock, L. Investigation of the Pseudomonas Aeruginosa AmpR Gene and Its Role at the Chromosomal AmpC β-Lactamase Promoter. FEMS Microbiol. Lett. 1993, 111, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, J.G. Nucleotide Sequence of the Ampicillin Resistance Gene of Escherichia Coli Plasmid PBR322. Proc. Natl. Acad. Sci. USA 1978, 75, 3737–3741. [Google Scholar] [CrossRef] [PubMed]
- Jaber, M.; Rattan, A.; Kumar, R. Presence of KatG Gene in Resistant Mycobacterium Tuberculosis. J. Clin. Pathol. 1996, 49, 945–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivekar, S.S.; Kaliaperumal, V.; Brammacharry, U.; Sakkaravarthy, A.; Raj, C.K.V.; Alagappan, C.; Muthaiah, M. Prevalence and Factors Associated with Multidrug-Resistant Tuberculosis in South India. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jaktaji, R.P.; Mohiti, E. Study of Mutations in the DNA Gyrase GyrA Gene of Escherichia Coli. Iran. J. Pharm. Res. 2010, 9, 43–48. [Google Scholar]
- Johnning, A.; Kristiansson, E.; Fick, J.; Weijdegård, B.; Larsson, D.G.J. Resistance Mutations in GyrA and ParC Are Common in Escherichia Communities of Both Fluoroquinolone-Polluted and Uncontaminated Aquatic Environments. Front. Microbiol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Dubnau, E.; Quemard, A.; Balasubramanian, V.; Um, K.S.; Wilson, T.; Collins, D.; de Lisle, G.; Jr, W.R.J. InhA, Encoding Target Mycobacterium. Science 1993, 263, 227–230. [Google Scholar] [CrossRef]
- Uniprot UniProtKB—P9WGR1 (INHA_MYCTU). Available online: https://www.uniprot.org/uniprot/P9WGR1 (accessed on 23 March 2022).
- Hynes, W.L.; Dixon, A.R.; Walton, S.L.; Aridgides, L.J. The Extracellular Hyaluronidase Gene (HylA) of Streptococcus Pyogenes. FEMS Microbiol. Lett. 2000, 184, 109–112. [Google Scholar] [CrossRef]
- Yue, S.; Zhang, Z.; Liu, Y.; Zhou, Y.; Wu, C.; Huang, W.; Chen, N.; Zhu, Z. Phenotypic and Molecular Characterizations of Multidrug-Resistant Diarrheagenic E. coli of Calf Origin. Anim. Dis. 2021, 1, 1–13. [Google Scholar] [CrossRef]
- Warner, N.; Locarnini, S. Mechanisms of Hepatitis B Virus Resistance Development. Intervirology 2014, 57, 218–224. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Wei, M.; Zhang, C.; Xu, T.; Liu, L.; Xu, Z. Potential Resistant Mutations within HBV Reverse Transcriptase Sequences in Nucleos(t)Ide Analogues-Experienced Patients with Hepatitis B Virus Infection. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zaw, M.T.; Lin, Z.; Emran, N.A. Importance of Kelch 13 C580Y Mutation in the Studies of Artemisinin Resistance in Plasmodium Falciparum in Greater Mekong Subregion. J. Microbiol. Immunol. Infect. 2020, 53, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.U.; Maryam, L.; Zarrilli, R. Structure, Genetics and Worldwide Spread of New Delhi Metallo- β -Lactamase (NDM): A Threat to Public Health. BMC Microbiol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Sugui, J.A. What Do We Know about the Role of Gliotoxin in the Pathobiology of Aspergillus Fumigatus? Med. Mycol. 2009, 47, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Kosmidis, C.; Denning, D.W. The Clinical Spectrum of Pulmonary Aspergillosis. Thorax 2015, 70, 270–277. [Google Scholar] [CrossRef]
- Haddaoui, M.; Sola, C.; Raouafi, N.; Korri-Youssoufi, H. E-DNA Detection of RpoB Gene Resistance in Mycobacterium Tuberculosis in Real Samples Using Fe 3 O 4 /Polypyrrole Nanocomposite. Biosens. Bioelectron. 2019, 128, 76–82. [Google Scholar] [CrossRef]
- Khammanee, T.; Sawangjaroen, N.; Buncherd, H.; Tun, A.W. A LAMP-SNP Assay Detecting C580Y Mutation in Pfkelch13 Gene from Clinically Dried Blood Spot Samples. Korean J. Parasitol. 2021, 59, 15–22. [Google Scholar] [CrossRef]
- Malpartida-Cardenas, K.; Miscourides, N.; Rodriguez-Manzano, J.; Yu, L.S.; Moser, N.; Baum, J.; Georgiou, P. Quantitative and Rapid Plasmodium Falciparum Malaria Diagnosis and Artemisinin-Resistance Detection Using a CMOS Lab-on-Chip Platform. Biosens. Bioelectron. 2019, 145, 111678. [Google Scholar] [CrossRef]
- Heliyaneh Moeinizadeh, M.S. Frequency of hlyA, hlyB, hlyC and hlyD genes in uropathogenic Escherichia coli isolated from UTI patients in Shiraz. GMS Hyg. Infect. Control 2021, 16, 396. [Google Scholar] [CrossRef]
- Shi, D.; Shi, H. Combining Loop-Mediated Isothermal Amplification and Nanozyme-Strip for Ultrasensitive and Rapid Detection of Viable Listeria Monocytogenes Cells and Biofilms. LWT 2022, 154, 112641. [Google Scholar] [CrossRef]
- Bhatnagar, I.; Mahato, K.; Ealla, K.K.R.; Asthana, A.; Chandra, P. Chitosan Stabilized Gold Nanoparticle Mediated Self-Assembled GliP Nanobiosensor for Diagnosis of Invasive Aspergillosis. Int. J. Biol. Macromol. 2018, 110, 449–456. [Google Scholar] [CrossRef]
- Dhar, B.C.; Reed, A.J.; Mitra, S.; Rodriguez Sanchez, P.; Nedorezova, D.D.; Connelly, R.P.; Rohde, K.H.; Gerasimova, Y.v. Cascade of Deoxyribozymes for the Colorimetric Analysis of Drug Resistance in Mycobacterium Tuberculosis. Biosens. Bioelectron. 2020, 165, 112385. [Google Scholar] [CrossRef] [PubMed]
- Bengtson, H.N.; Homolka, S.; Niemann, S.; Reis, A.J.; da Silva, P.E.; Gerasimova, Y.v.; Kolpashchikov, D.M.; Rohde, K.H. Multiplex Detection of Extensively Drug Resistant Tuberculosis Using Binary Deoxyribozyme Sensors. Biosens. Bioelectron. 2017, 94, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Dallaire, A.M.; Patskovsky, S.; Vallée-Bélisle, A.; Meunier, M. Electrochemical Plasmonic Sensing System for Highly Selective Multiplexed Detection of Biomolecules Based on Redox Nanoswitches. Biosens. Bioelectron. 2015, 71, 75–81. [Google Scholar] [CrossRef]
- Bizid, S.; Blili, S.; Mlika, R.; Haj Said, A.; Korri-Youssoufi, H. Direct Electrochemical DNA Sensor Based on a New Redox Oligomer Modified with Ferrocene and Carboxylic Acid: Application to the Detection of Mycobacterium Tuberculosis Mutant Strain. Anal. Chim. Acta 2017, 994, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tsao, K.C.; Chiou, C.C.; Chen, T.L.; Huang, C.G.; Hsieh, E.F.; Shih, S.R. Detection of Low Copies of Drug-Resistant Influenza Viral Gene by a Single-Tube Reaction Using Peptide Nucleic Acid as Both PCR Clamp and Sensor Probe. J. Microbiol. Immunol. Infect. 2014, 47, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Qi, P.; Zhang, D. Identification of Bacteria by a Fluorescence Sensor Array Based on Three Kinds of Receptors Functionalized Carbon Dots. Sens. Actuators B Chem. 2019, 286, 206–213. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, L.; Xu, Y.; Xue, Y.; Qiao, L.; Ding, C.; Ling, L.; Yu, S. A Platform for Specific and Sensitive Detection of Target Bacteria by Selective Magnetic Enrichment and a Broad-Spectrum Fluorescent Probe. Sens. Actuators B Chem. 2021, 349, 130762. [Google Scholar] [CrossRef]
- Gill, A.A.S.; Singh, S.; Nate, Z.; Chauhan, R.; Thapliyal, N.B.; Karpoormath, R.; Maru, S.M.; Reddy, T.M. A Novel Copper-Based 3D Porous Nanocomposite for Electrochemical Detection and Inactivation of Pathogenic Bacteria. Sens. Actuators B Chem. 2020, 321, 128449. [Google Scholar] [CrossRef]
- Rawat, D.; Nair, D. Extended-Spectrum ß-Lactamases in Gram Negative Bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile β-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC Β-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Thai, H.B.D.; Yu, J.K.; Park, B.S.; Park, Y.J.; Min, S.J.; Ahn, D.R. A Fluorogenic Substrate of Beta-Lactamases and Its Potential as a Probe to Detect the Bacteria Resistant to the Third-Generation Oxyimino-Cephalosporins. Biosens. Bioelectron. 2016, 77, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Torrelles, J.B. Mannose-Capped Lipoarabinomannan in Mycobacterium Tuberculosis Pathogenesis. Pathog. Dis. 2018, 76, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Divagar, M.; Bandaru, R.; Janakiraman, V.; Sai, V.V.R. A plasmonic fiberoptic absorbance biosensor for mannose-capped lipoarabinomannan based tuberculosis diagnosis. Biosens. Bioelectron. 2020, 167, 112488. [Google Scholar] [CrossRef]
- Lüderitz, O.; Freudenberg, M.A.; Galanos, C.; Lehmann, V.; Rietschel, E.T.; Shaw, D.H. Lipopolysaccharides of Gram-Negative Bacteria. Curr. Top. Membr. Transp. 1982, 17, 79–151. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide Endotoxins Endotoxins as Activators of Innate Immunity. Annu. Rev. Biochem. 2008, 71, 635–700. [Google Scholar] [CrossRef]
- Yeo, T.Y.; Choi, J.S.; Lee, B.K.; Kim, B.S.; Yoon, H.I.; Lee, H.Y.; Cho, Y.W. Electrochemical Endotoxin Sensors Based on TLR4/MD-2 Complexes Immobilized on Gold Electrodes. Biosens. Bioelectron. 2011, 28, 139–145. [Google Scholar] [CrossRef]
- Lau, G.W.; Hassett, D.J.; Ran, H.; Kong, F. The Role of Pyocyanin in Pseudomonas Aeruginosa Infection. Trends Mol. Med. 2004, 10, 599–606. [Google Scholar] [CrossRef]
- Fuse, K.; Fujimura, S.; Kikuchi, T.; Gomi, K.; Iida, Y.; Nukiwa, T.; Watanabe, A. Reduction of Virulence Factor Pyocyanin Production in Multidrug-Resistant Pseudomonas Aeruginosa. J. Infect. Chemother. 2013, 19, 82–88. [Google Scholar] [CrossRef]
- Rashid, J.I.A.; Kannan, V.; Ahmad, M.H.; Mon, A.A.; Taufik, S.; Miskon, A.; Ong, K.K.; Yusof, N.A. An Electrochemical Sensor Based on Gold Nanoparticles-Functionalized Reduced Graphene Oxide Screen Printed Electrode for the Detection of Pyocyanin Biomarker in Pseudomonas Aeruginosa Infection. Mater. Sci. Eng. C 2021, 120, 111625. [Google Scholar] [CrossRef]
- Xu, Y.; Dhaouadi, Y.; Stoodley, P.; Ren, D. Sensing the Unreachable: Challenges and Opportunities in Biofilm Detection. Curr. Opin. Biotechnol. 2020, 64, 79–84. [Google Scholar] [CrossRef]
- Kim, Y.W.; Sardari, S.E.; Meyer, M.T.; Iliadis, A.A.; Wu, H.C.; Bentley, W.E.; Ghodssi, R. An ALD Aluminum Oxide Passivated Surface Acoustic Wave Sensor for Early Biofilm Detection. Sens. Actuators B Chem. 2012, 163, 136–145. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Z.; Li, Y.; Xie, G. Amplified Electrochemical Detection of MecA Gene in Methicillin-Resistant Staphylococcus Aureus Based on Target Recycling Amplification and Isothermal Strand-Displacement Polymerization Reaction. Sens. Actuators B Chem. 2015, 221, 148–154. [Google Scholar] [CrossRef]
- Trinh, K.T.L.; Stabler, R.A.; Lee, N.Y. Fabrication of a Foldable All-in-One Point-of-Care Molecular Diagnostic Microdevice for the Facile Identification of Multiple Pathogens. Sens. Actuators B Chem. 2020, 314, 128057. [Google Scholar] [CrossRef]
- Pedrosa, P.; Veigas, B.; Machado, D.; Couto, I.; Viveiros, M.; Baptista, P.v. Gold Nanoprobes for Multi Loci Assessment of Multi-Drug Resistant Tuberculosis. Tuberculosis 2014, 94, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Matsishin, M.; Rachkov, A.; Errachid, A.; Dzyadevych, S.; Soldatkin, A. Development of Impedimetric DNA Biosensor for Selective Detection and Discrimination of Oligonucleotide Sequences of the RpoB Gene of Mycobacterium Tuberculosis. Sens. Actuators B Chem. 2016, 222, 1152–1158. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.; Zhang, L.; Yang, Y.; Yang, S.; Yang, L.; Chen, H.; Liu, C.; Li, J.; Xie, G. Integration of multiplex PCR and CRISPR-Cas allows highly specific detection of multidrug-resistant Acinetobacter baumannii. Sens. Actuators Part B 2021, 334, 129600. [Google Scholar] [CrossRef]
- Hu, C.; Kalsi, S.; Zeimpekis, I.; Sun, K.; Ashburn, P.; Turner, C.; Sutton, J.M.; Morgan, H. Ultra-Fast Electronic Detection of Antimicrobial Resistance Genes Using Isothermal Amplification and Thin Film Transistor Sensors. Biosens. Bioelectron. 2017, 96, 281–287. [Google Scholar] [CrossRef]
- Maldonado, J.; González-Guerrero, A.B.; Fernández-Gavela, A.; González-López, J.J.; Lechuga, L.M. Ultrasensitive label-free detection of unamplified multidrug-resistance bacteria genes with a bimodal waveguide interferometric biosensor. Diagnostics 2020, 10, 845. [Google Scholar] [CrossRef]
- Shin, H.H.; Lee, H.J.; Hwang, M.J.; Kim, J.; Kim, H.; Nam, S.H.; Park, J.S.; Hwang, J.E.; Kim, E.S.; Park, Y.S.; et al. Gold-Silver Core-Shell Nanodumbbells in Solution State as a Highly Sensitive and Reproducible Assay Platform for Bacterial Genome Detection. Sens. Actuators B Chem. 2021, 349, 130784. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Chen, P.; Zhou, X.; Yang, Y.; Wu, S.; Niu, L.; Han, Y.; Wang, L.; Chen, P.; et al. Label-Free, Electrochemical Detection of Methicillin-Resistant Staphylococcus Aureus DNA with Reduced Graphene Oxide-Modified Electrodes. Biosens. Bioelectron. 2011, 26, 3881–3886. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Deng, K.; Xia, H.; Yao, C.; Chen, Q.; Zhang, L.; Liu, Z.; Fu, W. Isothermal Detection of Multiple Point Mutations by a Surface Plasmon Resonance Biosensor with Au Nanoparticles Enhanced Surface-Anchored Rolling Circle Amplification. Biosens. Bioelectron. 2013, 49, 442–449. [Google Scholar] [CrossRef]
- Sun, S.C.; Dou, H.Y.; Chuang, M.C.; Kolpashchikov, D.M. Multi-Labeled Electrochemical Sensor for Cost-Efficient Detection of Single Nucleotide Substitutions in Folded Nucleic Acids. Sens. Actuators B Chem. 2019, 287, 569–575. [Google Scholar] [CrossRef]
- Trinh, T.N.D.; Thai, D.A.; Lee, N.Y. Pop-up Paper-Based and Fully Integrated Microdevice for Point-of-Care Testing of Vancomycin-Resistant Enterococcus. Sens. Actuators B Chem. 2021, 345, 130362. [Google Scholar] [CrossRef]
- Tawil, N.; Mouawad, F.; Lévesque, S.; Sacher, E.; Mandeville, R.; Meunier, M. The Differential Detection of Methicillin-Resistant, Methicillin-Susceptible and Borderline Oxacillin-Resistant Staphylococcus Aureus by Surface Plasmon Resonance. Biosens. Bioelectron. 2013, 49, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, Q.; Wu, Z.; Lu, Z. Detection of hlyA Gene of Listeria Monocytogenes with Electrochemical DNA Biosensor. In Proceedings of the 2nd International Conference on Bioinformatics and Biomedical Engineering, Shanghai, China, 16–18 May 2008; pp. 375–378. [Google Scholar] [CrossRef]
- Shin, Y.; Perera, A.P.; Tang, W.Y.; Fu, D.L.; Liu, Q.; Sheng, J.K.; Gu, Z.; Lee, T.Y.; Barkham, T.; Kyoung Park, M. A Rapid Amplification/Detection Assay for Analysis of Mycobacterium Tuberculosis Using an Isothermal and Silicon Bio-Photonic Sensor Complex. Biosens. Bioelectron. 2015, 68, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wei, L.; Chen, L. Application of Metal-Organic Framework (MOF)-Based Enzymatic Amplification Strategy for the Sensitive Electrochemical Detection of Tuberculosis. Sens. Actuators B Chem. 2020, 324, 128724. [Google Scholar] [CrossRef]
- Tiwari, I.; Singh, M.; Pandey, C.M.; Sumana, G. RSC Advances Electrochemical Detection of a Pathogenic Escherichia Coli Speci Fi c DNA Sequence Based on a Graphene Oxide—Chitosan Composite Decorated with Nickel Ferrite Nanoparticles. RSC Adv. 2015, 5, 67115–67124. [Google Scholar] [CrossRef]
- Wang, J.C.; Tung, Y.C.; Ichiki, K.; Sakamoto, H.; Yang, T.H.; ichiro Suye, S.; Chuang, H.S. Culture-Free Detection of Methicillin-Resistant Staphylococcus Aureus by Using Self-Driving Diffusometric DNA Nanosensors. Biosens. Bioelectron. 2020, 148, 111817. [Google Scholar] [CrossRef]
- Ahangar, L.E.; Mehrgardi, M.A. Amplified Detection of Hepatitis B Virus Using an Electrochemical DNA Biosensor on a Nanoporous Gold Platform. Bioelectrochemistry 2017, 117, 83–88. [Google Scholar] [CrossRef]
- Chandra, S.; Mahto, T.K.; Chowdhuri, A.R.; Das, B.; Sahu, S. kumar One Step Synthesis of Functionalized Carbon Dots for the Ultrasensitive Detection of Escherichia Coli and Iron (III). Sens. Actuators B Chem. 2017, 245, 835–844. [Google Scholar] [CrossRef]
- Safardoust-Hojaghan, H.; Salavati-Niasari, M.; Amiri, O.; Hassanpour, M. Preparation of Highly Luminescent Nitrogen Doped Graphene Quantum Dots and Their Application as a Probe for Detection of Staphylococcus Aureus and E. Coli. J. Mol. Liq 2017, 241, 1114–1119. [Google Scholar] [CrossRef]
- Wang, N.; Wang, Y.; Guo, T.; Yang, T.; Chen, M.; Wang, J. Green Preparation of Carbon Dots with Papaya as Carbon Source for Effective Fluorescent Sensing of Iron (III) and Escherichia Coli. Biosens. Bioelectron. 2016, 85, 68–75. [Google Scholar] [CrossRef]
- Chen, K.H.; Lee, S.H.; Kok, L.C.; Ishdorj, T.O.; Chang, H.Y.; Tseng, F.G. A 3D-ACEK/SERS System for Highly Efficient and Selectable Electrokinetic Bacteria Concentration/Detection/ Antibiotic-Susceptibility-Test on Whole Blood. Biosens. Bioelectron. 2022, 197, 113740. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zheng, Y.; Liu, Y.; Feng, X.; Dong, Q.; Li, J.; Wang, J.; Zhao, C. A Concise Detection Strategy of Staphylococcus Aureus Using N-Succinyl-Chitosan-Dopped Bacteria-Imprinted Composite Film and AIE Fluorescence Sensor. J. Hazard. Mater. 2022, 423, 126934. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, S.D.; Bhosle, A.A.; Nayse, A.; Biswas, S.; Biswas, M.; Bhasikuttan, A.C.; Banerjee, M.; Chatterjee, A. A Redox-Coupled Carbon Dots-MnO2 Nanosheets Based Sensory Platform for Label-Free and Sensitive Detection of E. Coli. Sens. Actuators B Chem. 2021, 339, 129918. [Google Scholar] [CrossRef]
- Wang, H.Y.; Chen, C.Y.; Chu, P.Y.; Zhu, Y.X.; Hsieh, C.H.; Lu, J.J.; Wu, M.H. Application of an Optically Induced Dielectrophoresis (ODEP)-Based Microfluidic System for the Detection and Isolation of Bacteria with Heterogeneity of Antibiotic Susceptibility. Sens. Actuators B Chem. 2020, 307, 127540. [Google Scholar] [CrossRef]
- Xia, H.; Huang, J.; Lu, X.; Wang, G.; Zhang, Z.; Yue, J.; Li, Q.; Wang, S.; Yan, J.; Deng, L.; et al. Autocatalytic MNAzyme-Integrated Surface Plasmon Resonance Biosensor for Simultaneous Detection of Bacteria from Nosocomial Bloodstream Infection Specimens. Sens. Actuators B Chem. 2021, 330, 129255. [Google Scholar] [CrossRef]
- Wang, H.; Chi, Z.; Cong, Y.; Wang, Z.; Jiang, F.; Geng, J.; Zhang, P.; Ju, P.; Dong, Q.; Liu, C. Development of a Fluorescence Assay for Highly Sensitive Detection of: Pseudomonas Aeruginosa Based on an Aptamer-Carbon Dots/Graphene Oxide System. RSC Adv. 2018, 8, 32454–32460. [Google Scholar] [CrossRef]
- Villamizar, R.A.; Maroto, A.; Rius, F.X.; Inza, I.; Figueras, M.J. Fast Detection of Salmonella Infantis with Carbon Nanotube Field Effect Transistors. Biosens. Bioelectron. 2008, 24, 279–283. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Jiang, H. Label-Free Detection of Acinetobacter Baumannii through the Induced Fluorescence Quenching of Thiolated AuAg Nanoclusters. Sens. Actuators B Chem. 2018, 277, 388–393. [Google Scholar] [CrossRef]
- Kumar, N.; Wang, W.; Ortiz-Marquez, J.C.; Catalano, M.; Gray, M.; Biglari, N.; Hikari, K.; Ling, X.; Gao, J.; van Opijnen, T.; et al. Dielectrophoresis Assisted Rapid, Selective and Single Cell Detection of Antibiotic Resistant Bacteria with G-FETs. Biosens. Bioelectron. 2020, 156, 112123. [Google Scholar] [CrossRef]
- Quero, G.; Zuppolini, S.; Consales, M.; Diodato, L.; Vaiano, P.; Venturelli, A.; Santucci, M.; Spyrakis, F.; Costi, M.P.; Giordano, M.; et al. Long Period Fiber Grating Working in Reflection Mode as Valuable Biosensing Platform for the Detection of Drug Resistant Bacteria. Sens. Actuators B Chem. 2016, 230, 510–520. [Google Scholar] [CrossRef]
- Atchudan, R.; Edison, T.N.J.I.; Chakradhar, D.; Perumal, S.; Shim, J.J.; Lee, Y.R. Facile Green Synthesis of Nitrogen-Doped Carbon Dots Using Chionanthus Retusus Fruit Extract and Investigation of Their Suitability for Metal Ion Sensing and Biological Applications. Sens. Actuators B Chem. 2017, 246, 497–509. [Google Scholar] [CrossRef]
- Sypabekova, M.; Jolly, P.; Estrela, P.; Kanayeva, D. Electrochemical Aptasensor Using Optimized Surface Chemistry for the Detection of Mycobacterium Tuberculosis Secreted Protein MPT64 in Human Serum. Biosens. Bioelectron. 2019, 123, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Z.; Yu, H.W.; Wang, N.; Zhang, Z.; Wan, G.C.; Liu, H.; Guan, X.; Chang, D. Electrochemical DNA Biosensor Based on a Glassy Carbon Electrode Modified with Gold Nanoparticles and Graphene for Sensitive Determination of Klebsiella Pneumoniae Carbapenemase. J. Biotechnol 2015, 214, 133–138. [Google Scholar] [CrossRef]
- Hilton, S.H.; Hall, C.; Nguyen, H.T.; Everitt, M.L.; DeShong, P.; White, I.M. Phenotypically Distinguishing ESBL-Producing Pathogens Using Paper-Based Surface Enhanced Raman Sensors. Anal. Chim. Acta 2020, 1127, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Andersson, T.; Bläckberg, A.; Lood, R.; Bergdahl, G.E. Development of a Molecular Imprinting-Based Surface Plasmon Resonance Biosensor for Rapid and Sensitive Detection of Staphylococcus Aureus Alpha Hemolysin From Human Serum. Front. Cell. Infect. Microbiol. 2020, 10, 571578. [Google Scholar] [CrossRef]
- Poma, N.; Vivaldi, F.; Bonini, A.; Salvo, P.; Kirchhain, A.; Melai, B.; Bottai, D.; Tavanti, A.; di Francesco, F. A Graphenic and Potentiometric Sensor for Monitoring the Growth of Bacterial Biofilms. Sens. Actuators B Chem. 2020, 323, 128662. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Brozovic, A.; Gonçalves, A.C.; Jurkovicova, D.; Linē, A.; Machuqueiro, M.; Saponara, S.; Sarmento-Ribeiro, A.B.; Xavier, C.P.R.; Vasconcelos, M.H. The Multi-Factorial Nature of Clinical Multidrug Resistance in Cancer. Drug Resist. Updat. 2019, 46, 100645. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Feng, S.S.; Guo, Y. Nanomedicine against Multidrug Resistance in Cancer Treatment. Nanomedicine 2012, 7, 465–468. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. In Cancer Drug Resistance; OAE Publishing, Inc.: Alhambra, CA, USA, 2019; Volume 2, pp. 141–160. [Google Scholar] [CrossRef] [Green Version]
- Szakács, G.; Hall, M.D.; Gottesman, M.M.; Boumendjel, A.; Kachadourian, R.; Day, B.J.; Baubichon-Cortay, H.; di Pietro, A. Targeting the Achilles Heel of Multidrug-Resistant Cancer by Exploiting the Fitness Cost of Resistance. Chem Rev. 2014, 114, 5753–5774. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The different mechanisms of cancer drug resistance: A brief review. In Advanced Pharmaceutical Bulletin; Tabriz University of Medical Sciences: Tabriz, Iran, 2017; Volume 7, pp. 339–348. [Google Scholar] [CrossRef]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of multidrug resistance in cancer chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef] [PubMed]
- van Veen, H.W.; Konings, W.N. The ABC Family of Multidrug Transporters in Microorganisms. Biochim. Biophys. Acta Bioenerg. 1998, 1365, 31–36. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. In Nature Reviews. Cancer; Nature Publishing Group: Berlin, Germany, 2018; Volume 18, pp. 452–464. [Google Scholar] [CrossRef]
- Gillet, J.P.; Gottesman, M.M. Mechanisms of Multidrug Resistance in Cancer. Methods Mol. Biol. 2010, 596, 47–76. [Google Scholar] [CrossRef]
- Kimchi-Sarfaty, C.; Marple, A.H.; Shinar, S.; Kimchi, A.M.; Scavo, D.; Roma, M.I.; Kim, I.W.; Jones, A.; Arora, M.; Gribar, J.; et al. Ethnicity-related polymorphisms and haplotypes in the human ABCB1 gene. Pharmacogenomics 2007, 8, 29–39. [Google Scholar] [CrossRef]
- Wen-Hsuan, C.; Cerione Richard, A.; Antonyak Mark, A. Cancer cell signaling methods and protocols third edition. In Cancer Cell Signaling; Springer: Berlin, Germany, 2021; Available online: springer.com/book/10.1007/978-1-0716-0759-6 (accessed on 26 January 2022).
- Momen-Heravi, F.; Bala, S. The miRNA and Extracellular Vesicles in Alcoholic Liver Disease; Elsevier, Inc.: Amsterdam, The Netherlands, 2016; Volume 2000, ISBN 9780128007730. [Google Scholar] [CrossRef]
- Paoli, P.; Giannoni, E.; Chiarugi, P. Anoikis Molecular Pathways and Its Role in Cancer Progression. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 3481–3498. [Google Scholar] [CrossRef]
- Arneth, B. Tumor microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Bae, J.S.; Tam, M. Tumor-associated macrophages and neutrophils in tumor microenvironment. Mediat. Inflamm. 2016, 2016, 6058147. [Google Scholar] [CrossRef]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer stem cells (CSCs) in drug resistance and their therapeutic implications in cancer treatment. In Stem Cells International; Hindawi Publishing Limited: London, UK, 2018; Volume 2018, p. 5416923. [Google Scholar] [CrossRef]
- Robles-Flores. Cancer Cell Signaling Methods and Protocols Third Edition; Fighting Cancer Resistance. 2021. Available online: www.springer.com/series/7651 (accessed on 20 July 2022).
- Ricardo, P. Lactate in the Tumor Microenvironment: An Essential; MDPI: Basel, Switzerland, 2020; pp. 1–29. [Google Scholar]
- Pérez-Tomás, R.; Pérez-Guillén, I. Lactate in the tumor microenvironment: An essential molecule in cancer progression and treatment. Cancers 2020, 12, 3244. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.P.; Hu, Y.; Liu, P.; Deng, Y.N.; Wang, P.; Chen, W.; Liu, A.L.; Chen, Y.Z.; Lin, X.H. Label-Free Electrochemical DNA Biosensor for Rapid Detection of Mutidrug Resistance Gene Based on Au Nanoparticles/Toluidine Blue-Graphene Oxide Nanocomposites. Sens. Actuators B Chem. 2015, 207, 269–276. [Google Scholar] [CrossRef]
- Chen, M.; Hou, C.; Huo, D.; Bao, J.; Fa, H.; Shen, C. An Electrochemical DNA Biosensor Based on Nitrogen-Doped Graphene/Au Nanoparticles for Human Multidrug Resistance Gene Detection. Biosens. Bioelectron. 2016, 85, 684–691. [Google Scholar] [CrossRef]
- Prattis, I.; Hui, E.; Gubeljak, P.; Kaminski Schierle, G.S.; Lombardo, A.; Luigi, G. Occhipinti, Graphene for Biosensing Applications in Point-of-Care Testing. Trends Biotechnol. 2021, 39, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Zou, B.; Zhang, L.; Ma, X.; Qi, X.; Wei, W.; Song, Q.; Zhou, G. Ultra-Sensitive and Multiplex Digital-PCR for Quantifying the Mutants in Cell Free DNA by Employing Invasive Reaction as Identifier. Sens. Actuators B Chem. 2020, 320, 128362. [Google Scholar] [CrossRef]
- Ma, C.; Wei, S.; Song, Y. T790M and Acquired Resistance of EGFR TKI: A Literature Review of Clinical Reports; Journal for Thoracic Diseases: Hong Kong, China, 2010; Volume 3. [Google Scholar] [CrossRef]
- Brien, J.O.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, function and role in cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
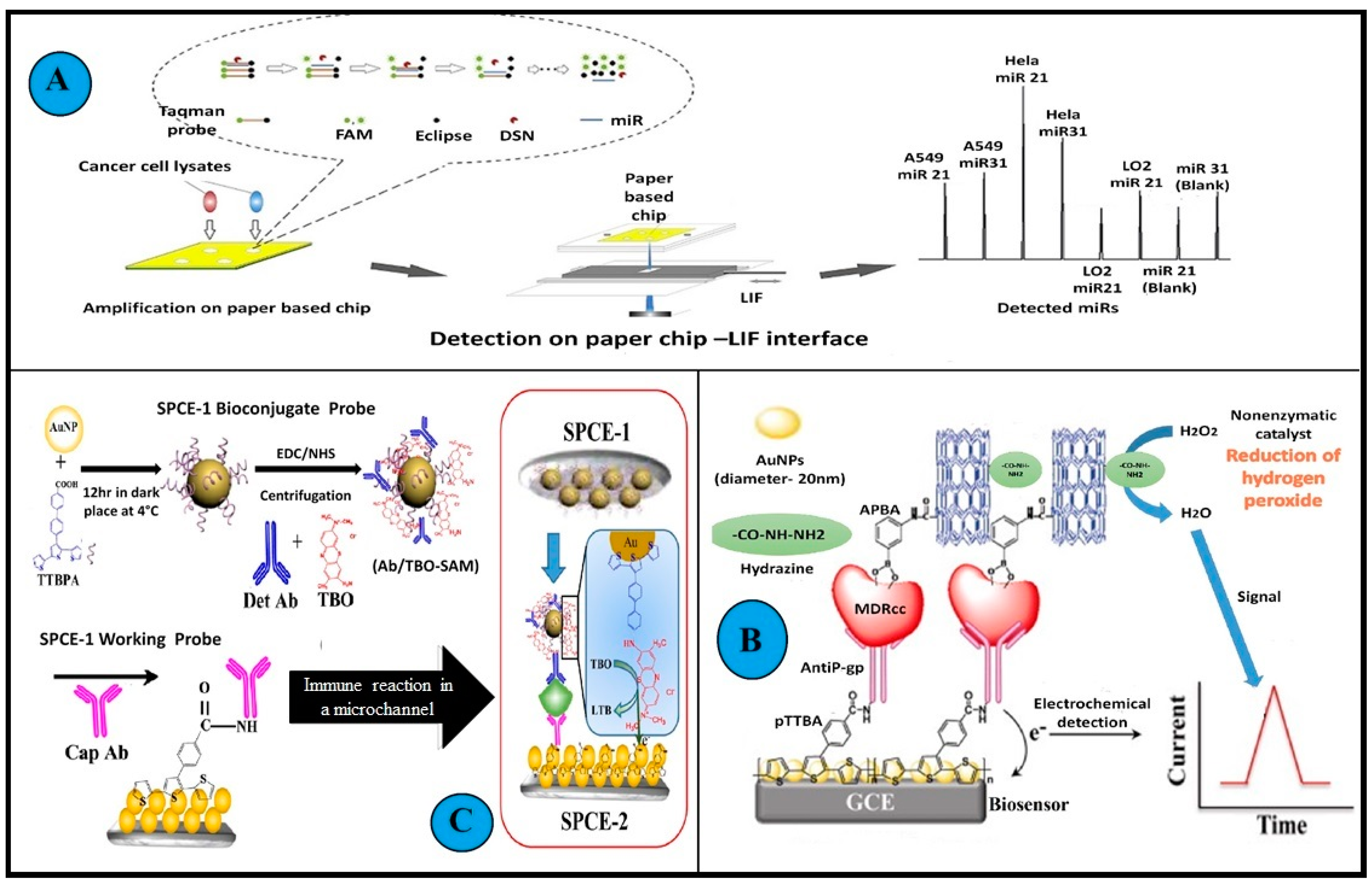
- Cai, X.; Zhang, H.; Yu, X.; Wang, W. A Microfluidic Paper-Based Laser-Induced Fluorescence Sensor Based on Duplex-Specific Nuclease Amplification for Selective and Sensitive Detection of MiRNAs in Cancer Cells. Talanta 2020, 216, 120996. [Google Scholar] [CrossRef] [PubMed]
- Yaman, Y.T.; Vural, O.A.; Bolat, G.; Abaci, S. One-Pot Synthesized Gold Nanoparticle-Peptide Nanotube Modified Disposable Sensor for Impedimetric Recognition of MiRNA 410. Sens. Actuators B Chem. 2020, 320, 128343. [Google Scholar] [CrossRef]
- Qu, H.; Fan, C.; Chen, M.; Zhang, X.; Yan, Q.; Wang, Y.; Zhang, S.; Gong, Z.; Shi, L.; Li, X.; et al. Recent Advances of Fluorescent Biosensors Based on Cyclic Signal Amplification Technology in Biomedical Detection. J. Nanobiotechnol. 2021, 19, 403. [Google Scholar] [CrossRef]
- Dong, Z.; Ren, L.; Lin, L.; Li, J.; Huang, Y.; Li, J. Effect of MicroRNA-21 on Multidrug Resistance Reversal in A549/DDP Human Lung Cancer Cells. Mol. Med. Rep. 2015, 11, 682–690. [Google Scholar] [CrossRef]
- Feng, Y.H.; Tsao, C.J. Emerging Role of MicroRNA-21 in Cancer (Review). Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef]
- National Library of Medicine MIR155 MicroRNA 155 [Homo Sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/406947#summary (accessed on 26 July 2022).
- Bayraktar, R.; van Roosbroeck, K. miR-155 in cancer drug resistance and as target for miRNA-based therapeutics. Cancer Metastasis Rev. 2018, 37, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Qiu, Y.; Yagüe, E.; Ji, W.; Liu, J.; Zhang, J. MiRNA-205 Targets VEGFA and FGF2 and Regulates Resistance to Chemotherapeutics in Breast Cancer. Cell Death Dis. 2016, 7, e2291. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.L. Liver-specific microRNA-122. RNA Biol. 2012, 9, 137–142. [Google Scholar] [CrossRef]
- Yahya, S.M.M.; Fathy, S.A. Possible Role of MicroRNA-122 in Modulating Multidrug Resistance of Hepatocellular Carcinoma. Indian J. Clin. Biochem. 2018, 33, 21–30. [Google Scholar] [CrossRef]
- Dong, J.; Liu, Y.; Liao, W.; Liu, R.; Shi, P.; Wang, L. MiRNA-223 Is a Potential Diagnostic and Prognostic Marker for Osteosarcoma. J. Bone Oncol. 2016, 5, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, J.; Zhang, S.; Zheng, X.; Xie, S.; Mao, J.; Cai, Y.; Lu, X.; Hu, L.; Shen, J.; et al. MiR—223 regulates autophagy associated with cisplatin resistance by targeting FBXW7 in human non—Small cell lung cancer. Cancer Cell Int. 2020, 20, 258. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zheng, Z.M.; Li, X.N.; Li, Z.F.; Wang, Y.; Geng, Y.F.; Bai, L.; Zhang, X.B. MiR-223 modulates multidrug resistance via downregulation of ABCB1 in hepatocellular carcinoma cells. Exp. Biol. Med. 2013, 238, 1024–1032. [Google Scholar] [CrossRef]
- Stepicheva, N.A.; Song, J.I.A.L. Function and Regulation of MicroRNA-31 in Development and Disease. Mol. Reprod. Dev. 2018, 83, 654–674. [Google Scholar] [CrossRef]
- Humphries, B.; Yang, C. The microRNA-200 family: Small molecules with novel roles in cancer development, progression and therapy. Oncotarget 2015, 6, 6472–6498. [Google Scholar] [CrossRef]
- Lee, H.; Kim, C.; Kang, H.; Tak, H.; Ahn, S.; Yoon, S.K.; Kuh, H.J.; Kim, W.; Lee, E.K. microRNA-200a-3p increases 5- fluorouracil resistance by regulating dual speci fi city phosphatase 6 expression. Exp. Mol. Med. 2017, 49, e327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 family: A potential tumor suppressor and therapeutic candidate in cancer. J. Exp. Clin. Cancer Res. 2019, 5, 53. [Google Scholar] [CrossRef]
- Physiology, C. MicroRNA-34a Attenuates Metastasis and Chemoresistance of Bladder Cancer Cells by Targeting the TCF1/LEF1 Axis. Cell. Physiol. Biochem. 2018, 400037, 87–98. [Google Scholar] [CrossRef]
- Schneider, J.W.; Dittmer, D.P. Diagnosis and Treatment of Kaposi Sarcoma Johann. Am. J. Clin. Dermatol. 2018, 18, 529–539. [Google Scholar] [CrossRef]
- Zhang, J.; Pu, X.; Xiong, Y. kshv-mir-k12-1-5p Promotes Cell Growth and Metastasis by Targeting SOCS6 in Kaposi’s Sarcoma Cells; Dove Medical Press: Macclesfield, UK, 2019; pp. 4985–4995. [Google Scholar]
- Rta, O.R.F.; Lin, X.; Liang, D.; He, Z.; Deng, Q.; Robertson, E.S.; Lan, K. MiR-K12-7-5p Encoded by Kaposi’ s Sarcoma-Associated Herpesvirus Stabilizes the Latent State by Targeting Viral. PLoS ONE 2011, 6, e0016224. [Google Scholar] [CrossRef]
- Wang, K. Role of microRNA—410 in molecular oncology: A double edged sword. J. Cell. Biochem. 2018, 119, 8737–8742. [Google Scholar] [CrossRef]
- Chen, R.E.N.; Li, X.; He, B.I.N.; Hu, W.E.I. MicroRNA-410 Regulates Autophagy-Related Gene ATG16L1 Expression and Enhances Chemosensitivity via Autophagy Inhibition in Osteosarcoma. Mol. Med. Rep. 2017, 15, 1326–1334. [Google Scholar] [CrossRef]
- Milevskiy, M.J.G.; Gujral, U.; Del, C.; Marques, L.; Stone, A.; Northwood, K.; Burke, L.J.; Gee, J.M.W.; Nephew, K.; Clark, S.; et al. MicroRNA-196a Is Regulated by ER and Is a Prognostic Biomarker in ER + Breast Cancer. Br. J. Cancer 2019, 120, 621–632. [Google Scholar] [CrossRef]
- Xin, H.; Wang, C.; Liu, Z. miR-196a-5p promotes metastasis of colorectal cancer via targeting I κ B α. BMC Cancer 2019, 19, 30. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, B.; Han, S.; Zhang, K.; Chen, J.; Li, C.; Wang, R. The Roles of MicroRNA-141 in Human Cancers: From Diagnosis to Treatment. Cell. Physiol. Biochem. 2016, 38, 427–448. [Google Scholar] [CrossRef]
- Kinose, Y.; Sawada, K.; Nakamura, K.; Kimura, T. The role of microRNAs in ovarian cancer. BioMed Res. Int. 2014, 2014, 249393. [Google Scholar] [CrossRef] [PubMed]
- Su, J.L.; Chen, P.S.; Johansson, G.; Kuo, M.L. Function and regulation of Let-7 family microRNAs. MicroRNA 2012, 1, 34–39. [Google Scholar] [CrossRef]
- Kolenda, T.; Przybyła, W.; Teresiak, A. The Mystery of Let-7d—A Small RNA with Great Power. Contemp. Oncol./Współczesna Onkol. 2014, 18, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Yang, W.; Feng, W.; Cao, L.; Wang, X.; Niu, L.; Li, Y.; Zhou, W.; Zhang, Y.; Liu, J.; et al. Molecular Mechanisms and Clinical Implications of MiRNAs in Drug Resistance of Colorectal Cancer. Ther. Adv. Med. Oncol. 2020, 12, 1758835920947342. [Google Scholar] [CrossRef]
- Garg, H.; Suri, P.; Gupta, J.C.; Talwar, G.P.; Dubey, S. Survivin: A Unique Target for Tumor Therapy. Cancer Cell Int. 2016, 16, 49. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.; Gui-mei, Z.; Zuo-Hua, F. Down-Regulation of Survivin Expression Reversed Multidrug Resistance in Adriamycin- Resistant HL-60/ADR Cell Line 1. Acta Pharmacol. Sin. 2003, 24, 1235–1240. [Google Scholar]
- Hu, H.; Jiang, H.; Wang, X.; Chen, B. Detection and Distinguishability of Leukemia Cancer Cells Based on Au Nanoparticles Modified Electrodes. Electrochem. Commun. 2008, 10, 1121–1124. [Google Scholar] [CrossRef]
- Tao, Y.; Li, M.; Auguste, D.T. Pattern-Based Sensing of Triple Negative Breast Cancer Cells with Dual-Ligand Cofunctionalized Gold Nanoclusters. Biomaterials 2017, 116, 21–33. [Google Scholar] [CrossRef]
- Chandra, P.; Noh, H.B.; Pallela, R.; Shim, Y.B. Ultrasensitive Detection of Drug Resistant Cancer Cells in Biological Matrixes Using an Amperometric Nanobiosensor. Biosens. Bioelectron. 2015, 70, 418–425. [Google Scholar] [CrossRef]
- Gulati, P.; Kaur, P.; Rajam, M.v.; Srivastava, T.; Ali, M.A.; Mishra, P.; Islam, S.S. Leukemia Biomarker Detection by Using Photoconductive Response of CNT Electrode: Analysis of Sensing Mechanism Based on Charge Transfer Induced Fermi Level Fluctuation. Sens. Actuators B Chem. 2018, 270, 45–55. [Google Scholar] [CrossRef]
- Li, Y.; Chen, D.; Zhang, Y.; Liu, C.; Chen, P.; Wang, Y.; Feng, X.; Du, W.; Liu, B.F. High-Throughput Single Cell Multidrug Resistance Analysis with Multifunctional Gradients-Customizing Microfluidic Device. Sens. Actuators B Chem. 2016, 225, 563–571. [Google Scholar] [CrossRef]
- Berrazouane, S.; Boisvert, M.; Salti, S.; Mourad, W.; Al-Daccak, R.; Barabé, F.; Aoudjit, F. Beta1 integrin blockade overcomes doxorubicin resistance in human T-cell acute lymphoblastic leukemia. Cell Death Dis. 2019, 10, 357. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Guo, J.; Liao, Q.; Zhao, Y. β 1 and β 3 Integrins in Breast, Prostate and Pancreatic Cancer: A Novel Implication (Review). Oncol. Lett. 2018, 15, 5412–5416. [Google Scholar] [CrossRef] [PubMed]
- Morozevich, G.E.; Kozlova, N.I.; Preobrazhenskaya, M.E.; Ushakova, N.A.; Eltsov, I.A.; Shtil, A.A.; Berman, A.E. The role of β 1 integrin subfamily in anchorage-dependent apoptosis of breast carcinoma cells differing in multidrug resistance. Biochem. Biokhimiia. 2006, 71, 489–495. [Google Scholar] [CrossRef]
- Jiang, X.; Tan, L.; Zhang, B.; Zhang, Y.; Tang, H.; Xie, Q.; Yao, S. Detection of Adherent Cells Using Electrochemical Impedance Spectroscopy Based on Molecular Recognition of Integrin Β1. Sens. Actuators B Chem. 2010, 149, 87–93. [Google Scholar] [CrossRef]
- Odening, K.E.; Li, W.; Rutz, R. Enhanced complement resistance in drug-selected P-glycoprotein expressing multi-drug-resistant ovarian carcinoma cells. Clin. Exp. Immunol. 2008, 155, 239–248. [Google Scholar] [CrossRef]
- Choudhary, M.; Yadav, P.; Singh, A.; Kaur, S.; Ramirez-Vick, J. CD 59 Targeted Ultrasensitive Electrochemical Immunosensor for Fast and Noninvasive Diagnosis of Oral Cancer. Electroanalysis 2016, 28, 2565–2574. [Google Scholar] [CrossRef]
- Meng, L.; Liu, B.; Ji, R.U.I.; Jiang, X.I.N.; Yan, X.; Xin, Y. Targeting the BDNF / TrkB Pathway for the Treatment of Tumors. Oncol. Lett. 2019, 17, 2031–2039. [Google Scholar] [CrossRef]
- Akhtar, M.H.; Hussain, K.K.; Gurudatt, N.G.; Chandra, P.; Shim, Y. Ultrasensitive Dual Probe Immunosensor for the Monitoring of Nicotine Induced-Brain Derived Neurotrophic Factor Released from Cancer Cells. Biosens. Bioelectron. 2018, 116, 108–115. [Google Scholar] [CrossRef]
- Wang, X.; Yang, D.; Liu, M.; Cao, D.; He, N.; Wang, Z. Highly Sensitive Fluorescence Biosensor for Intracellular Telomerase Detection Based on a Single Patchy Gold/Carbon Nanosphere via the Combination of Nanoflare and Hybridization Chain Reaction. Biosens. Bioelectron. 2019, 137, 110–116. [Google Scholar] [CrossRef]
- Lipinska, N.; Romaniuk, A.; Paszel-Jaworska, A.; Toton, E.; Kopczynski, P.; Rubis, B. Telomerase and drug resistance in cancer. Cell. Mol. Life Sci. 2017, 74, 4121–4132. [Google Scholar] [CrossRef]
- Gao, P.; Yang, X.; Xue, Y.W.; Zhang, X.F.; Wang, Y.; Liu, W.J.; Wu, X.J. Promoter methylation of glutathione S-transferase p 1 and multidrug resistance gene 1 in bronchioloalveolar carcinoma and its correlation with DNA methyltransferase 1 expression. Cancer 2009, 115, 3222–3232. [Google Scholar] [CrossRef] [PubMed]
- Sodani, K.; Patel, A.; Kathawala, R.J.; Chen, Z.S. Multidrug Resistance Associated Proteins in Multindrung Resistance. Chin. J. Cancer 2012, 31, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.K.; El-osta, A. The Rise of DNA Methylation and the Importance of Chromatin on Multidrug Resistance in Cancer. Exp. Cell Res. 2003, 290, 177–194. [Google Scholar] [CrossRef]
- Jones, P.M.; George, A.M. Multidrug Resistance in Parasites: ABC Transporters, P-Glycoproteins and Molecular Modelling. Int. J. Parasitol 2005, 35, 555–566. [Google Scholar] [CrossRef]
- Yang, P.; Zheng, Y.; Wang, Y.; Yue, J.; Gao, G.; Ma, Y.; Tang, B. A novel sensitive NMOF fluorescent probe for two photon imaging of glutathione in chemo-resistant cancer cells. Sens. Actuators Part B 2021, 348, 130680. [Google Scholar] [CrossRef]
- Fattahi, S.; Golpour, M.; Amjadi-Moheb, F.; Sharifi-Pasandi, M.; Khodadadi, P.; Pilehchian-Langroudi, M.; Ashrafi, G.H.; Akhavan-Niaki, H. DNA Methyltransferases and Gastric Cancer: Insight into Targeted Therapy. Epigenomics 2018, 10, 1477–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Liu, F.; Fan, T.; Ren, Y.; Jiang, Y. Rapid Detection of Methyltransferases Utilizing Dumbbell DNA-Templated Copper Nanoparticles. Sens. Actuators B Chem. 2018, 276, 499–506. [Google Scholar] [CrossRef]
- Zastawna, M.; Colas, C.; Hiebel, M.; Suzenet, F. Nitro-Pyrazinotriazapentalene Scaffolds—Nitroreductase Quantification and In Vitro Fluorescence Imaging of Hypoxia; Sensors and Actuators: Amsterdam, The Netherlands, 2021; Volume 346. [Google Scholar]
- Wang, Y.; Zhang, L.; Huang, Y.; Wang, X.; Zhang, L.; Chen, L. Rational Design of a Nitroreductase-Activatable Two-Photon Fluorescent Probe for Hypoxia Imaging in Cell and in Vivo. Sens. Actuators B Chem. 2020, 310, 127755. [Google Scholar] [CrossRef]
- Ge, Y.; Wei, P.; Wang, T.; Cao, X.; Zhang, D.; Li, F. A Simple Fluorescent Probe for Monitoring PH in Cellsbased on New Fluorophorepyrido [1,2-a] Benzimidazole. Sens. Actuators B Chem. 2018, 254, 314–320. [Google Scholar] [CrossRef]
- Zhao, S.S.; Bukar, N.; Toulouse, J.L.; Pelechacz, D.; Robitaille, R.; Pelletier, J.N.; Masson, J.F. Miniature Multi-Channel SPR Instrument for Methotrexate Monitoring in Clinical Samples. Biosens. Bioelectron. 2015, 64, 664–670. [Google Scholar] [CrossRef]
- Hunter, R.A.; Asare-Werehene, M.; Mandour, A.; Tsang, B.K.; Anis, H. Determination of Chemoresistance in Ovarian Cancer by Simultaneous Quantification of Exosomes and Exosomal Cisplatin with Surface Enhanced Raman Scattering. Sens. Actuators B Chem. 2021, 354, 131237. [Google Scholar] [CrossRef]
- Kalyane, D.; Choudhary, D.; Polaka, S.; Goykar, H.; Anup, N.; Tambe, V.; Kalia, K.; Tekade, R.K. Exosomes in Multidrug-Resistant Cancer. Curr. Opin. Pharm. 2020, 54, 109–120. [Google Scholar] [CrossRef]
- Tai, Y.L.; Chen, K.C.; Hsieh, J.T.; Shen, T.L. Exosomes in Cancer Development and Clinical Applications. Cancer Sci. 2018, 109, 2364–2374. [Google Scholar] [CrossRef] [PubMed]
- Zada, S.; Lu, H.; Dai, W.; Tang, S.; Khan, S.; Yang, F.; Qiao, Y.; Fu, P.; Dong, H.; Zhang, X. Multiple Amplified MicroRNAs Monitoring in Living Cells Based on Fluorescence Quenching of Mo2B and Hybridization Chain Reaction. Biosens. Bioelectron. 2022, 197, 113815. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, M.; Xiong, X.; Liu, Y.; Chen, X. Simultaneous Fluorescent Detection of Multiplexed MiRNA of Liver Cancer Based on DNA Tetrahedron Nanotags. Talanta 2020, 210, 120677. [Google Scholar] [CrossRef]
- Peng, W.; Huang, Y.; Zhao, Q.; Lian, G.; Chen, M.; Piao, J.; Gong, X.; Chang, J. A Fluorescent Signal “Removal” Sensor via Duplex-Specific Nuclease-Aided Cleavage for MiRNA Detection in Flow Cytometry. Colloids Surf. B Biointerfaces 2020, 185, 110570. [Google Scholar] [CrossRef]
- Na, H.K.; Wi, J.S.; Son, H.Y.; Ok, J.G.; Huh, Y.M.; Lee, T.G. Discrimination of Single Nucleotide Mismatches Using a Scalable, Flexible, and Transparent Three-Dimensional Nanostructure-Based Plasmonic MiRNA Sensor with High Sensitivity. Biosens. Bioelectron. 2018, 113, 39–45. [Google Scholar] [CrossRef]
- Wang, B.; Chen, Z.; Ren, D.; You, Z. A Novel Dual Energy Transfer Probe for Intracellular MRNA Detection with High Robustness and Specificity. Sens. Actuators B Chem. 2019, 279, 342–350. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, S.; Zhang, T.; Zheng, B.; Tang, H. Bioinspired Sensor Chip for Detection of MiRNA-21 Based on Photonic Crystals Assisted Cyclic Enzymatic Amplification Method. Biosens. Bioelectron. 2020, 150, 111866. [Google Scholar] [CrossRef]
- Isin, D.; Eksin, E.; Erdem, A. Graphene Oxide Modified Single-Use Electrodes and Their Application for Voltammetric MiRNA Analysis. Mater. Sci. Eng. C 2017, 75, 1242–1249. [Google Scholar] [CrossRef]
- Bahari, D.; Babamiri, B.; Salimi, A.; Rashidi, A. Graphdiyne/Graphene Quantum Dots for Development of FRET Ratiometric Fluorescent Assay toward Sensitive Detection of MiRNA in Human Serum and Bioimaging of Living Cancer Cells. J. Lumin 2021, 239, 118371. [Google Scholar] [CrossRef]
- Han, Y.; Qiang, L.; Gao, Y.; Gao, J.; He, Q.; Liu, H.; Han, L.; Zhang, Y. Large-Area Surface-Enhanced Raman Spectroscopy Substrate by Hybrid Porous GaN with Au/Ag for Breast Cancer MiRNA Detection. Appl Surf. Sci. 2021, 541, 148456. [Google Scholar] [CrossRef]
- Mao, Y.; Sun, Y.; Xue, J.; Lu, W.; Cao, X. Ultra-Sensitive and High Efficiency Detection of Multiple Non-Small Cell Lung Cancer-Related MiRNAs on a Single Test Line in Catalytic Hairpin Assembly-Based SERS-LFA Strip. Anal. Chim. Acta 2021, 1178, 338800. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, H.; Wen, D.; Wang, L.; Li, L.; Kong, J.; Zhang, X. Ultrasensitive Electrochemical Detection of MiRNA Based on Polymerization Signal Amplification. Talanta 2021, 235, 122744. [Google Scholar] [CrossRef]
- Wu, X.; Li, Y.; Yang, M.Y.; Mao, C.B. Simultaneous ultrasensitive detection of two breast cancer microRNA biomarkers by using a dual nanoparticle/nanosheet fluorescence resonance energy transfer sensor. Mater. Today Adv. 2021, 12, 100163. [Google Scholar] [CrossRef]
- Gulati, P.; Kaur, P.; Rajam, M.v.; Srivastava, T.; Mishra, P.; Islam, S.S. Vertically Aligned Multi-Walled Carbon Nanotubes Based Flexible Immunosensor for Extreme Low-Level Detection of Multidrug Resistant Leukemia Cells. Sens. Actuators B Chem. 2019, 301, 127047. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, H.; Sun, F.; Wang, H.; Zhao, J.; Chen, B.; Wang, X. Rapid Diagnosis of Multidrug Resistance in Cancer by Electrochemical Sensor Based on Carbon Nanotubes-Drug Supramolecular Nanocomposites. Biosens. Bioelectron. 2011, 26, 3361–3366. [Google Scholar] [CrossRef]
- WHO. Antimicrobial Resistance—À Global Epidemic. 2010. Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance (accessed on 3 August 2022).
- Eduardo, C.R.; Laschi, S.; Ilaria Palchetti, E.T. Advances in Antimicrobial Resistance Monitoring Using Sensor and Biosensor: A Review; MDPI: Berlin, Germany, 2021. [Google Scholar]
- Global Multidrug Resistant Bacteria Market-Industry Trends and Forcast to 2029. Available online: https://www.databridgemarketresearch.com/reports/global-multidrug-resistant-bacteria-market (accessed on 15 August 2022).
- Monedero-Recuero, I.; Gegia, M.; Wares, D.F.; Chadha, S.S.; Mirzayev, F. Situational Analysis of 10 Countries with a High Burden of Drug-Resistant Tuberculosis 2 Years Post-UNHLM Declaration: Progress and Setbacks in a Changing Landscape. Int. J. Infect. Dis. 2021, 108, 557–567. [Google Scholar] [CrossRef]
- WHO Newsroom. Cancer. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer#:~:text=Cancer isa leading cause, and rectum and prostate cancers (accessed on 6 July 2022).
- Rebecca, A.J.; Seigel, L.; Miller, K.D.; Fuchs, H.E. Cancer Statistics; American Cancer Society: Atlanta, Georgia, 2022. [Google Scholar]
- Sin, M.L.; Mach, K.E.; Wong, P.K.; Liao, J.C. Advances and Challenges in Biosensor- Based Diagnosis of Infectious Diseases. Adolesc. Med. 2003, 14, 337–364. [Google Scholar] [CrossRef]
- Mahato, K.; Srivastava, A.; Chandra, P. Paper Based Diagnostics for Personalized Health Care: Emerging Technologies and Commercial Aspects. Biosens. Bioelectron. 2017, 96, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Global Biosensors Market Size to Reach USD 50.7 Billion in 2030|Rising Need for Real-Time Monitoring Devices and Increasing Demand for Point of Care Healthcare Services Are Some Key Factors Driving Industry Demand, Says Emergen Research. Available online: https://www.prnewswire.com/news-releases/global-biosensors-market-size-to-reach-usd-50-7-billion-in-2030--rising-need-for-real-time-monitoring-devices-and-increasing-demand-for-point-of-care-healthcare-services-are-some-key-factors-driving-industry-demand (accessed on 15 August 2022).
- Mancini, A.; Vito, L.; Marcelli, E.; Piangerelli, M.; de Leone, R.; Pucciarelli, S.; Merelli, E. Machine Learning Models Predicting Multidrug Resistant Urinary Tract Infections Using “DsaaS”. BMC Bioinform. 2020, 21, 347. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.H.; Khan, D.M.; Jamal, K.; Ahmad, Z.; Manzoor, S.; Khan, Z. Prediction of Multidrug-Resistant Tuberculosis Using Machine Learning Algorithms in SWAT, Pakistan. J. Healthc Eng. 2021, 2021, 2567080. [Google Scholar] [CrossRef] [PubMed]





| S.No. | Microorganism(s) (M/O) Detected | Target Analyte of the Biosensor | Method of Detection | Linearity | Limit of Detection | Response Time | Salient Features of the Biosensor Design or Principle | Reference |
|---|---|---|---|---|---|---|---|---|
| I. Genotypic Biosensors for MDR Microbe Detection | ||||||||
| 1 | Multiple M/O | ampR gene | Electrochemical | 1 pM to 1 nM | <1 pM (ss ampR) 4 pM (ds ampR) | 20 min | Capacitive DNA biosensor Label free Probe functionalized electrodes reusable for at least 6 cycles. | [44] |
| 2 | Staphylococcus aureus | mecA gene | Electrochemical | 10 to 166 pM | 10 pM | -- | Selective for MRSA and S. aureus 2 types of nanoparticle-modified probes were used. Linearity observed from 10–166 pM | [43] |
| 3 | S. aureus | mecA gene | Electrochemical | 0.075 to 200 pM | 63 fM | 2 h | Isothermal strand-displacement polymerization reaction based Methylene blue hairpin probes used on gold electrodes | [94] |
| 4 | M. tuberculosis | rpoB gene katG gene gyrA gene | Optical | -- | 1.5–13 nM | 1.5–2 h | Colorimetric detection system Deoxy ribozyme sensors used Point mutations in mentioned genes identified | [72] |
| 5 | Listeria monocytogenes | hlyA gene | Optical | -- | 10 CFU/mL | -- | • Can distinguish between dead and viable cells. • Visual detection and quantification. • Also, able to detect L. monocytogenes biofilm (mentioned further) | [70] |
| 6 | Aspergillus fumigatus | gliT gene | Electrochemical | 1 × 10−14 to 1 × 10−2 M | 0.32 ± 0.01 × 10−14 M | ≤20 min | • The self-assembled probes (gliP) were immobilized over Au electrodes. • Au nanoparticles were stabilized by 1,6-Hexanedithiol and chitosan. | [71] |
| 7 | M. tuberculosis | rpoB gene | Electrochemical | 1 fM to 0.1 pM | 4 fM or 3 × 104 copies of DNA | -- | • Polypyrrole-coated Fe3O4NPs functionalized with PAMAM dendrimers were used as scaffolds • The Naphthoquinone redox group and DNA probes were bound to the scaffolds | [66] |
| 8 | Acinetobacter baumannii | Carbapeneme resistance genes | Optical | -- | -- | <2 h | • Multiplex detection system • DNA extraction, PCR amplification, and silver nitrate-based colorimetric are essential components of the process. | [95] |
| 9 | M. tuberculosis | rpoB inhA | Optical | 20 µg/mL to 50 µg/mL | 30 µg/mL | 30 min | • A duplex colorimetric detection system • Au nanoprobes used | [96] |
| 10 | M. tuberculosis | rpoB gene | Electrochemical | -- | 1 nM | -- | • Thiolated DNA probes used • Impedimetric biosensor | [97] |
| 11 | M. tuberculosis | rpoB gene inhA gene gyrA gene katG gene 23S r RNA | Optical | -- | -- | -- | • Binary deoxy ribozyme sensors used • Fluorescence-based multiplex detection system • Can also detect other mutations | [73] |
| 12 | A. baumannii | β-lactamase gene | Optical | 102 to 105 CFU/mL | 50 CFU/mL | <= 2hr | • PCR and CRISPR-CAS-based multiplex fluorescence-based detection system fabricated in array format | [98] |
| 13 | -- | NDM1 gene | Electrochemical | 1 pg/L to 100 μg/L | 0.042 pg /L | 1 min | • No PCR amplification is required • Sandwich-type LNA electrochemical biochips used • Detection carried out in clinical samples | [45] |
| 14 | Escherichia coli Klebsiella pneumoniae | NDM1 gene | Optoelectrical | 100 copies | <3 min | • Thin Film Transistor sensors • Isothermal DNA amplification | [99] | |
| 15 | E. coli | blaNDM-5 gene blaCTX-M15 gene | Optical | 20 to 30 aM | 25 aM | <30 min | • Microfluidic system • Bimodal waveguide interferometric biosensor detecting carbapenemase and ESBL encoding genes | [100] |
| 16 | M. tuberculosis | DNA | Electrochemical | 1 fM to 100 pM | 0.2 fM | -- | • Can detect single point/nucleotide mutation | [75] |
| 17 | M. tuberculosis E. coli | DNA | Electrochemical | -- | 5 nM | Few minutes | • Duplex Surface Plasmon Resonance (SPR) sensing system • Can be extended to the detection of other biomolecules too | [74] |
| 18 | E. coli K. pneumoniae P. aeruginosa S. aureus Enterobacter faecalis | DNA | Optical | 102 to 103 CFU/mL | 4.5 CFU/mL | -- | • Surface-enhanced Raman spectroscopy (SERS) based detection • Au-Ag core-shell nano dumbbells used | [101] |
| 19 | S.aureus | DNA | Electrochemical | -- | 100 fM | -- | • An electrode made of reduced graphene oxide was used. | [102] |
| 20 | M. tuberculosis | DNA | Optical | 10−12 M to 10−8 M | 5 pM | -- | • SER-based detection • Au nanoparticles modified probes • Enhanced surface-anchored rolling circle amplification employed • The positive mutation detection is achieved with a wild-type to the mutant ratio of 5000:1 | [103] |
| 21 | M. tuberculosis | DNA | Electrochemical | -- | ~nM | -- | • Able to detect single nucleotide substitution in folded NA structure | [104] |
| 22 | Enterococcus | DNA | Optical | -- | 102 CFU/mL | 45 min | • Colorimetric detection clubbed with loop-mediated isothermal amplification. • Vancomycin-resistant Enterococcus was detected. | [105] |
| 23 | S. aureus | DNA | Optical | -- | 10 CFU/mL | <20 min | • SPR-based detection • Distinguishes between MRSA MSSA and borderline oxacillin-resistant Staphylococcus aureus | [106] |
| 24 | L. monocytogenes | DNA | Electrochemical | -- | -- | -- | • Single-stranded DNA probe immobilized over Au surface. • Changes in cyclic voltammetry peak current were recorded | [107] |
| 25 | M. tuberculosis | DNA | Optical | -- | 1µM | <20 min | • Label-free DNA detection and amplification | [108] |
| 26 | M. tuberculosis | DNA | Electrochemical | 10−18 moL/L to 10−14 moL/L | 0.330 aM | -- | • G4-hemin used as an enzyme | [109] |
| 27 | E. coli | DNA | Electrochemical | 10−6 to 10−16 M | 0.1 fM | -- | • Graphene oxide-nickel ferrite-chitosan nanocomposite film-based sensing platform | [110] |
| 28 | S. aureus | DNA | Diffusion based | 10 to 60 pM | 10 pM | 10 sec | • Based on nanobead diffusometry and non-PCR-based DNA monitoring | [111] |
| 29 | Salmonella spp. S. aureus E. coli | DNA | Optical | -- | 3.0 × 102 CFU/sample (Gram-negative) 3.0 × 103 CFU/sample (Gram Positive) | <2 h | • Nucleic acid testing • Performs DNA extraction, polymerase chain reaction, and on-site colorimetric detection for point-of-care diagnosis • Multiplex detection system • DNA extraction and PCR amplification can be performed. • Colorimetric biosensor | [95] |
| 30 | Plasmodium falciparum | C580Y mutation | Electrochemical | -- | 1 copy/reaction volume | <25 min | • Potentiometric biosensor • Ion-Sensitive Field-Effect Transistors based lab-on-chip model. | [68] |
| 31 | Influenza virus | RNA | Optical | -- | 10 copies/mL of RNA | -- | • Fluorescence-based detection • 10 copies/mL of RNA from the resistant strain among 2 × 104 copies/mL of RNA from the sensitive strain | [76] |
| 32 | Hepatitis B Virus | DNA | Electrochemical | 4 × 10−10 to 1 × 10−8 mol | 1 × 10−11 mol | -- | • Nonporous gold platform • Low cycles of PCR for amplification are required by the use of the Au platform | [112] |
| II. Phenotypic Biosensors for MDR Microbe Detection | ||||||||
| Phenotypic Biosensors Detecting the MDR Microbes Themselves | ||||||||
| 33 | E. coli | M/O | Optical | 3.81 × 102 to 2.44 × 104 CFU/mL | 460 CFU/mL | -- | • Real-time detection in human urine, tap water, and apple juice • Colistin-modified carbon dots used • Fluorescence-based detection. | [113] |
| 34 | S. aureus E. coli | M/O | Optical | 9 × 107 CFU/mL | -- | 2 h | • Photoluminescence-based biosensor. • Graphene quantum dots based. | [114] |
| 35 | E. coli | M/O | Optical | 105–108 CFU/mL | 9.5 × 104 CFU/mL | -- | • Fluorescence-based detection • Water-soluble carbon dots used • Efficient in HeLa cell imaging | [115] |
| 36 | E. coli Desulfovibrio desulfuricans S. sciuri L. monocytogenes S. aureus Pseudomonas aeruginosa | M/O | Optical | -- | -- | -- | • Multiplex detection and differential analysis of microbes • Carbon dots functionalized with 3 different receptors, boronic acid, polymyxin, and vancomycin, present on the fluorescence-based array • Discrimination of the six kinds of bacteria with 91.6% accuracy | [77] |
| 37 | S. aureus E. coli | M/O | Optical | 101 to 107 CFU/mL | 3 CFU/mL 3.5 CFU/mL, respectively | ~2 h | • A multifunctional alternative current electro-kinetic SERS-based microfluidic system. • Can concentrate bacteria from whole blood, identify bacterial species, and determine antibiotic susceptibilities of the bacteria rapidly. • Label-free antibiotic susceptibility testing is possible with the device. | [116] |
| 38 | S. aureus | M/O | Optical | 10 to 106 CFU/mL | 6.9 CFU/mL | -- | • Bacteria imprinted film with N-Succinyl-Chitosan doping. • Fluorescence-based sensor • Au disulfide NP used. | [117] |
| 39 | S. aureus | M/O | Electrochemical | 10 to 107 CFU/mL | 5 CFU/mL | 30 min | • 3D porous copper nanocomposite modified with vancomycin was used. Also designed for the treatment of MRSA. • MIC:1.93 μg/mL | [79] |
| 40 | E. coli | M/O | Optical | 5.0 × 101 to 1.0 × 109 CFU/mL | 50 CFU/mL | -- | • Enzymatic redox reaction employed • CD-MnO2 nanosheets are used as a platform • Label-free fluorescent biosensor • Considerable selectivity for E. coli | [118] |
| 41 | S. aureus K. pneumoniae | M/O | Optical | 20 to 108 CFU/mL | ~20 CFU/mL | 15 min | • Aptamer-coated magnetic beads used • A broad-spectrum fluorescent probe was used. • MRSA and Klebsiella pneumoniae carbapenemase 2-expressing Klebsiella pneumoniae (KPC-2 KP) can be detected • Crystallizable mannose-binding lectin-coated Au nanoclusters-based duplex detection system | [78] |
| 42 | E. coli | M/O | Optical | -- | 1.6 × 103 CFU/min | 10 min | • Optically induced electrophoresis phenomena are used to segregate resistant and non-resistant bacteria in a heterogeneous sample. | [119] |
| 43 | S. aureus K. pneumoniae E. coli | M/O | Optical | 1 × 102 to 1 × 106 CFU/mL | 67CFU/mL 57CFU/mL 61CFU/mL | 4 h | • DNAzyme integrated with SPR system in the biosensor | [120] |
| 44 | P. aeruginosa | M/O | Optical | 101 CFU/mL to 107 CFU/mL | 9 CFU/mL | -- | • Aptamers conjugated with photoluminescent carbon dots as probes • Graphene oxide is used as a quencher | [121] |
| 45 | Salmonella infantis | M/O | Optical | -- | 100 CFU/mL | 1 h | • Anti-salmonella antibodies were adsorbed on single-walled carbon nanotubes • Field Effect Transistor (FET) based biosensor | [122] |
| 46 | A. baumannii | M/O | Optical | 1 × 104 to 5 × 107 CFU/mL | 2.3 × 103 CFU/mL | -- | • Diagnosis in sputum • Photoluminescent Au-Ag nanoclusters used | [123] |
| 47 | S. aureus A. baumannii | M/O | Electrochemical | -- | 104 cells/mL | 5 min | • Single-cell detection of antibiotic-resistant bacteria. • Voltametric biosensor | [124] |
| 48 | S. aureus | M/O | Optical | 102 to 107 CFU/mL | 33 CFU/mL | 20 min | • IgY-modified immunosensor used. • Based on long-period fiber grating | [125] |
| 49 | Candida albicans Cryptococcus neoformans | M/O | Optical | 0 to 2 µM | -- | - | • Can also be used for Fe detection • Fluorescence-based biosensor • Uses N-doped carbon dots obtained from Chionanthus retusus | [126] |
| Phenotypic Biosensors Detecting MDR Associated Analytes | ||||||||
| 50 | 20 different strains with extended-spectrum ß-lactamase (ESBL) activity | β-lactamase activity | Optical | -- | 10 CFU/mL | 90 min | • BODIPY fluorescence-based probe was used • Can identify ceftazidime-resistant bacteria | [83] |
| 51 | P. aeruginosa | Pyocyanin | Electrochemical | 1–100 μM | 0.27 μM(PBS) 1.34 μM(Saliva) 2.3 μM(Urine) | -- | • Reduced graphene oxide with Au nanoparticles used | [91] |
| 52 | M. tuberculosis | Mannose-capped lipoarabinomannan | Optical | 5 fg/mL to 10 pg/ mL (PBS) 10 fg/mL to 10 pg/ mL (synthetic urine) | 1–10 fg/mL | -- | • A plasmonic fiber optic biosensor (P-FAB) strategy used | [85] |
| 53 | E. coli | Endotoxin | Electrochemical | 0.0005 to 5 EU/mL | 0.0002 EU/mL | -- | • rhTLR4/MD-2 complex is the Bio-recognition Element (BRE) • Au electrodes used • High specificity | [88] |
| 54 | M. tuberculosis | MPT64 protein | Electrochemical | 1 to 50 nM | 81 pM | 30 min | • Aptamers used as BRE • Gold electrode used | [127] |
| 55 | K. pneumoniae | Carbapenemase | Electrochemical | 1 × 10−12 to 1 × 10−7 mol/L | 0.2 pM | -- | • Glassy carbon electrode modified with Au nanoparticles and graphene nanocomposite used | [128] |
| 56 | Escherichia coli | ESBL production | Optical | -- | 105 CFU/mL | 20 min | • β-lactamase activity monitored • CENTA used as β-lactamase reporter • SER-based paper biosensor | [129] |
| 57 | S. aureus | α-haemolysin | Optical | 0.012 to 0.76 µM | 0.002 µM | <30 min | • SPR-based system of detection using a cantilever system in combination with molecular imprinted gold chips. • Detection from septic blood samples | [130] |
| Phenotypic Biosensors Detecting Biofilms | ||||||||
| 58 | S. aureus E. coli P. aeruginosa | Biofilm | Electrochemical | -- | 104 − 105 CFU/cm3 | -- | • Monitored biofilm growth • Graphene oxide-based potentiometric biosensors | [131] |
| 59 | E. coli | Biofilm | Mechanical | -- | 5.3 pg | -- | • An atomic layer deposition aluminum oxide sensor was used protected by ZnO • Surface Acoustic Wave (SAW) based detection | [93] |
| 60 | L. monocytogenes | Biofilm | Optical | -- | 1.164 × 101 CFU/mL (stainless steel) 1.021 × 101 CFU/mL (lettuce) | -- | • Can distinguish between dead and viable cells. • Visual detection and quantification of the hlyA gene | [70] |
| miRNA | Function | Expression Levels in the Case of MDR Phenotype | References |
|---|---|---|---|
| miR-21 | Regulatory role in apoptosis, development, and differentiation of normal cells. Role in metastasis and carcinogenesis. | Upregulated | [161,162] |
| miR-155 | Role in immune response, inflammation, and differentiation of hemopoietic lineages and tumorigenesis | Upregulated | [163,164] |
| miR-205 | Regulates cell survival, proliferation, and susceptibility to chemotherapy | Downregulated | [165] |
| miR-122 | Liver-specific miRNA. (70% of the liver’s miRNA pool) | Downregulated | [166,167] |
| miR-223 | Haematopoetic cell-specific miRNA. Important for the development of cells in myeloid lineage | Downregulated | [168,169,170] |
| miR-31 | Embryonic implantation and development, Muscle and bone homeostasis; Regulation of immune system function, and autoimmunity | Downregulated | [171] |
| miR-200a-3p | Inhibits malignant transformation and all stages of carcinogenesis | Downregulated | [172,173] |
| miR-34a | Tumor suppressor gene. Involved in the regulation of cell survival, migration, and remodeling properties | Downregulated | [143,174,175] |
| miR–k12-5-5p | Coded by Kaposi’s sarcoma (KS) associated with the herpes virus. Works in inhibiting replication and as a transcription activator | Upregulated (in metastasis and cell growth) and KS | [176,177,178] |
| miR-410 | May promote or suppress tumor formation. | Downregulated | [179,180] |
| miR-196a-5p | Involved in metastasis | Upregulated | [181,182] |
| miR-141 | Tumor suppressor gene | Upregulated | [183,184] |
| Let7 miR family | Roles in embryogenesis, tumorigenesis, development, and metabolism | Downregulated | [185,186,187] |
| Survivin | Inhibitor of apoptosis. Regulates cellular proliferation and death | Upregulated | [188,189] |
| S.No | Target Analyte Detected by the Biosensor | Method of Detection | Linearity | Limit of Detection | Response Time | Cell Lines/Samples Used | Salient Features of the Biosensors’ Principle/Design | Reference |
|---|---|---|---|---|---|---|---|---|
| I. GENOTYPIC BIOSENSORS | ||||||||
| I.i Genotypic Biosensors Detecting Mutations or Gene Segments | ||||||||
| 1 | MDR1 gene | Electrochemical | 1.0 × 10−14 to 1.0 × 10−7 M | 3.12 fM | 3.4 h | Clinical leukemic samples | Label-free biosensor N-doped graphene nanosheets functionalized over Au nanoparticle | [152] |
| 2 | MDR1 gene | Electrochemical | 1.0 × 10−11 to 1.0 × 10−9 M | 2.95 pM | -- | -- | Au nanoparticle/ toluidine blue–graphene oxide-modified electrodes were used. | [151] |
| 3 | EGFR T790 M mutation 16 drug-sensitive mutations | Optical | -- | 1–4 copies | 3–5 min | Plasma | Detection of MDR leukemia Vertically aligned multi-walled carbon nanotubes based immunosensor used Cell-free circulating DNA analyzed Multiplex detection system Fluorescence-based detection | [154] |
| I.ii. Detection and monitoring of miR | ||||||||
| 4 | miR-121 miR-155 miR-205 | Optical | 1 fM to 1 nM (for miR- 121) | 20.20 fM 15.32 fM 13.50 fM | -- | HeLa MCF-7 | Mo2B-based FL quenching platform Intracellular monitoring in live cell Fluorescence-based detection | [219] |
| 5 | miR-223 miR-122 miR-21 | Optical | 0.02 nM-10 nM | -- | Liver cancer | Förster resonance energy transfer (FRET) based detection Multiplex quantification system for the miR Successful detection in 10% of serum samples was achieved. | [220] | |
| 6 | MiR-21 Let-7d | Optical | -- | 33.93 pM | ~2 h | -- | Single-stranded DNA (ssDNA) used Detection instrumented by flow cytometry Fluorescence-based detection | [221] |
| 7 | miRNA 21 miRNA 31 | Optical | 50 to 200 fM (miRNA 21) 1.0 to 200 fM (miRNA 31) | 0.20 fM (miRNA 21) 0.50 fM (miRNA 31) | <40 min | HeLa A549 | Microfluidic paper-based system Laser-induced fluorescence used | [158] |
| 8 | miR-200a-3p | Optical | -- | 1 aM | 1–2 h | MKN45 and SNU1 | Single-base mismatch detection is possible too No amplification required | [222] |
| 9 | Survivin | Optical | -- | 827 pM | -- | HeLa | Au nanoparticles used Cy3 and Cy5 dyes were used as donors FRET-based approach | [223] |
| 10 | miR-21 | Optical | 1 pM to 10 nM | 55 fM | -- | MCF7 | Cyclic enzymatic amplification was achieved with help of a periodic nanostructure sensor chip Photonic biosensor | [224] |
| 11 | miR-34a | Electrochemical | 5 to 35 μg/mL | 7.52 μg/mL | ~1 min | -- | Label-free voltammetric detection | [225] |
| 12 | miR-21 | Optical | 5 pM to 200 nM | 0.5 pM | -- | Serum | Graphdiyne/graphene quantum dots were used. Detection of MCF-7 cells and live imaging of MB231 done FRET-based detection | [226] |
| 13 | miR-K12–5-5p | Optical | -- | 0.884 nM | -- | Breast cancer | SERS based biosensor GaN with Au/Ag used as the platform | [227] |
| 14 | miR 410 | Electrochemical | 10 fM to 300 pM | 3.90 fM | 5 min | Prostate cancer cells and serum | Disposable Au nanoparticles -peptide nanotubes used Impedimetric biosensor | [159] |
| 15 | miR 21 miRNA-196a-5p | Optical | 10 pM to 10 mM | 3.31 pM 2.18 pM | 30 min | Non-small cell lung cancer | SERS based biosensor Catalytic hairpin assembly-based SERS-LFA strip | [228] |
| 16 | miR- 141 | Electrochemical | 10 pM to 10 aM | 3.23 aM | -- | Prostrate and breast cancer | Sensing platform based on atom radical polymerization | [229] |
| 17 | miR-593 miR-155 | Optical | 5 nM to 50 nM | 0.17 nM 0.25 nM | <3 h | Body fluids and tumor tissues | FRET-based biosensor Breast cancer cell biomarkers are being identified. Based on core-shell upconversion NP and MoS2 nanosheets were used. | [230] |
| II. PHENOTYPIC BIOSENSORS | ||||||||
| II.i. Phenotypic Biosensors Detecting Drug Efflux Pump | ||||||||
| 18 | P-glycoprotein (Pgp) | Optical | 1.1 × 107 − 2 × 103 cells/mL | 27 cells/mL | -- | Chronic myeloid leukemia | Fermi level fluctuation induced charge transfer-based biosensor | [193] |
| 19 | Pgp | Electrochemical | 50 and 100,000 cells/mL | 23 ± 2 cells/mL | -- | MDRCC | Amperometric biosensor Monoclonal Pgp antibodies and amino phenylboronic acid used as BRE immobilized over Au nanoparticles | [192] |
| 20 | Pgp | Optical | 1.5 × 102 to 1.5 × 107 cells/mL | 10 cells/mL | -- | K562 cells | Fluorescence-based detection Detection of MDR leukemia Vertically aligned multi-walled carbon nanotubes based immunosensor used | [231] |
| 21 | MRP2 protein | Optical | -- | 1 cell | -- | HepG2 | Fluorescence-based detection A multifunctional gradients-customizing microfluidic device | [194] |
| 22 | Cell itself | Electrochemical | -- | ~50 cells | <12 min | Leukemia K562 | Au nanoparticles -modified glassy carbon electrodes (GCE) used | [190] |
| II.ii. Phenotypic Biosensors Observing Cellular Profile and TME | ||||||||
| 23 | Membrane protein profile | Optical | -- | 200 cells | -- | MDA-MB-231 | Fluorescence-based detection. Healthy, cancerous, and metastatic human breast cancer cells can be differentiated. Dual ligand co-functionalized Au-clusters | [191] |
| 24 | pH | Optical | pH range of 4.2–6.4 with a pKa value of 5.18 | -- | ~1 min | RAW 264.7 | Pyrido [1,2-a] benzimidazole derivative-based fluorescent probe | [214] |
| 25 | Nitroreductase (NTR) | Optical | 0 to 20 μg/mL | 26 ng/mL | -- | A549 A549/DDP | NTR is overexpressed in highly hypoxic TME Detection based on fluorescence | [213] |
| 26 | NTR | Optical | 0 to 4 μg/mL | 18.6 and 33.2 ng/mL of NTR for 1-NO2 and 2-NO2 | - | A2058 | Detection based on fluorescence. Sensors were designed based on the conjugation of pyridazine-1,3a,6a- triazapentalene to a para-nitrophenyl, forming two probes denoted as 1- NO2 and 2-NO2. Reduction by NTR led to the over 15- fold enhancement of fluorescence intensity in both probes. | [212] |
| 27 | CD59 protein | Electrochemical | 1 fg/mL and 1000 fg/mL | 0.38 ± 0.03 fg/mL | 10 min | Saliva | Impedimetric label free biosensor Protein probe (anti CD59 antibody) immobilized over a self-assembled monolayer of Cys over Au electrode | [200] |
| II.iii. Phenotypic Biosensors Detecting Enzymes and Proteins | ||||||||
| 28 | Glutathione | Optical | 0 to 80 μM | 87 nM | -- | HeLa HepG2 LO2 | Fluorescence-based biosensor Nanoscale metal-organic frameworks are used. Two-photon imaging used | [209] |
| 29 | Brain-Derived Neurotrophic Factor (BDNF) | Electrochemical | 4.0 to 600.0 pg/mL | 1.5 ± 0.012 pg/mL | -- | Serum PC12 SH-SY5Y | Microfluidics based immunosensor Biconjugate probe consisting of anti-BDNF and toluidine blue Was used to study the effects of nicotine, ethanol, and potassium ion on BDNF expression in cancer lines. | [202] |
| 30 | β1 Integrin | Electrochemical | 1.0 × 104 to 2.0 × 106 cells/mL−1 | 3.5 × 103 cells/ mL | -- | HeLa | Impendence spectroscopy-based sensor. Glass carbon electrode impinged with mouse anti-human integrin β 1 monoclonal antibody | [198] |
| 31 | Intracellular telomerase | Optical | 0 to 20,000 cells | 280 A549 cells | -- | A549 HepG2 MCF-7 | Fluorescence-based biosensor. Nanoflare and hybridization chain reaction (HCR)-based signal amplification was applied together with gold/carbon nanosphere | [203] |
| 32 | DNA methyl transferase | Optical | 0.1 to 0.2 U/μL | 1.6968 × 10−4 U/μL | -- | -- | Fluorescence-based biosensor Dumbbell-shaped DNA template copper nanoparticles used | [211] |
| III. Biosensors Based On Drug Pharmacokinetics | ||||||||
| 33 | Exosomes and exosomal cisplatin | Optical | Cisplatin: 0 to 0.2 µg/mL | Cisplatin: 0.17 µg/mL Exosome: 65 nM | -- | OVCA | SER based biosensor Cysteine-capped gold nanoparticles used Diagnosis of chemoresistance with accuracy greater than 90% | [216] |
| 34 | Methotrexate | Optical | 28 to 500 nM | 155 nM | -- | Serum and clinical samples | Multichannel SPR-based instrument | [215] |
| 35 | Daunomycin Residue | Electrochemical | -- Linearity coefficient: 0.995 | -- | 1 h | K562/A02 cells | Based on carbon nanotubes–drug interaction. Nanocomposites of daunorubicin and CNT were used. | [232] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jha, N.G.; Dkhar, D.S.; Singh, S.K.; Malode, S.J.; Shetti, N.P.; Chandra, P. Engineered Biosensors for Diagnosing Multidrug Resistance in Microbial and Malignant Cells. Biosensors 2023, 13, 235. https://doi.org/10.3390/bios13020235
Jha NG, Dkhar DS, Singh SK, Malode SJ, Shetti NP, Chandra P. Engineered Biosensors for Diagnosing Multidrug Resistance in Microbial and Malignant Cells. Biosensors. 2023; 13(2):235. https://doi.org/10.3390/bios13020235
Chicago/Turabian StyleJha, Niharika G., Daphika S. Dkhar, Sumit K. Singh, Shweta J. Malode, Nagaraj P. Shetti, and Pranjal Chandra. 2023. "Engineered Biosensors for Diagnosing Multidrug Resistance in Microbial and Malignant Cells" Biosensors 13, no. 2: 235. https://doi.org/10.3390/bios13020235