

Interstitial Fluid Flows along Perivascular and Adventitial Clearances around Neurovascular Bundles

Abstract
:1. Introduction
2. Results
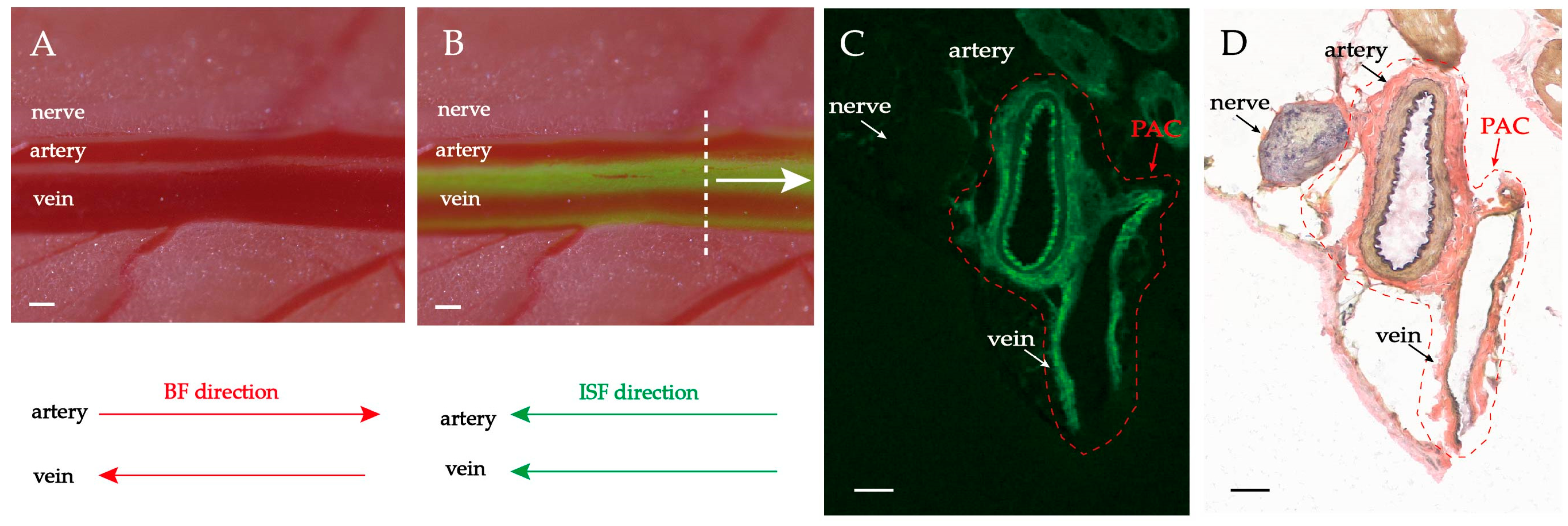
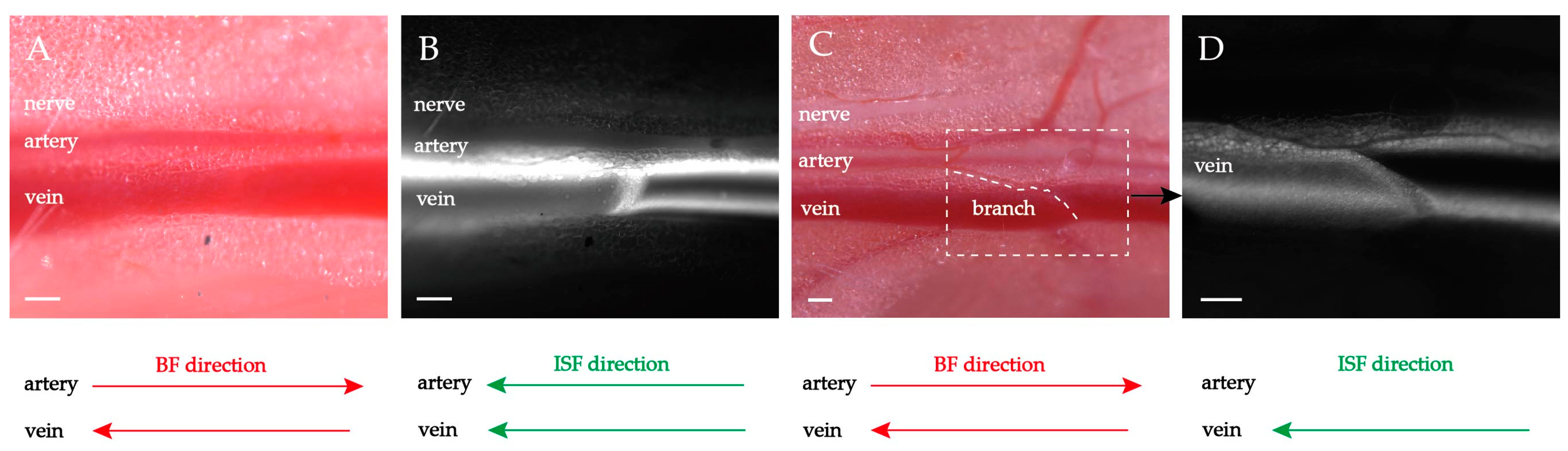
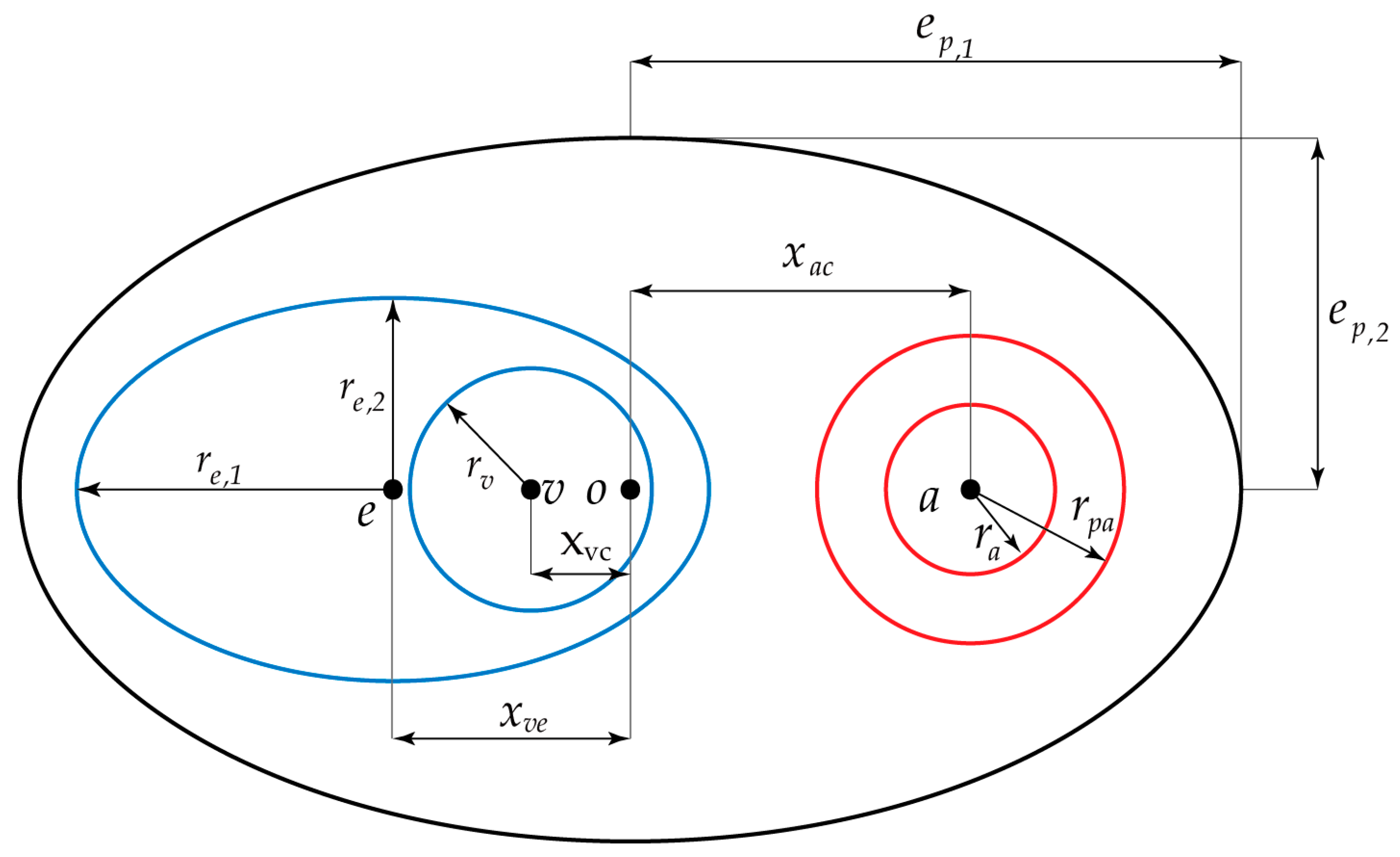
2.1. Fluorescence-Stained and Histological Analysis of the PAC Structure around Neurovascular Bundles
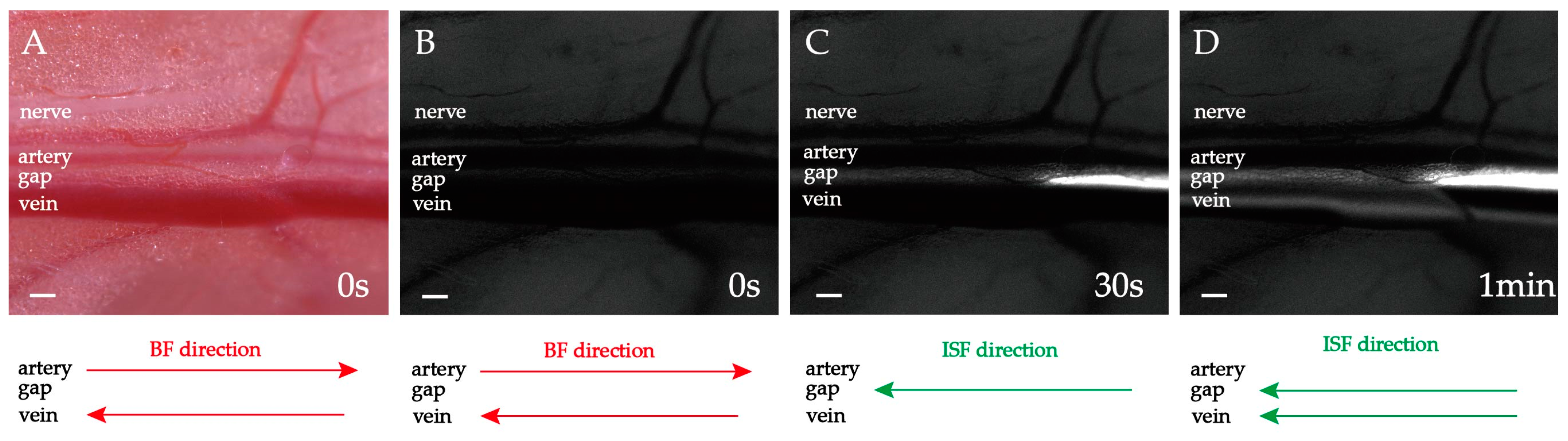
2.2. Heart-Orientated ISF Flow along PAC(p)
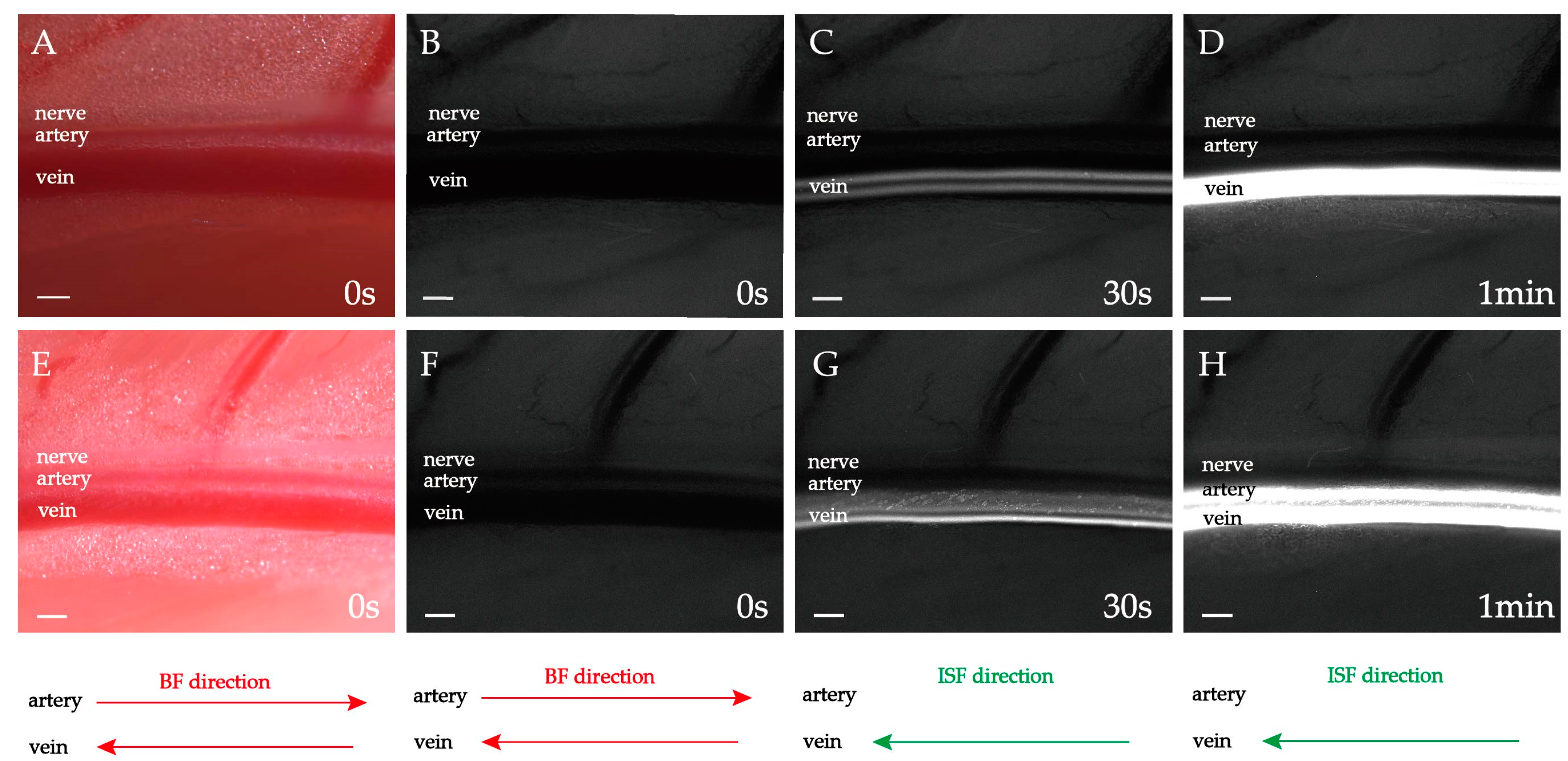
2.3. Double-Belt ISF Flow along PAC(v) around Neurovascular Bundles
2.4. Waterfall-like ISF Flow Induced by the Small Branching Vessel or Torn Fascia along PAC
3. Discussion
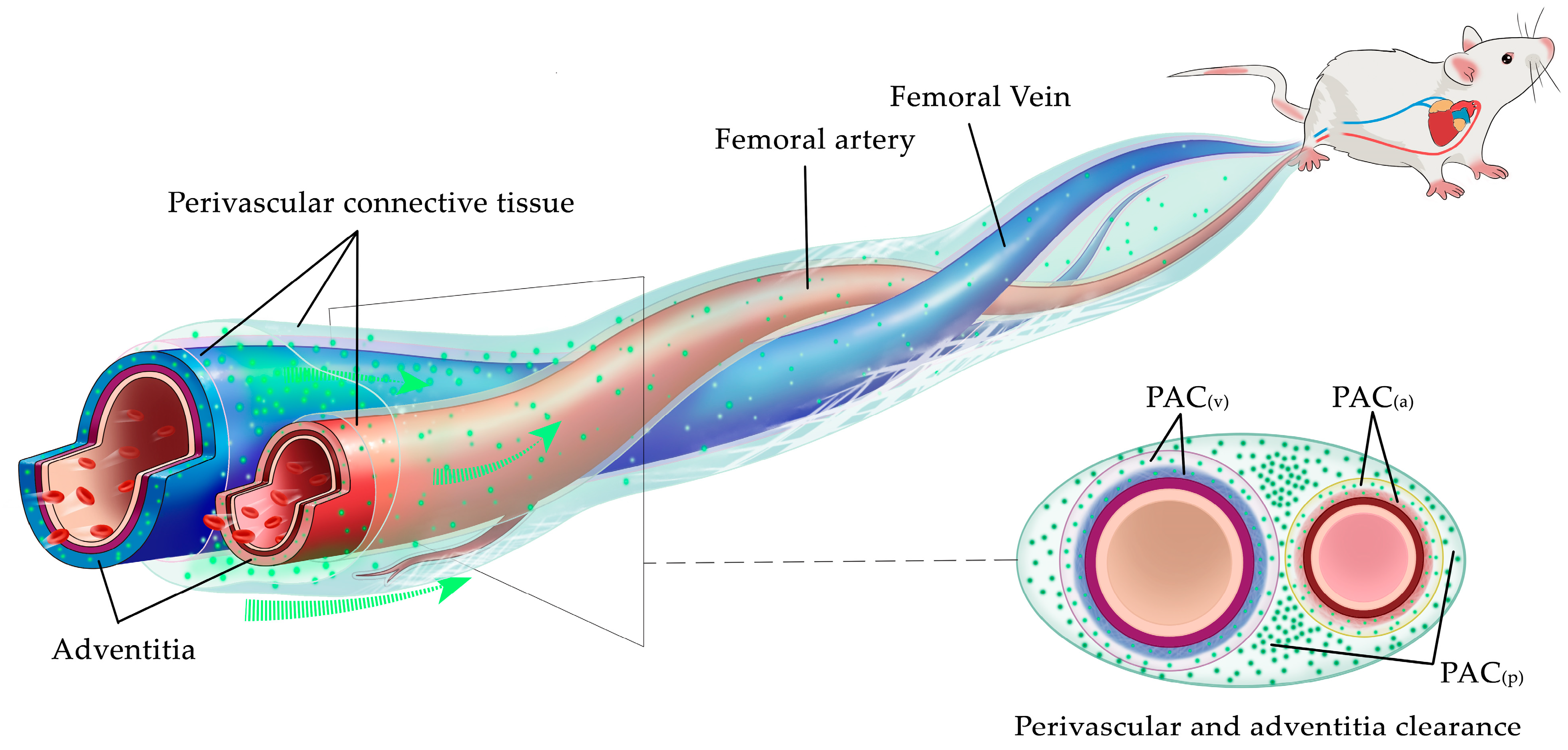
3.1. The PAC Is the Kernel Structure for ISF Flow
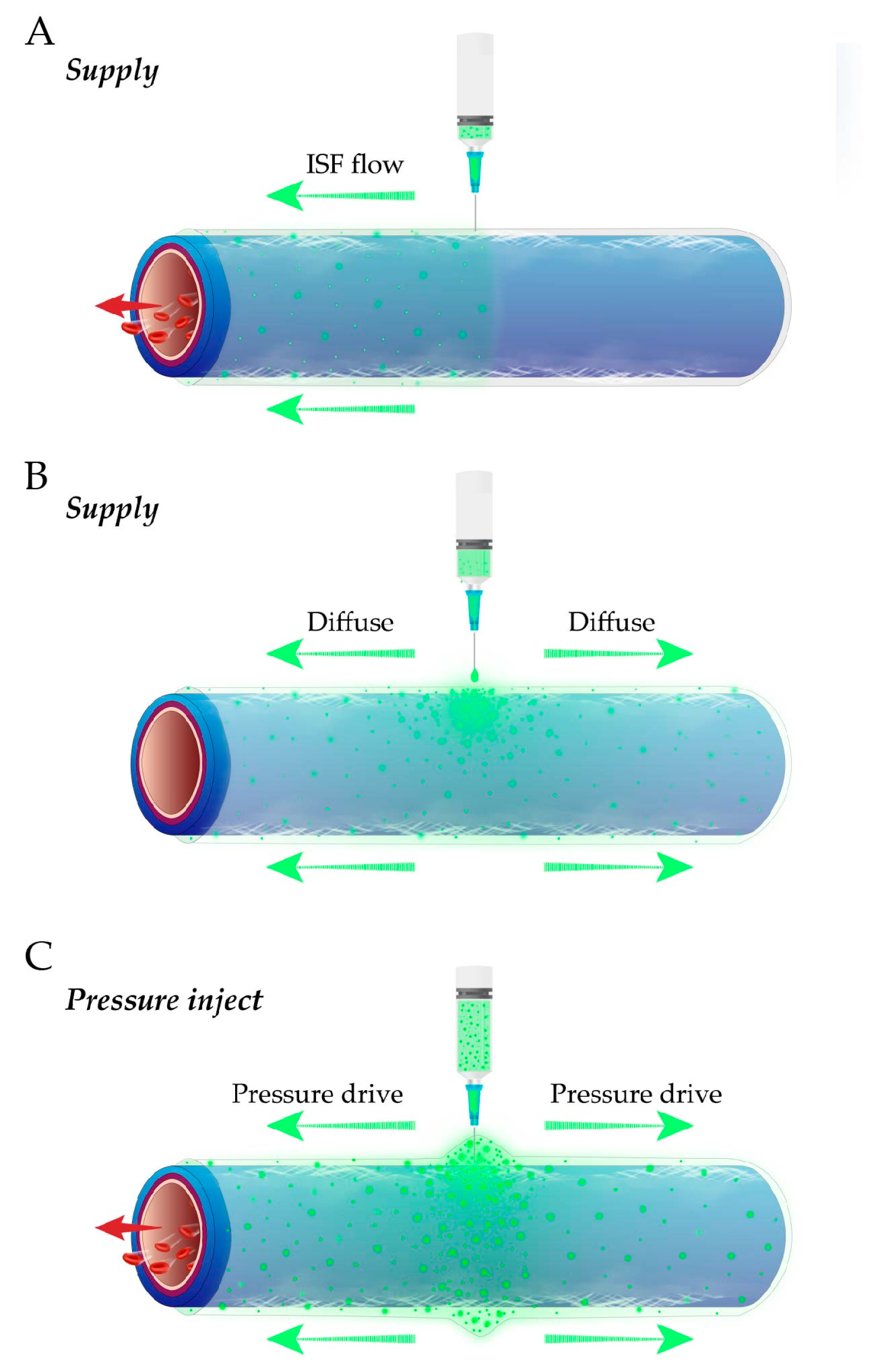
3.2. Spontaneous ISF Flow Instead of Pressure-Driven Flow or Tracer Diffusion
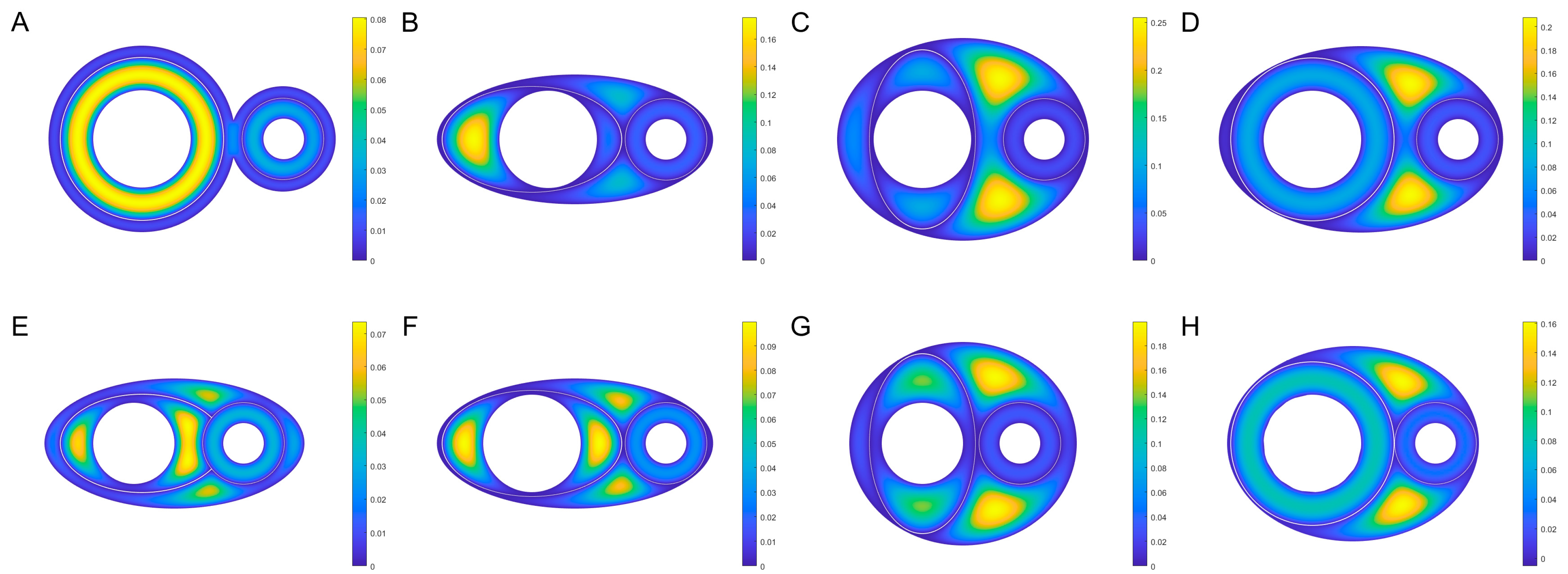
3.3. The Diversity of ISF Flow Patterns within PAC
3.4. Heart-Orientated ISF Flow
3.5. The Kinematic Laws of ISF Flow along PAC around Neurovascular Bundles
4. Materials and Methods
4.1. Experimental Animals
4.2. Surgical Operation and Fluorescent Tracing
4.3. Fluorescing Imaging
4.4. Histological Staining
4.5. Sample Size Calculation
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hall, J.E. Guyton and Hall Textbook of Medical Physiology; Saunders/Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Krueger, M.; Bechmann, I. CNS pericytes: Concepts, misconceptions, and a way out. Glia 2010, 58, 1–10. [Google Scholar] [CrossRef]
- Sepehrband, F.; Cabeen, R.P.; Choupan, J.; Barisano, G.; Law, M.; Toga, A.W.; Alzheimer’s Disease Neuroimaging Initiative. Perivascular space fluid contributes to diffusion tensor imaging changes in white matter. Neuroimage 2019, 197, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Iliff, J.J.; Wang, M.; Zeppenfeld, D.M.; Venkataraman, A.; Plog, B.A.; Liao, Y.; Deane, R.; Nedergaard, M. Cerebral arterial pulsation drives paravascular CSF-interstitial fluid exchange in the murine brain. J. Neurosci. 2013, 33, 18190–18199. [Google Scholar] [CrossRef] [Green Version]
- Iliff, J.; Wang, M.; Liao, Y.; Plogg, B.; Peng, W.; Gundersen, G.; Benveniste, H.; Vates, G.; Deane, R.; Goldman, S.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.; Yu, X.; Xu, W.; Lenahan, C.; Tu, S.; Shao, A. The role of glymphatic system in the cerebral edema formation after ischemic stroke. Exp. Neurol. 2021, 340, 113685. [Google Scholar] [CrossRef] [PubMed]
- Plog, B.; Nedergaard, M. The Glymphatic System in Central Nervous System Health and Disease: Past, Present, and Future. Annu. Rev. Pathol. 2018, 13, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Mestre, H.; Du, T.; Sweeney, A.M.; Liu, G.; Samson, A.J.; Peng, W.; Mortensen, K.N.; Staeger, F.F.; Bork, P.A.R.; Bashford, L.; et al. Cerebrospinal fluid influx drives acute ischemic tissue swelling. Science 2020, 367, eaax7171. [Google Scholar] [CrossRef]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.; Olveda, G.; Thomas, J.; Nedergaard, M.; Kelley, D. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nat. Commun. 2018, 9, 4878. [Google Scholar] [CrossRef] [Green Version]
- Kedarasetti, R.; Drew, P.; Costanzo, F. Arterial pulsations drive oscillatory flow of CSF but not directional pumping. Sci. Rep. 2020, 10, 10102. [Google Scholar] [CrossRef] [PubMed]
- Albargothy, N.; Johnston, D.; MacGregor-Sharp, M.; Weller, R.; Verma, A.; Hawkes, C.; Carare, R. Convective influx/glymphatic system: Tracers injected into the CSF enter and leave the brain along separate periarterial basement membrane pathways. Acta Neuropathol. 2018, 136, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, A.; Sharp, M.; Albargothy, N.; Fernandes, R.; Hawkes, C.; Verma, A.; Weller, R.; Carare, R. Vascular basement membranes as pathways for the passage of fluid into and out of the brain. Acta Neuropathol. 2016, 131, 725–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carare, R.; Bernardes-Silva, M.; Newman, T.; Page, A.; Nicoll, J.; Perry, V.; Weller, R. Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: Significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol. Appl. Neurobiol. 2008, 34, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Aldea, R.; Weller, R.; Wilcock, D.; Carare, R.; Richardson, G. Cerebrovascular Smooth Muscle Cells as the Drivers of Intramural Periarterial Drainage of the Brain. Front. Aging Neurosci. 2019, 11, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardlaw, J.M.; Benveniste, H.; Nedergaard, M.; Zlokovic, B.V.; Mestre, H.; Lee, H.; Doubal, F.N.; Brown, R.; Ramirez, J.; MacIntosh, B.J.; et al. Perivascular spaces in the brain: Anatomy, physiology and pathology. Nat. Rev. Neurol. 2020, 16, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yin, Y.; Yang, C.; Chen, M.; Wang, F.; Ma, C.; Li, H.; Kong, Y.; Ji, F.; Hu, J. Active interfacial dynamic transport of fluid in a network of fibrous connective tissues throughout the whole body. Cell Prolif. 2020, 53, e12760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, C.; Yin, Y.; Wang, F.; Chen, M.; Xu, L.; Wang, N.; Zhang, D.; Wang, X.; Kong, Y.; et al. An extravascular fluid transport system based on structural framework of fibrous connective tissues in human body. Cell Prolif. 2019, 52, e12667. [Google Scholar] [CrossRef] [Green Version]
- McGurk, S. Moore: Clinically Oriented Anatomy—Seventh international edition. Nurs. Stand. 2013, 28, 28. [Google Scholar] [CrossRef]
- Yin, Y.; Li, H.; Peng, G.; Yu, X.; Kong, Y. Fundamental kinematics laws of interstitial fluid flows on vascular walls. Theor. Appl. Mech. Lett. 2021, 11, 100245. [Google Scholar] [CrossRef]
- Benias, P.; Wells, R.; Sackey-Aboagye, B.; Klavan, H.; Reidy, J.; Buonocore, D.; Miranda, M.; Kornacki, S.; Wayne, M.; Carr-Locke, D.; et al. Structure and Distribution of an Unrecognized Interstitium in Human Tissues. Sci. Rep. 2018, 8, 4947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestre, H.; Mori, Y.; Nedergaard, M. The Brain’s Glymphatic System: Current Controversies. Trends Neurosci. 2020, 43, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Faghih, M.; Sharp, M. Is bulk flow plausible in perivascular, paravascular and paravenous channels? Fluids Barriers CNS 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Natali, F.; Dolce, C.; Peters, J.; Gerelli, Y.; Stelletta, C.; Leduc, G. Water dynamics in neural tissue. J. Phys. Soc. Jpn. 2013, 82 (Suppl. A), SA017. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, I.; Krizbai, I.A. In vitro models of the blood–brain barrier for the study of drug delivery to the brain. Mol. Pharm. 2014, 11, 1949–1963. [Google Scholar] [CrossRef]
- Tinajero, M.G.; Gotlieb, A.I. Recent Developments in Vascular Adventitial Pathobiology: The Dynamic Adventitia as a Complex Regulator of Vascular Disease. Am. J. Pathol. 2020, 190, 520–534. [Google Scholar] [CrossRef] [Green Version]
- Witter, K.; Tonar, Z.; Schopper, H. How many Layers has the Adventitia?—Structure of the Arterial Tunica Externa Revisited. Anat. Histol. Embryol. 2017, 46, 110–120. [Google Scholar] [CrossRef]
- Barallobre-Barreiro, J.; Loeys, B.; Mayr, M.; Rienks, M.; Verstraeten, A.; Kovacic, J. Extracellular Matrix in Vascular Disease, Part 2/4: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 2189–2203. [Google Scholar] [CrossRef] [PubMed]
- Enzmann, D.R.; Pelc, N. Normal flow patterns of intracranial and spinal cerebrospinal fluid defined with phase-contrast cine MR imaging. Radiology 1991, 178, 467–474. [Google Scholar] [CrossRef]
- Crescenzi, R.; Donahue, P.M.; Hartley, K.G.; Desai, A.A.; Scott, A.O.; Braxton, V.; Mahany, H.; Lants, D.K.; Donahue, M.J. Lymphedema evaluation using noninvasive 3T MR lymphangiography. J. Magn. Reson. Imaging 2017, 46, 1349–1360. [Google Scholar] [CrossRef] [PubMed]
- Bedussi, B.; Almasian, M.; de Vos, J.; VanBavel, E.; Bakker, E. Paravascular spacesat the brain surface: Low resistance pathways for cerebrospinal fluid flow. J. Cereb. Blood Flow Metab. 2018, 38, 719–726. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Vein | Artery | PVC | |||||
|---|---|---|---|---|---|---|---|---|
| (b) | 2.4 | 1.2 | 4.4 | 2.6 | 2.0 | 4.6 | 6.8 | 3.2 |
| (c) | 2.4 | 2.0 | 2.6 | 4.4 | 2.0 | 2.0 | 6.2 | 5.0 |
| (d) | 2.4 | 2.4 | 4.0 | 4.0 | 2.4 | 4.8 | 7.0 | 4.6 |
| (e) | 2.0 | 0.4 | 4.0 | 2.4 | 1.6 | 5.0 | 6.4 | 3.2 |
| (f) | 2.4 | 2.0 | 4.4 | 2.6 | 2.0 | 4.6 | 6.8 | 3.2 |
| (g) | 2.0 | 2.0 | 2.6 | 4.4 | 2.0 | 4.8 | 5.6 | 5.0 |
| (h) | 2.4 | 2.0 | 4.0 | 4.0 | 2.0 | 4.0 | 6.2 | 4.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Y.; Yu, X.; Peng, G.; Wang, F.; Yin, Y. Interstitial Fluid Flows along Perivascular and Adventitial Clearances around Neurovascular Bundles. J. Funct. Biomater. 2022, 13, 172. https://doi.org/10.3390/jfb13040172
Kong Y, Yu X, Peng G, Wang F, Yin Y. Interstitial Fluid Flows along Perivascular and Adventitial Clearances around Neurovascular Bundles. Journal of Functional Biomaterials. 2022; 13(4):172. https://doi.org/10.3390/jfb13040172
Chicago/Turabian StyleKong, Yiya, Xiaobin Yu, Gang Peng, Fang Wang, and Yajun Yin. 2022. "Interstitial Fluid Flows along Perivascular and Adventitial Clearances around Neurovascular Bundles" Journal of Functional Biomaterials 13, no. 4: 172. https://doi.org/10.3390/jfb13040172