1. Introduction
Molecular oxygen (O
2) is essential for cell survival. While the atmospheric oxygen concentration remains constant at around 20.9%, the concentration among different parts of human tissues can be as high as 19.7% in the tracheas or as low as about 1% in the superficial region of the skin [
1]. When oxygen availability is compromised, cells act accordingly to sustain life. The collective actions are called hypoxic responses [
2,
3,
4,
5,
6,
7,
8,
9]. Studying the hypoxic response and the underlying oxygen-sensing mechanism is therefore important. In both physiological and pathological conditions, hypoxic responses play key roles in regulating various biological activities. For instance, hypoxic pulmonary vasoconstriction (HPV) is one of the essential physiological hypoxic responses. Pulmonary arteries constrict in response to alveolar hypoxia to improve gas exchange in the lungs and blood delivery to systemic tissues [
10].
In addition, tumor-induced hypoxia is observed among various types of cancers, with an average 1.5% to 2% of oxygen tensions in cancer biology. Tumor-induced hypoxia has been proven to enhance progression, metastasis, angiogenesis and therapeutic resistance [
11]. As a result, targeting the hypoxic signaling mechanisms of cancer cells is believed to possess therapeutic values. In chronic hypoxia in the core region of a solid tumor, drug targeting of the hypoxia-induced factor (HIF-1α) has been proven to be a promising therapeutic strategy [
12,
13]. Moreover, under cyclic hypoxia conditions in which the cell experiences intermittent hypoxia and normoxia environments, recent studies show HIF-2α is the dominant signaling factor for the fact that prolonged (>4–12 hr) cyclic hypoxic/normoxic cycles enhance HIF-2α but not HIF-1α accumulation among different cancer cell lines [
14]. This observation suggests that different signal transduction mechanisms may exist for chronic and intermittent hypoxia.
In order to study the hypoxic response and oxygen-sensing mechanism of cells, precise and consistent control of extracellular oxygen tension in spatiotemporal domains is highly desired. In conventional broadly conducted cell culture experiments, cells are cultured using a bicarbonate-buffered medium at 37 °C with 5% CO
2 enriched ambient air and humidity under non-condensing conditions in incubators. When performing hypoxic experiments using a gas control incubator/chamber, the typical response time taken to reach a steady state is within several tens of minutes to an hour, depending on the chamber size and setpoint value. After the culture environment reaches the setpoint, it further takes several additional minutes to hours to equilibrate the dissolved oxygen into the gas phase [
15]. Despite the gas-controlling incubation/chamber proving to be useful for chronic hypoxic research over the last 2 decades, a cell culture system capable of manipulating oxygen tension at faster responses is needed to explore acute hypoxic response and sensing mechanisms of the cells.
To overcome the challenges, an approach capable of reliably altering extracellular environments between different hypoxia levels and the normoxia condition with a relatively short response time is needed. Among various methods, microfluidics has been broadly exploited for cell culture in dynamic microenvironments due to its desired advantages, including a small sample volume and a high surface area to volume ratio which can greatly help to reduce the response time required to reach an equilibrium. For example, Polinkovsky et al. developed a microfluidic device capable of generating a series of oxygen concentrations with a short response time (in the order of seconds) through on-chip gas mixing to study cellular responses [
16]. In addition, Chen et al. exploited the spatially-confined chemical reaction method to rapidly establish oxygen gradients within the microfluidic channel within minutes for cell culture applications [
17]. The same chemical reaction method has also been applied to investigate cancer cell migration, endothelial cell migration and three-dimensional (3D) network formation under various oxygen microenvironments [
18,
19].
However, the existing approaches often require complicated instrumentation and do not provide great real-time cellular analysis capabilities, which are critical to studying temporal cellular responses. As a result, an integrated approach combining a microfluidic device, fluorescence dyes sensitive to extra/intracellular oxygen tensions and calcium concentrations, and real-time fluorescence imaging is developed in this paper to investigate temporal cellular hypoxic responses under various cyclic oxygen tensions. In the experiments, a single-layered microfluidic device capable of generating cyclic oxygen gradients and switching between hypoxic oxygen gradients and normoxic conditions within minutes is constructed for cell culture. The device possesses a simple architecture while allowing easy access for optical imaging and sensing. The device is mainly made of an elastomeric material, polydimethylsiloxane (PDMS) (due to its desired gas permeability), optical transparency, and cell compatibility. In the developed device, the oxygen gradient is generated by flowing gas with specific compositions into a microfluidic channel next to the cell culture channel separated by thin walls. The gas flow is controlled using a computer-controlled three-way solenoid valve. The single-layer device architecture with a good optical transparency of PDMS and a glass substrate makes real-time observations of the acute hypoxic response of cells using microscopes feasible.
For the hypoxic response study, two types of cells, smooth muscle cells and epithelial cells, were cultured inside the devices in the cell experiments for demonstration. The function of the device was first characterized to confirm its capability for cyclic oxygen gradient generation. The intracellular oxygen tensions and calcium levels were then measured in a time-lapse manner for the two distinct cell types under the generated cyclic oxygen gradients. The experiment results confirmed the capability of the device to generate the cyclic oxygen gradients for the cell experiments. Furthermore, the cell experiments showed the application of real-time fluorescence imaging for the study of cellular hypoxic responses under the cyclic oxygen gradients in variation of the intracellular oxygen tensions and calcium levels. The cell experimental results indicated distinct responses of the two tested cell types and suggested the importance of investigating hypoxic responses of the cells in the temporal domain. With the short response time, high throughput on control of oxygen tensions and high compatibility to optical assays of the device, the developed approach integrating the real-time fluorescence imaging serves as an efficient method to explore cell kinetic responses under extracellular oxygen tension variation as well as critical oxygen tension triggering acute hypoxic responses and underlying mechanisms for various biomedical studies.
2. Materials and Methods
2.1. Device Design and Fabrication
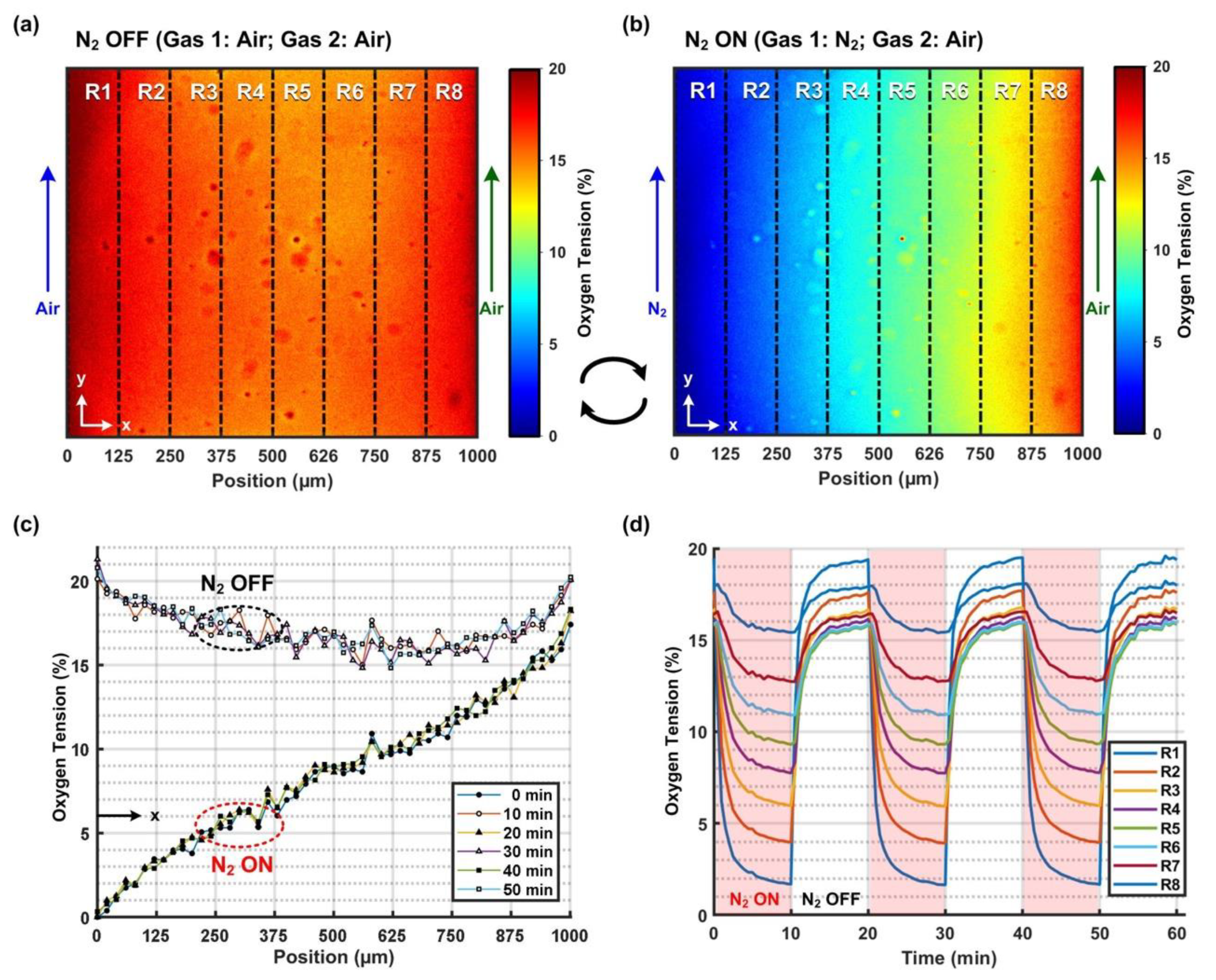
The microfluidic device capable of generating cyclic oxygen gradients is made of PDMS, and it is composed of a top layer with microfluidic channel patterns and a PDMS-coated glass bottom layer. Three sets of microfluidic channels were designed in the top layer, including one 1 mm-wide middle channel for cell culture and two sets of 100 μm-wide side channels (
Figure 1a). The channels were all designed with single inlets and outlets. Air or nitrogen gas was injected into the side channel from the inlet for the cyclic oxygen gradient generation. The middle and side channels are separated by 60 μm-wide oxygen-permeable PDMS thin walls to prevent direct chemical contact and gas bubbling into the medium while altering oxygen tensions in the middle channel.
The top layer was fabricated using the well-developed soft lithography replica molding process. A master mold with positive relief features of the microfluidic channels was fabricated on a 4-inch silicon wafer using negative tone photoresist (SU-8 2050, MicroChem Co., Newton, MA, USA) via conventional optical photolithography. To prevent the adhesion of the cured PDMS onto the mold, the fabricated mold was silanized using 1H,1H,2H,2H-perfluorooctyltrichlorosilane (78560-45-9, Alfa Aesar, Tewksbury, MA, USA) for 30 min in a desiccator at room temperature. A PDMS pre-polymer with 1:10 (
w/
w) of curing agent and base was mixed, poured onto the mold, degassed in a desiccator, and then cured at 60 °C for more than 4 h. After curing, the interconnection inlets and outlets were created via a biopsy punch with a diameter of 1.5 mm. For the bottom layer fabrication, a glass slide was spin-coated with the aforementioned PDMS pre-polymer and cured at 60 °C for more than 4 h. Both layers were treated with oxygen plasma (PX-250, Nordson MARCH Co., Concord, CA, USA) at 95 W for 35 s, sealed against each other, and heated at 60 °C for more than 4 h to promote the bonding between the layers.
Figure 1b shows the experiment photos of the fabricated devices filled with food dyes. The simple device design and fabrication process make the device easy to set up for practical biological applications.
2.2. Oxygen Gradient Generation and Characterization
In order to generate cyclic oxygen gradients within the middle cell culture channel, mixed air (air with 5% CO
2) was constantly introduced into one side channel and pure nitrogen (i.e., 0% O
2) and the mixed air were alternately introduced into another side channel with a period of 10 min. Both mixed air and nitrogen gases were injected into the side channels with inlet pressures of 0.4 bar controlled via manual gas regulators. The alternation between the gases was accomplished by switching the power of a three-way solenoid valve (ZHV-0519, ZonHen Co., Ltd., Shenzhen, China) on/off using a personal computer-controlled DC power supply (E3631A, Agilent Technologies, Inc., Santa Clara, CA, USA) via a GPIB interface, as shown in
Figure 1c. In the experiments, the solenoid valve was operated under 5 V/0.2A. The photo of the experimental setup is shown in
Figure S1 in the Supplementary Information. By introducing the gas with a specific oxygen composition into the side channel, the oxygen tension in the ambient air surrounding the channel can be adjusted to that of the introduced gas. The method has been broadly applied to control cell culture oxygen microenvironments in various microfluidic devices with relatively simple instrumentation [
20].
In order to characterize the oxygen tensions generated in the middle cell culture channel, an oxygen-sensitive fluorescence dye, Tris(2,2′-bipyridyl)dichlororuthenium(II) hexahydrate (RTDP) (224758, Sigma-Aldrich, Inc., St. Louis, MO, USA) was perfused into the middle channel with a flow rate of 10 μl/min. For the RTDP intensity profile measurement, 25% power blue LED illumination, 200 ms exposure, no average and binning were set as imaging parameters. The fluorescence intensity of the dye was quenched by the presence of oxygen; therefore, the spatial distribution of the oxygen tension inside the channel can be estimated by measuring the intensity profile [
21]. The relationship between the fluorescence intensity and oxygen tension is given by the Stern-Volmer equation as
, where
and
are the fluorescence intensity values with and without quencher (oxygen), respectively.
represents the quenching coefficient, and
is the concentration of the quencher (oxygen). The quenching coefficient
was calibrated by introducing pure nitrogen and oxygen, representing 0% and 100% oxygen tension, into the side channels, respectively [
17]. The calibrated quenching coefficient was further validated by calculating the oxygen tension under mixed air gas (ambient air with 5% CO
2). Since oxygen tension in air is approximately 20.9%, the oxygen tension in the mixed air should be around 20.9 × 95% = 19.9% with an additional 5% CO
2 mixed in the air. The resulted oxygen tension needs to be close to 20% to ensure the proper estimation of the quenching constant for the accurate oxygen tension characterization in the device.
2.3. Cell Culture and Seeding
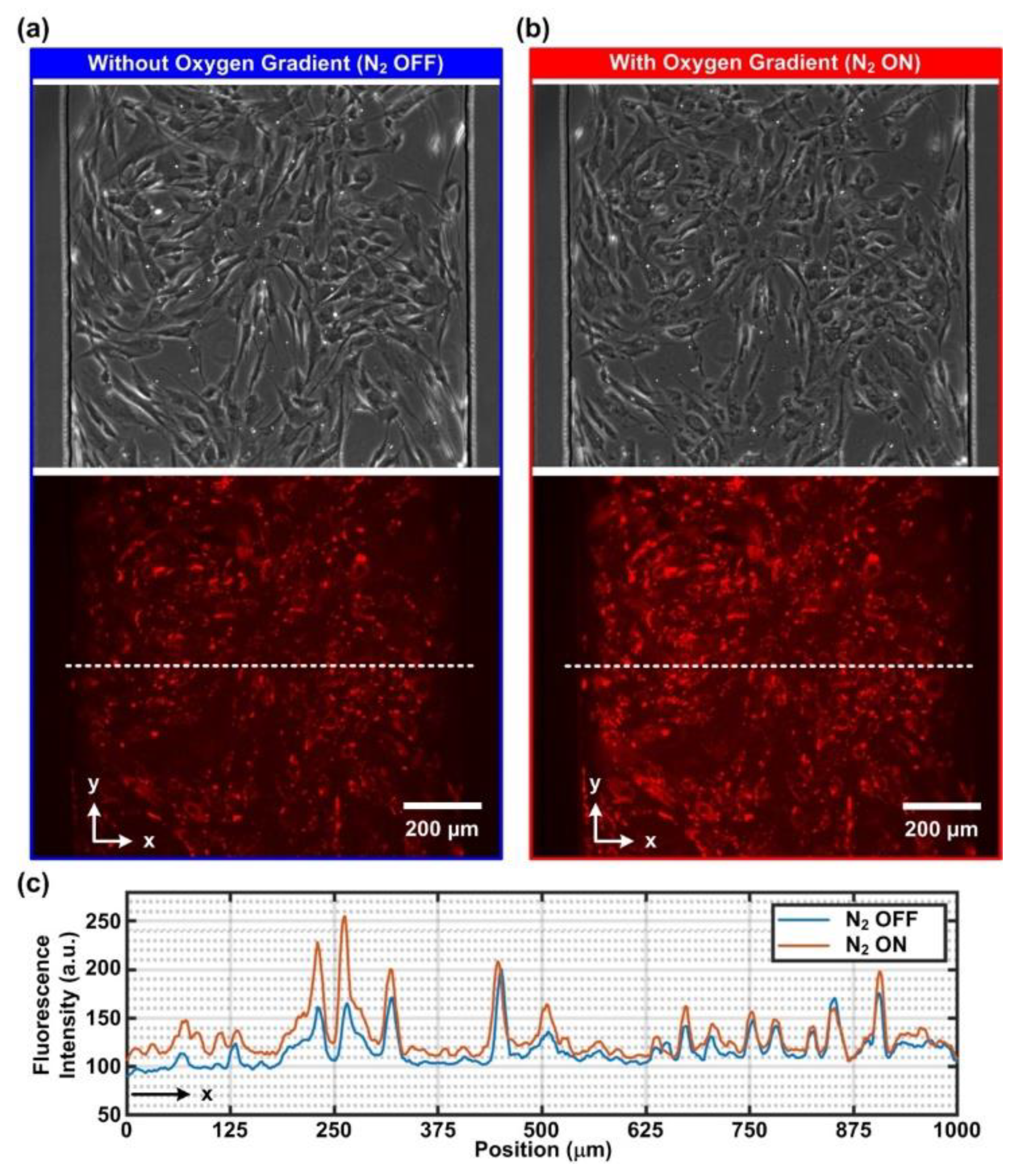
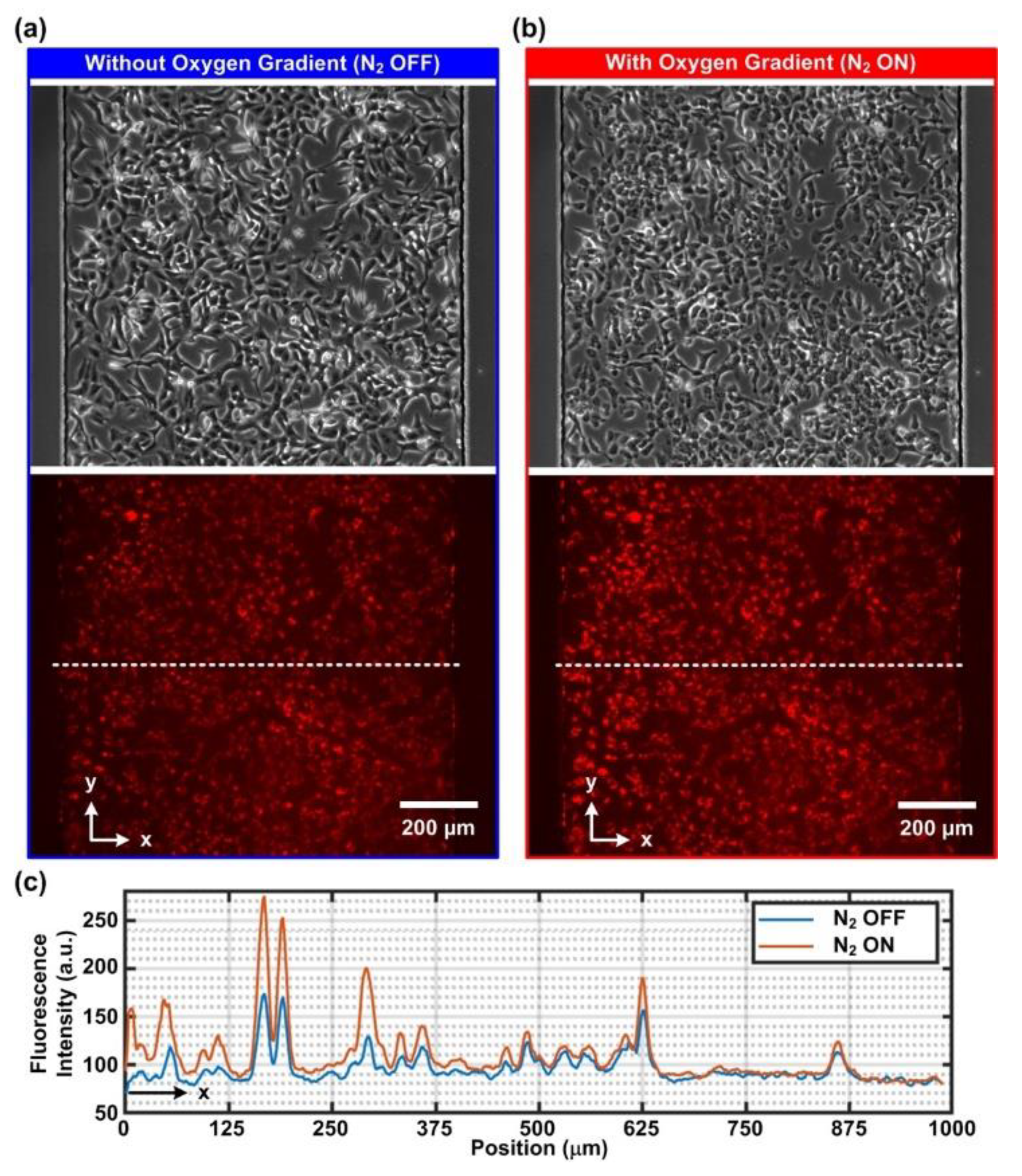
To demonstrate the capability of the integrated approach, two types of cells, smooth muscle cells and epithelial cells, were cultured and investigated in the experiments. First, a human aortic smooth muscle cell (AoSMC) (CC-2571, Lonza, Basel, Switzerland) was exploited. The cells were cultured in a smooth muscle cell basal medium (CC-3182, Lonza) with growth factor supplements (CC-4149, Lonza) and maintained as per the following process. The attached cells were trypsinized with trypLE (12604013, Gibco), centrifuged at 209× g for 5 min and resuspended in 1 mL of the prewarmed medium. After cell counting, 7 × 104 cells were seeded into a 25 cm2 flask (156367, Thermo Fisher Scientific, Waltham, MA, USA). The medium was changed every 2–3 days, and the cells were passaged when they reached 70–80% confluency, which took around 1 week. For optimized cell conditions and consistent results, the AoSMCs used in the experiments were all under 10 passages. In addition, human alveolar basal epithelial cells (A549, ATCC, Manassas, VA, USA) were also cultured in the experiments for comparison. The cells were cultured in DMEM (Gibco 10566, Thermo Fisher Scientific) with 10% v/v FBS (Gibco 10082, Thermo Fisher Scientific) and 1% v/v antibiotics (Gibco 15240, Thermo Fisher Scientific). The cell stocks were all maintained in a 37 °C humidified incubator supplied with 5% CO2. Cell suspension used for the cell culture in the devices was prepared by a process similar to the routine passage, except that after centrifugation, cell suspension was concentrated by resuspending the pellets with 100 μL of the prewarmed medium. After cell number counting, concentrated cell suspension was further diluted into final concentrations of 5000 cells/μL and 10,000 cells/μL for AoSMC and A549, respectively.
Before seeding the cells into the microfluidic device, the cell culture channel was first made hydrophilic through oxygen plasma treatment at 95 W for 35 s. The channel was then coated with a cellular matrix protein, fibronectin (FC010, Merck Millipore, Burlington, MA, USA), at a concentration of 100 μg/mL to promote cell attachment. The cell suspension was then pipetted into the cell culture channel from the inlet. Approximately 20 μL of cell suspension was needed for single device seeding. The cells were cultured inside the device for more than 18 h under static conditions to ensure the proper adhesion of the cells to the substrate for the following experiments.
2.4. Fluorescence Imaging Analysis
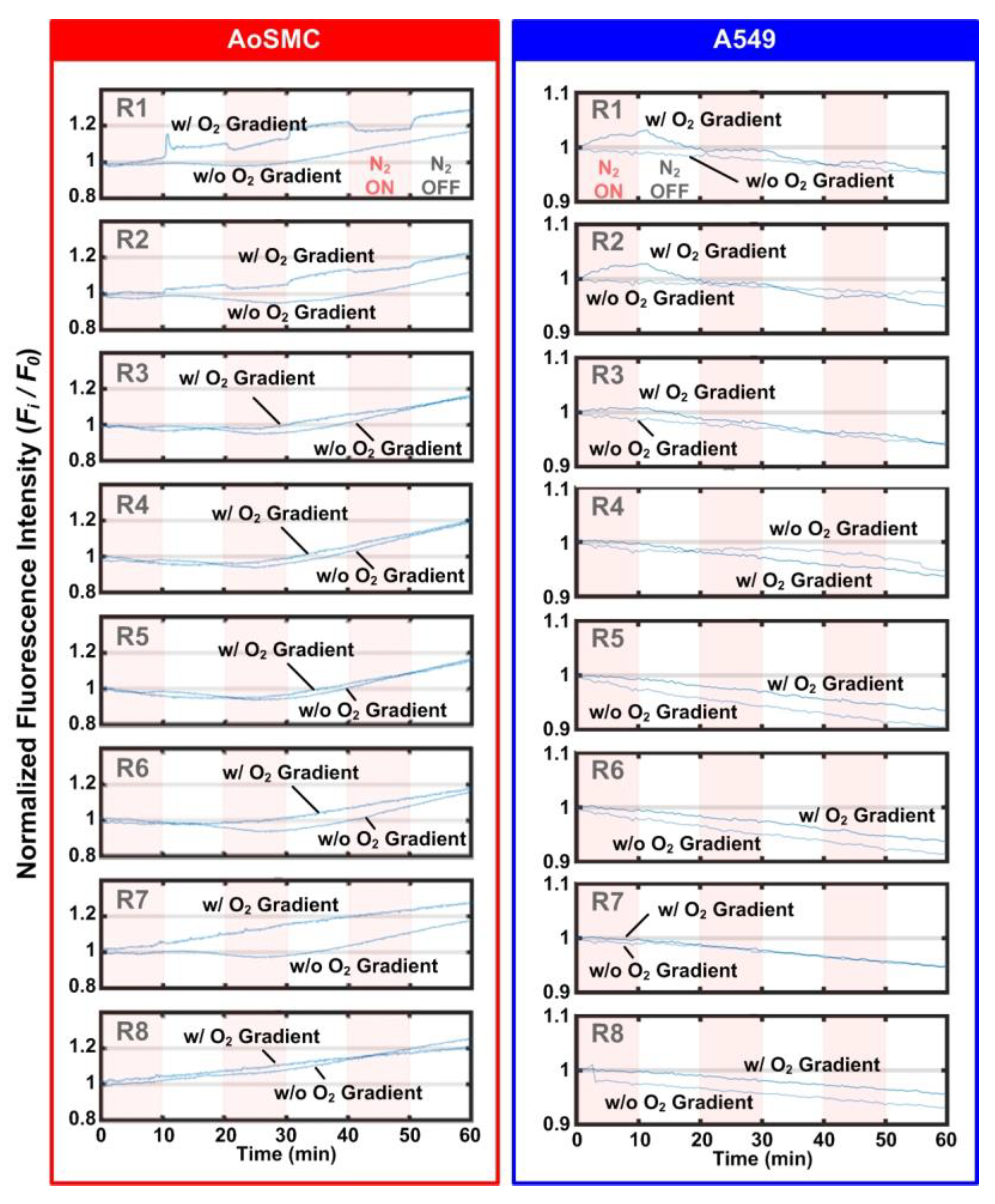
For the study of cellular responses under cyclic oxygen gradients, two types of analyses were performed in the experiments. First, the intracellular oxygen tension was characterized using a commercially available MitoXpress Intra intracellular oxygen assay (MX-300, Agilent Technologies, Inc.). The reagent was diluted with the complete cell culture medium at a volumetric ratio of 1:10 and applied into the cell-seeded device while the cells were attached to the culture surface. The cells were further incubated in the diluted reagent overnight.
In addition, intracellular calcium concentration was also investigated in the experiments. Calcium ion (Ca2+) is an essential intracellular messenger regulating various cell functions. Monitoring calcium levels under temporal and spatial oxygen tension change potentially gives insights into understanding the fast-acting oxygen sensing mechanism and hypoxic response behind it. In the Ca2+ measurement experiments, after the cell seeding and overnight culture, the growth medium inside the device was replaced by the complete medium with 3 μM of X-Rhod-1 Acetoxymethyl (AM) ester (Invitrogen X14210, Thermo Fisher Scientific). The cells were incubated for 30 min at 37 °C for dye loading. After the dye loading, the cells were then washed using the complete medium and incubated at 37 °C for an additional 30 min to make AM ester completely hydrolyzed and thus make X-Rhod-1 fluoresces.
The stained cells cultured in the microfluidic devices were imaged using an inverted fluorescence microscope (DM IL, Leica Microsystems, Wetzlar, Germany) equipped with a CCD camera (DFX360, Leica Microsystems, Wetzlar, Germany), a multi-color LED light source (X-Cite XLED1, Excelitas Technologies Corp., Waltham, MA, USA), and a 10X objective (HI Plan, Leica Microsystems). Control of the microscope and automatic time-lapse image acquisition were achieved through a commercially available microscopy automation and image analysis software (Metamorph Version 7.7, Molecular Devices, LLC, San Jose, CA, USA). For the intracellular oxygen characterization, a multiband filter cube suitable for MitoXpress Intra imaging (380–395 nm band-pass excitation filter, 575 nm dichromatic mirror, and 580–650 nm band-pass emission filter) was exploited (Semrock DA-FI-TR-3X-A-000, IDEX Health & Science, Rochester, NY, USA) (
Figure 1c). The imaging parameters were set as UV LED excitation at 5% intensity, 200 ms exposure, 4 times frame averaging and 2 × 2 binning. To minimize the photocytotoxicity during the imaging, the images were captured every 1 min for an hour. For the intracellular calcium observation, the same multiband filter cube with a blue band-pass excitation filter (470–490 nm), 500 nm dichromatic mirror and 500–535 nm band-pass emission filter was exploited. The imaging parameters were set as blue LED excitation at 5% intensity, 200 ms exposure and 2 × 2 binning. For AoSMC and A549, the images were captured every 2 s and 30 s for an hour, respectively.
The captured time-lapsed fluorescence images were quantitatively analyzed using the imaging analysis software ImageJ (Ver. 1.52p, NIH) with Time Series Analyzer plugin (
https://imagej.nih.gov/ij/plugins/time-series.html Access Date: 15 August 2020) and further visualized via a mathematics software, MATLAB (R2017a, MathWorks, Natick, MA, USA). All time-lapsed fluorescence intensity values were reported through fold change compared to the intensity obtained from the first frame in the same experiment (normalized fluorescence intensity, F
i/F
0). For intracellular oxygen measurements, baseline correction was applied by fitting the time-lapsed normalized fluorescence intensity data against asymmetric truncated quadratic function to compensate for the effect of photobleaching.
4. Conclusions
In this paper, an integrated approach combining microfluidic devices and real-time fluorescence imaging was developed to study cellular hypoxic response under cyclic oxygen gradients. To demonstrate the capability of the approach, we measured the intracellular oxygen tension and calcium levels of two distinct human cells (AoSMC and A549 cell) and discussed their differences. The intracellular oxygen tension measurement indicated that, even though extracellular and intracellular oxygen tension is closely related, different types of cells can still exhibit different intracellular oxygen tension values in certain extracellular oxygen tensions (4% to 8%) that is not necessary to be the normoxic and hypoxic values (20.9 and 1%, respectively) conventionally recognized. The intracellular calcium tension measurement for AoSMCs show that the reoxygenation from hypoxic (<6%) to normoxic (−21%) environments triggers an increase in intracellular calcium on AoSMCs, which implies that AoSMC, instead of being solely a vessel contraction/dilation actuator, has its own standalone roles in blood pressure regulation. With the two demonstrated applications on monitoring intracellular oxygen tension and the calcium level variation, it is confirmed that the real-time imaging can be exploited to study kinetic cellular hypoxic response due to the simple design and great optical transparency of the device. With the demonstrated functionalities and capabilities, the developed method can pave a way to study the seldom explored kinetic cellular hypoxic response which is essential in various physiological and pathological conditions.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





