
Bioremediation of Multiple Heavy Metals Mediated by Antarctic Marine Isolated Dietzia psychralcaliphila JI1D

 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media and Chemicals
2.2. Dietzia Psychralcaliphila JI1D Culture Condition
2.3. Determination of Maximum Tolerance Concentration (MTC)
2.4. Experimental Setup for Growth with Heavy Metals
2.5. Heavy Metal Analysis by ICP-MS
2.6. Genomic Analysis for Heavy Metal Resistance
3. Results and Discussion
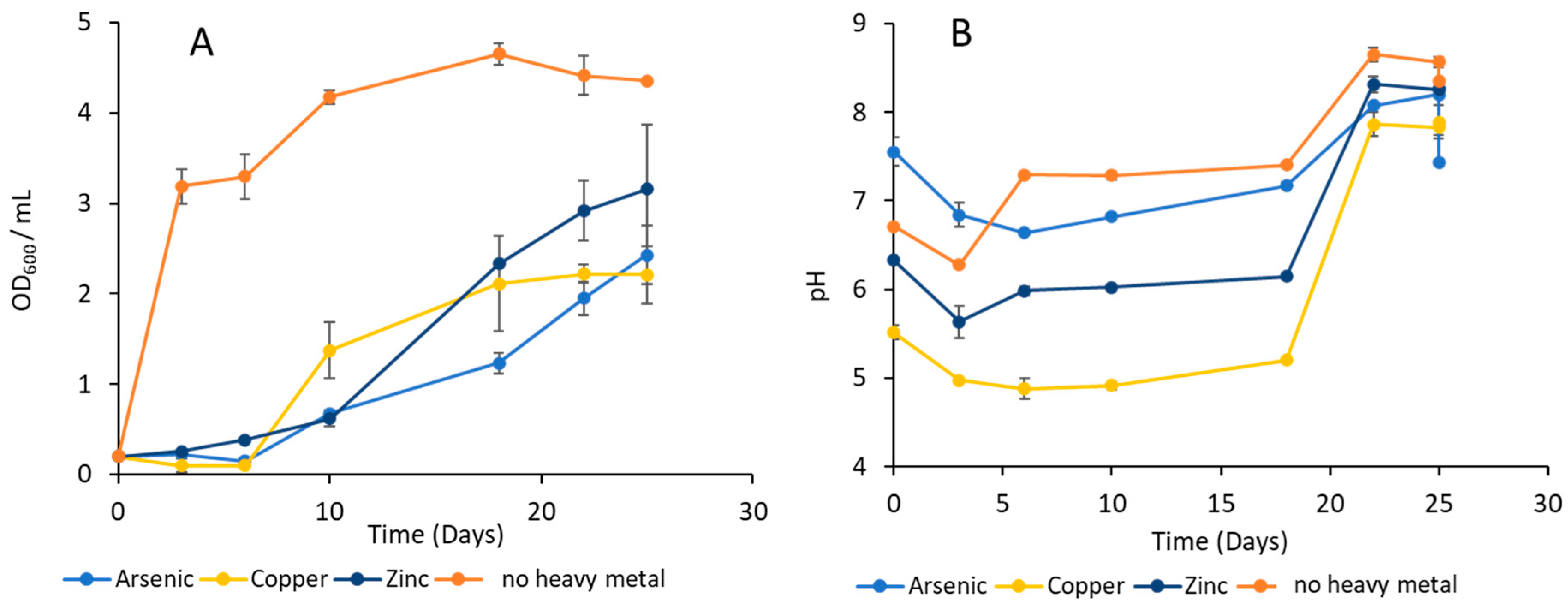
3.1. Measurement of MTC with D. psychralcaliphila JI1D
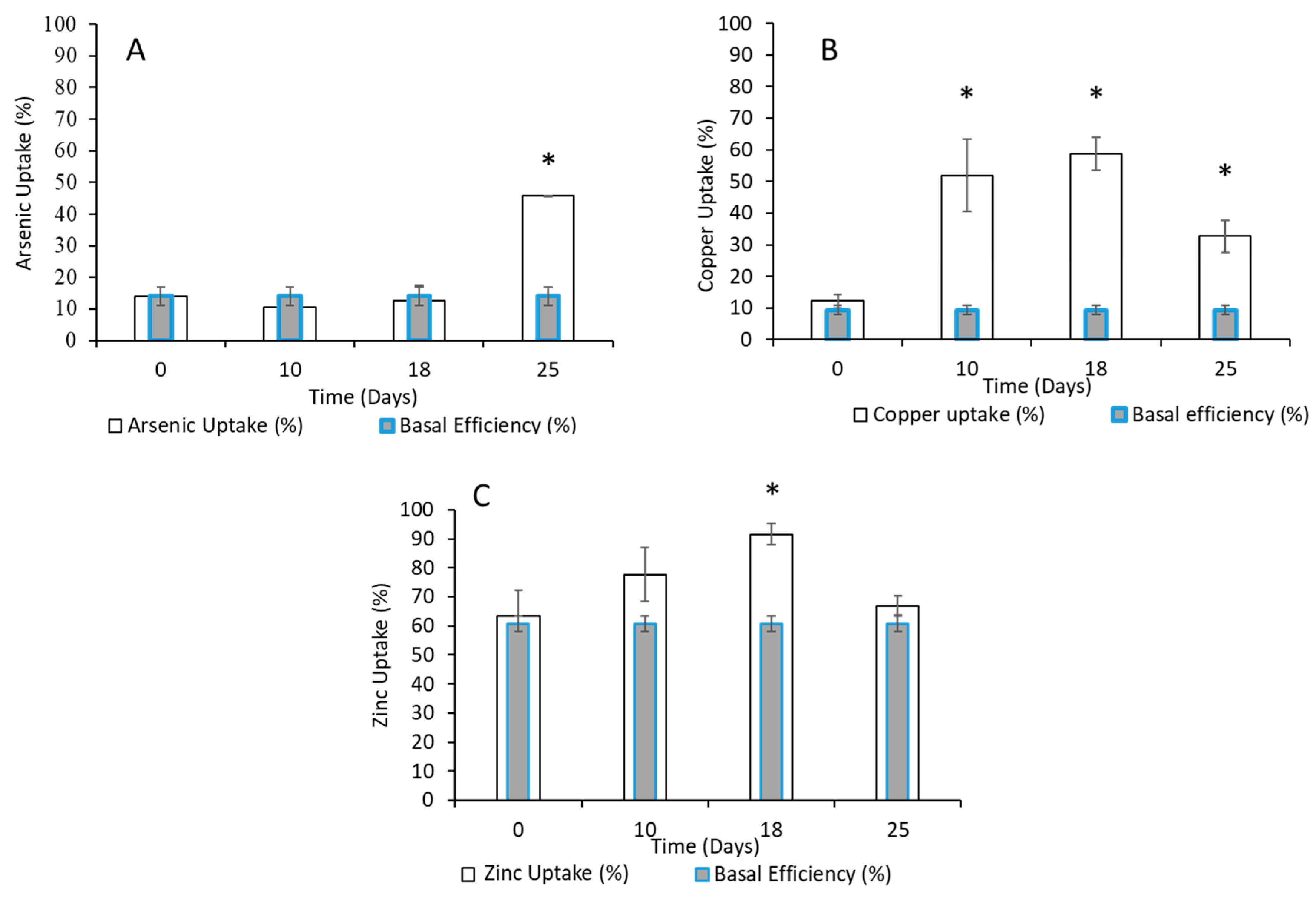
3.2. Quantification of Heavy Metals by ICP-MS
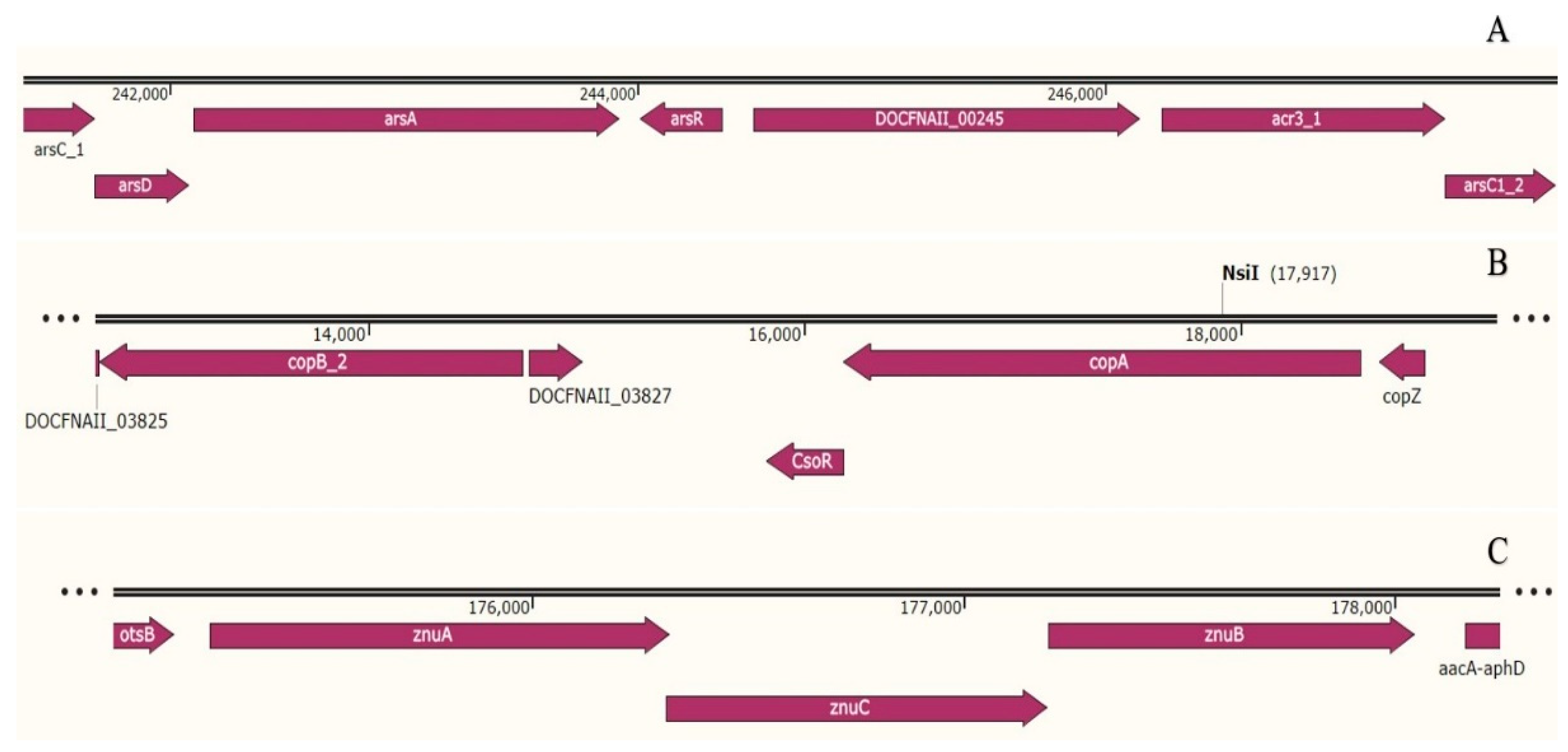
3.3. Genomic Insights of Heavy Metal Tolerance and Stress Response
3.3.1. Arsenic Resistance Determinants
3.3.2. Copper Resistant Determinants
3.3.3. Zinc Resistant Determinants
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayangbenro, A.S.; Babalola, O.O. A New Strategy for Heavy Metal Polluted Environments: A Review of Microbial Biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Photolo, M.; Sitole, L.; Mavumengwana, V.; Tlou, M. Genomic and Physiological Investigation of Heavy Metal Resistance from Plant Endophytic Methylobacterium radiotolerans MAMP 4754, Isolated from Combretum erythrophyllum. Int. J. Environ. Res. Public Health 2021, 18, 997. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.M.; Rink, L.; Haase, H. The Essential Toxin: Impact of Zinc on Human Health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarekegn, M.M.; Salilih, F.Z.; Ishetu, A.I. Microbes Used as a Tool for Bioremediation of Heavy Metal from the Environment. Cogent Food Agric. 2020, 6, 1783174. [Google Scholar] [CrossRef]
- Batey, T.; Berryman, C.; Line, C. The Disposal of Copper-Enriched Pig-Manure Slurry on Grassland. Grass Forage Sci. 2006, 27, 139–144. [Google Scholar] [CrossRef]
- Chung, J.-Y.; Yu, S.-D.; Hong, Y.-S. Environmental Source of Arsenic Exposure. J. Prev. Med. Public Health 2014, 47, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Soto, J.; Charles, T.; Lynch, M.; Larama, G.; Herrera, H.; Arriagada, C. Genome Sequence of Brevundimonas sp., an Arsenic Resistant Soil Bacterium. Diversity 2021, 13, 344. [Google Scholar] [CrossRef]
- Kapahi, M.; Sachdeva, S. Bioremediation Options for Heavy Metal Pollution. J. Health Pollut. 2019, 9, 191203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abioye, O.P. Biological Remediation of Hydrocarbon and Heavy Metals Contaminated Soil. In Soil Contamination; Pascucci, S., Ed.; IntechOpen: Rijeka, Croatia, 2011. [Google Scholar]
- Gambino, E.; Chandrasekhar, K.; Nastro, R.A. SMFC as a tool for the removal of hydrocarbons and metals in the marine environment: A concise research update. Environ. Sci. Pollut. Res. 2021, 28, 30436–30451. [Google Scholar] [CrossRef]
- Arora, V.; Khosla, B. Conventional and Contemporary Techniques for Removal of Heavy Metals from Soil. In Biodegradation Technology of Organic and Inorganic Pollutants; Mendes, K.F., de Sousa, R.N., Mielke, K.C., Eds.; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar]
- Mosa, K.A.; Saadoun, I.; Kumar, K.; Helmy, M.; Dhankher, O.P. Potential Biotechnological Strategies for the Cleanup of Heavy Metals and Metalloids. Front. Plant Sci. 2016, 7, 303. [Google Scholar] [CrossRef]
- Perelo, L.W. Review: In situ and bioremediation of organic pollutants in aquatic sediments. J. Hazard. Mater. 2010, 177, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuppusamy, S.; Thavamani, P.; Venkateswarlu, K.; Lee, Y.B.; Naidu, R.; Megharaj, M. Remediation Approaches for Polycyclic Aromatic Hydrocarbons (PAHs) Contaminated Soils: Technological Constraints, Emerging Trends and Future Directions. Chemosphere 2017, 168, 944–968. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; Macaya, C.; Bravo, G.; Dorochesi, F.; Cumsille, A.; Valencia, R.; Rojas, C.; Seeger, M. Living at the Frontiers of Life: Extremophiles in Chile and Their Potential for Bioremediation. Front. Microbiol. 2018, 9, 2309. [Google Scholar] [CrossRef] [PubMed]
- Giudice, A.L.; Caruso, G.; Rizzo, C.; Papale, M.; Azzaro, M. Bacterial communities versus anthropogenic disturbances in the Antarctic coastal marine environment. Environ. Sustain. 2019, 2, 297–310. [Google Scholar] [CrossRef]
- Alvarez, L.M.; Bolhuis, H.; Mau, G.K.; Kok-Gan, C.; Sing, C.C.; Mac Cormack, W.; Ruberto, L. Identification of key bacterial players during successful full-scale soil field bioremediation in Antarctica. Int. Biodeterior. Biodegradation 2022, 168, 105354. [Google Scholar] [CrossRef]
- Darham, S.; Syed-Muhaimin, S.N.; Subramaniam, K.; Zulkharnain, A.; Shaharuddin, N.A.; Khalil, K.A.; Ahmad, S.A. Optimisation of Various Physicochemical Variables Affecting Molybdenum Bioremediation Using Antarctic Bacterium, Arthrobacter sp. Strain AQ5-05. Water 2021, 13, 2367. [Google Scholar] [CrossRef]
- Michaud, L.; Di Marco, G.; Bruni, V.; Giudice, A.L. Biodegradative potential and characterization of psychrotolerant polychlorinated biphenyl-degrading marine bacteria isolated from a coastal station in the Terra Nova Bay (Ross Sea, Antarctica). Mar. Pollut. Bull. 2007, 54, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Barrios, R.; Rubiano-Labrador, C.; Navarro-Narvaez, D.; Escobar-Galarza, J.; González, D.; Mira, S.; Moreno, D.; Contreras, A.; Miranda-Castro, W. Perchlorate-reducing bacteria from Antarctic marine sediments. Environ. Monit. Assess. 2022, 194, 654. [Google Scholar] [CrossRef] [PubMed]
- Ausuri, J.; Vitale, G.; Coppola, D.; Esposito, F.P.; Buonocore, C.; de Pascale, D. Assessment of the Degradation Potential and Genomic Insights towards Phenanthrene by Dietzia psychralcaliphila JI1D. Microorganisms 2021, 9, 1327. [Google Scholar] [CrossRef] [PubMed]
- Centurion, V.; Silva, J.; Duarte, A.; Rosa, L.; Oliveira, V. Comparing resistome profiles from anthropogenically impacted and non-impacted areas of two South Shetland Islands—Maritime Antarctica. Environ. Pollut. 2022, 304, 119219. [Google Scholar] [CrossRef] [PubMed]
- Centurion, V.; Delforno, T.; Lacerda-Júnior, G.; Duarte, A.; Silva, L.; Bellini, G.; Rosa, L.; Oliveira, V. Unveiling resistome profiles in the sediments of an Antarctic volcanic island. Environ. Pollut. 2019, 255, 113240. [Google Scholar] [CrossRef] [PubMed]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Molecular, Clinical and Environmental Toxicicology Volume 3: Environmental Toxicology. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar] [CrossRef]
- Lambert, M.; Leven, B.A.; Green, R.M. New Methods of Cleaning Up Heavy Metal in Soils and Water Innovative Solutions to an Environmental Problem. Environ. Sci. Technol. Briefs Citiz. 2000, 7, 1–3. [Google Scholar]
- Kuznetsova, O.V.; Khlebtsov, N.G.; Jarosz, M.; Timerbaev, A.R. Metal-Specific Response of High-Resolution ICP-MS for Proteins Binding to Gold Nanoparticles in Human Serum. Anal. Chem. 2021, 93, 14918–14922. [Google Scholar] [CrossRef] [PubMed]
- EPA. U.S. Method 6020B Inductively Coupled Plasma-Mass Spectrometry; EPA: Washington, DC, USA, 2014; pp. 1–33. [Google Scholar]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; De Jongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35 (Suppl. 2), W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, N.; Bhatt, R. Arsenic Resistance and Accumulation by Two Bacteria Isolated from a Natural Arsenic Contaminated Site. J. Basic Microbiol. 2015, 55, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Gikas, P.; Sengör, S.S.; Ginn, T.; Moberly, J.; Peyton, B. The Effects of Heavy Metals and Temperature on Microbial Growth and Lag. Glob. Nest J. 2009, 11, 325–332. [Google Scholar] [CrossRef]
- Abbas, S.Z.; Riaz, M.; Ramzan, N.; Zahid, M.T.; Shakoori, F.R.; Rafatullah, M. Isolation and Characterization of Arsenic Resistant Bacteria from Wastewater. Braz. J. Microbiol. 2014, 45, 1309–1315. [Google Scholar] [CrossRef] [Green Version]
- Prosun, T.A.; Younus, A.; Monira, Y.; Brun, M.S.; Badiuzzaman, K.; Harun-Or, R. Identification and Evaluation of Arsenic Tolerant Bacteria for Arsenic Mitigation in Contaminated Soil. J. Bacteriol. Mycol. 2020, 7, 1156. [Google Scholar]
- Porcheron, G.; Garénaux, A.; Proulx, J.; Sabri, M.; Dozois, C.M. Iron, Copper, Zinc, and Manganese Transport and Regulation in Pathogenic Enterobacteria: Correlations between Strains, Site of Infection and the Relative Importance of the Different Metal Transport Systems for Virulence. Front. Cell. Infect. Microbiol. 2013, 3, 90. [Google Scholar] [CrossRef] [Green Version]
- Posacka, A.M.; Semeniuk, D.M.; Maldonado, M.T. Effects of Copper Availability on the Physiology of Marine Heterotrophic Bacteria. Front. Mar. Sci. 2019, 5, 523. [Google Scholar] [CrossRef]
- Romaniuk, K.; Ciok, A.; Decewicz, P.; Uhrynowski, W.; Budzik, K.; Nieckarz, M.; Pawlowska, J.; Zdanowski, M.K.; Bartosik, D.; Dziewit, L. Insight into heavy metal resistome of soil psychrotolerant bacteria originating from King George Island (Antarctica). Polar Biol. 2018, 41, 1319–1333. [Google Scholar] [CrossRef] [Green Version]
- Gholami, M.; Etemadifar, Z. Isolation and characterization of a novel strain of genus Dietzia capable of multiple-extreme resistance. Microbiology 2015, 84, 389–397. [Google Scholar] [CrossRef]
- Máthé, I.; Benedek, T.; Táncsics, A.; Palatinszky, M.; Lányi, S.; Márialigeti, K. Diversity, activity, antibiotic and heavy metal resistance of bacteria from petroleum hydrocarbon contaminated soils located in Harghita County (Romania). Int. Biodeterior. Biodegrad. 2012, 73, 41–49. [Google Scholar] [CrossRef]
- Freitas, D.; Lima-Bittencourt, C.; Reis, M.; Costa, P.; Assis, P.; Chartone-Souza, E.; Nascimento, A. Molecular characterization of early colonizer bacteria from wastes in a steel plant. Lett. Appl. Microbiol. 2008, 47, 241–249. [Google Scholar] [CrossRef]
- Gusain, P.; Paliwal, R.; Singh, V. Rhizoremediation of cadmium-contaminated soil associated with hydroxamate siderophores isolated from Cd-resistant plant growth–promoting Dietzia maris and Lysinibacillus strains. Int. J. Phytoremediation 2017, 19, 290–299. [Google Scholar] [CrossRef]
- Morton-Bermea, O.; Álvarez, E.H.; Gaso, I.; Segovia, N. Heavy Metal Concentrations in Surface Soils from Mexico City. Bull. Environ. Contam. Toxicol. 2002, 68, 0383–0388. [Google Scholar] [CrossRef] [PubMed]
- Galiulin, R.V.; Galiulina, R.A. Removing heavy metals from soil with plants. Her. Russ. Acad. Sci. 2008, 78, 141–143. [Google Scholar] [CrossRef]
- Rodríguez-Tirado, V.; Green-Ruiz, C.; Gómez-Gil, B. Cu and Pb biosorption on Bacillus thioparans strain U3 in aqueous solution: Kinetic and equilibrium studies. Chem. Eng. J. 2012, 181–182, 352–359. [Google Scholar] [CrossRef]
- Wierzba, S. Biosorption of lead(II), zinc(II) and nickel(II) from industrial wastewater by Stenotrophomonas maltophilia and Bacillus subtilis. Pol. J. Chem. Technol. 2015, 17, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Babu, A.G.; Shea, P.J.; Sudhakar, D.; Jung, I.-B.; Oh, B.-T. Potential use of Pseudomonas koreensis AGB-1 in association with Miscanthus sinensis to remediate heavy metal(loid)-contaminated mining site soil. J. Environ. Manag. 2015, 151, 160–166. [Google Scholar] [CrossRef]
- Diep, P.; Mahadevan, R.; Yakunin, A.F. Heavy Metal Removal by Bioaccumulation Using Genetically Engineered Microorganisms. Front. Bioeng. Biotechnol. 2018, 6, 157. [Google Scholar] [CrossRef] [Green Version]
- Slyemi, D.; Bonnefoy, V. How prokaryotes deal with arsenic†. Environ. Microbiol. Rep. 2012, 4, 571–586. [Google Scholar] [CrossRef]
- Kotoky, R.; Pandey, P. The genomic attributes of Cd-resistant, hydrocarbonoclastic Bacillus subtilis SR1 for rhizodegradation of benzo(a)pyrene under co-contaminated conditions. Genomics 2020, 113, 613–623. [Google Scholar] [CrossRef]
- Rawle, R.; Saley, T.C.; Kang, Y.-S.; Wang, Q.; Walk, S.; Bothner, B.; McDermott, T.R. Introducing the ArsR-Regulated Arsenic Stimulon. Front. Microbiol. 2021, 12, 630562. [Google Scholar] [CrossRef]
- Lin, Y.-F.; Walmsley, A.R.; Rosen, B.P. An arsenic metallochaperone for an arsenic detoxification pump. Proc. Natl. Acad. Sci. USA 2006, 103, 15617–15622. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, M.; Kumar, V.; Sharma, D. Multimetal tolerance mechanisms in bacteria: The resistance strategies acquired by bacteria that can be exploited to ‘clean-up’ heavy metal contaminants from water. Aquat. Toxicol. 2019, 212, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Procópio, L.; Alvarez, V.M.; Jurelevicius, D.A.; Hansen, L.; Sørensen, S.J.; Cardoso, J.S.; Pádula, M.; Leitão, Á.C.; Seldin, L.; Van Elsas, J.D. Insight from the Draft Genome of Dietzia Cinnamea P4 Reveals Mechanisms of Survival in Complex Tropical Soil Habitats and Biotechnology Potential. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2012, 101, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Yang, Z.; Wu, X.; Wang, J.; Du, D.; Cai, J.; Lv, K.; Chen, H.; Mei, J.; Chen, M.; et al. Sediment biomarker, bacterial community characterization of high arsenic aquifers in Jianghan Plain, China. Sci. Rep. 2017, 7, srep42037. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Mou, T.J.; Chakrabarty, R.P.; Hoque, S.; Hossain, M.A.; Sultana, M. Diversity of arsenite oxidase gene and arsenotrophic bacteria in arsenic affected Bangladesh soils. AMB Express 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thummeepak, R.; Pooalai, R.; Harrison, C.; Gannon, L.; Thanwisai, A.; Chantratita, N.; Millard, A.D.; Sitthisak, S. Essential Gene Clusters Involved in Copper Tolerance Identified in Acinetobacter baumannii Clinical and Environmental Isolates. Pathogens 2020, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Chang, F.M.J.; Coyne, H.J.; Cubillas, C.; Vinuesa, P.; Fang, X.; Ma, Z.; Ma, D.; Helmann, J.D.; García-De Los Santos, A.; Wang, Y.X.; et al. Cu(I)-mediated Allosteric Switching in a Copper-sensing Operon Repressor (CsoR). J. Biol. Chem. 2014, 289, 19204–19217. [Google Scholar] [CrossRef] [Green Version]
- Völlmecke, C.; Drees, S.L.; Reimann, J.; Albers, S.-V.; Lübben, M. The ATPases CopA and CopB both contribute to copper resistance of the thermoacidophilic archaeon Sulfolobus solfataricus. Microbiology 2012, 158, 1622–1633. [Google Scholar] [CrossRef]
- Zhang, L.; Koay, M.; Maher, M.J.; Xiao, Z.; Wedd, A.G. Intermolecular Transfer of Copper Ions from the CopC Protein of Pseudomonas syringae. Crystal Structures of Fully Loaded CuICuII Forms. J. Am. Chem. Soc. 2006, 128, 5834–5850. [Google Scholar] [CrossRef]
- Li, L.-G.; Cai, L.; Zhang, X.-X.; Zhang, T. Potentially novel copper resistance genes in copper-enriched activated sludge revealed by metagenomic analysis. Appl. Microbiol. Biotechnol. 2014, 98, 10255–10266. [Google Scholar] [CrossRef]
- Xie, J.; Gu, J.; Wang, X.; Hu, T.; Sun, W.; Lei, L.; Zhang, R.; Guo, H. Insights into the beneficial effects of woody peat for reducing abundances of antibiotic resistance genes during composting. Bioresour. Technol. 2021, 342, 125903. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Nojima, Y.; Sakamoto, T.; Iwabuchi, K.; Nakazato, T.; Bono, H.; Toyoda, A.; Fujiyama, A.; Kanost, M.; Tabunoki, H. Comparative analysis of seven types of superoxide dismutases for their ability to respond to oxidative stress in Bombyx mori. Sci. Rep. 2019, 9, 2170. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ely, R.L. Genome-Wide Transcriptional Responses of Nitrosomonas Europaea to Zinc. Arch. Microbiol. 2008, 189, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Patzer, S.I.; Hantke, K. The Zinc-responsive Regulator Zur and Its Control of theznu Gene Cluster Encoding the ZnuABC Zinc Uptake System in Escherichia coli. J. Biol. Chem. 2000, 275, 24321–24332. [Google Scholar] [CrossRef] [Green Version]
- Anton, A.; Große, C.; Reißmann, J.; Pribyl, T.; Nies, D.H. CzcD Is a Heavy Metal Ion Transporter Involved in Regulation of Heavy Metal Resistance in Ralstonia sp. Strain CH34. J. Bacteriol. 1999, 181, 6876–6881. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Description | Accession Number |
|---|---|---|---|
| DOCFNAII_00244 | arsR | arsenate/arsenite-responsive transcriptional repressor | P37309 |
| DOCFNAII_00242 | arsD | arsenical resistance operon trans-acting repressor | O52028 |
| DOCFNAII_00243 | arsA | transmembrane arsenical efflux pump | P52145 |
| DOCFNAII_00246 | arsB | arsenite transporter | P0AB93 |
| DOCFNAII_00247 | arsC | arsenate reductase | P0A006 |
| DOCFNAII_00656 | glpF | glycerol uptake facilitator protein | Q51389 |
| DOCFNAII_03828 DOCFNAII_00088 | csoR | copper sensing transcriptional repressor | P9WP49 |
| DOCFNAII-03829 DOCFNAII_00090 | copA | CadA-like heavy metal translocating P-type ATP-ase | O32220 |
| DOCFNAII_03960 DOCFNAII_02731 DOCFNAII_03826 | copB | P-type Cu2+ transporter | Q59385 |
| DOCFNAII_03084 DOCFNAII_01147 | copC | copper resistance protein C | P12376 |
| DOCFNAII_00341 | copZ | periplasmic copper chaperone | O32221 |
| DOCFNAII_03834 DOCFNAII_03956 | mmco | multicopper oxidase | I6WZK7 |
| DOCFNAII_00093 | sod1 | superoxide dismutase belonging to Cu-Zn family | P53649 |
| DOCFNAII_03463 | znt A | zinc efflux system | Q8NEW0 |
| DOCFNAII_02177 | znuA | ABC zinc/manganese transport system (substrate-binding protein) | O34966 |
| DOCFNAII_02178 | znuB | ABC zinc/manganese transport system ATP-binding protein | P39832 |
| DOCFNAII_02179 | znuC | ABC zinc/manganese transport system permease protein | P0A9X1 |
| DOCFNAII_02964 | zur | metalloregulatory protein family | P54479 |
| DOCFNAII_01529 DOCFNAII_03837 | czcD | cobalt/zinc/cadmium efflux system components | P13512 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ausuri, J.; Dell’Anno, F.; Vitale, G.A.; Palma Esposito, F.; Funari, V.; Franci, G.; Galdiero, M.; Della Sala, G.; Tedesco, P.; Coppola, D.; et al. Bioremediation of Multiple Heavy Metals Mediated by Antarctic Marine Isolated Dietzia psychralcaliphila JI1D. J. Mar. Sci. Eng. 2022, 10, 1669. https://doi.org/10.3390/jmse10111669
Ausuri J, Dell’Anno F, Vitale GA, Palma Esposito F, Funari V, Franci G, Galdiero M, Della Sala G, Tedesco P, Coppola D, et al. Bioremediation of Multiple Heavy Metals Mediated by Antarctic Marine Isolated Dietzia psychralcaliphila JI1D. Journal of Marine Science and Engineering. 2022; 10(11):1669. https://doi.org/10.3390/jmse10111669
Chicago/Turabian StyleAusuri, Janardan, Filippo Dell’Anno, Giovanni Andrea Vitale, Fortunato Palma Esposito, Valerio Funari, Gianluigi Franci, Massimiliano Galdiero, Gerardo Della Sala, Pietro Tedesco, Daniela Coppola, and et al. 2022. "Bioremediation of Multiple Heavy Metals Mediated by Antarctic Marine Isolated Dietzia psychralcaliphila JI1D" Journal of Marine Science and Engineering 10, no. 11: 1669. https://doi.org/10.3390/jmse10111669