
Dental Morphology, Palaeoecology and Palaeobiogeographic Significance of a New Species of Requiem Shark (Genus Carcharhinus) from the Lower Miocene of Peru (East Pisco Basin, Chilcatay Formation)

 , , and
, , and 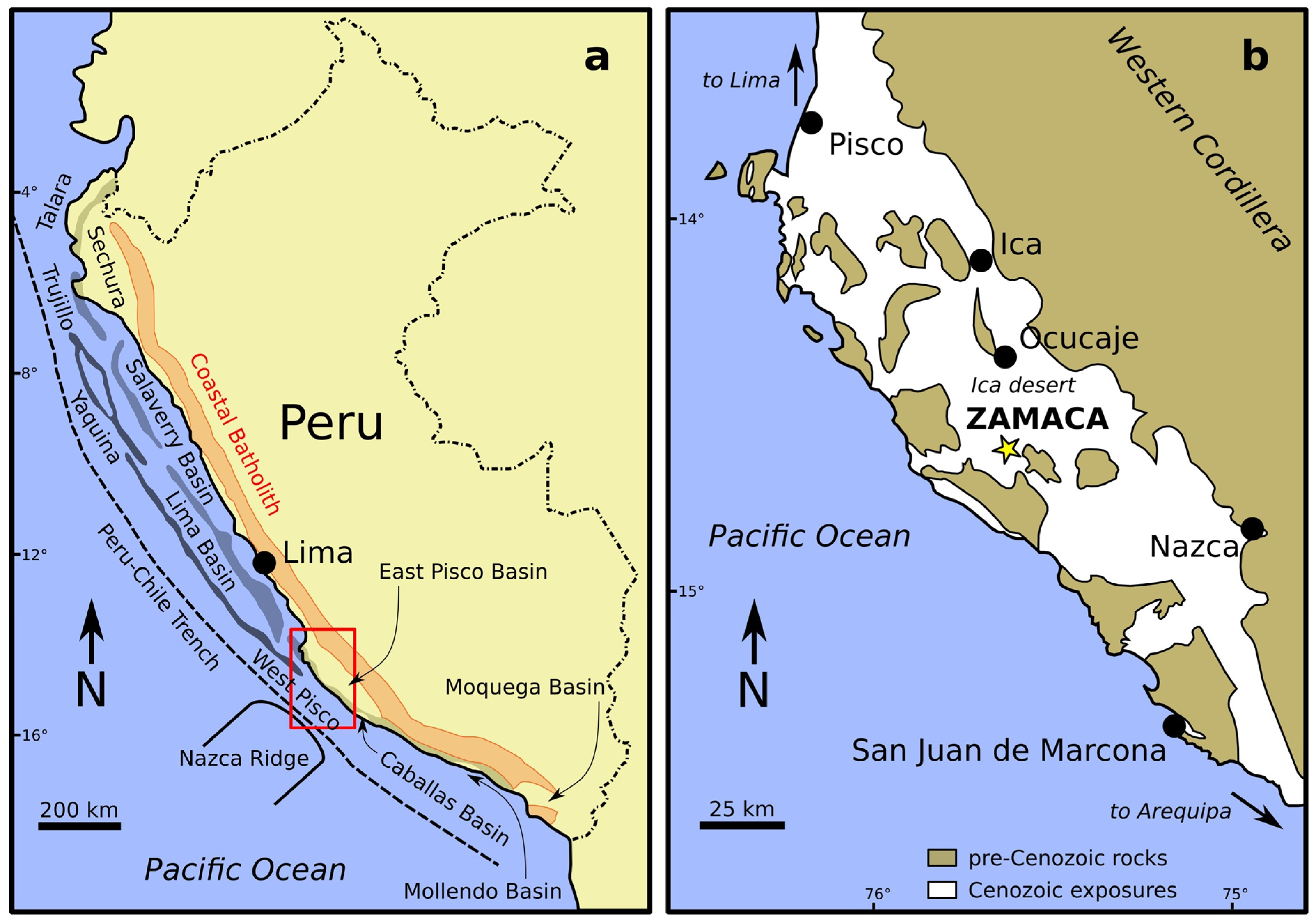{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Abbreviations
2.2. Nomenclatural Acts
2.3. (Allo)Stratigraphic Context
2.4. Palaeontological Background
2.5. Geochronological Framework
3. Results
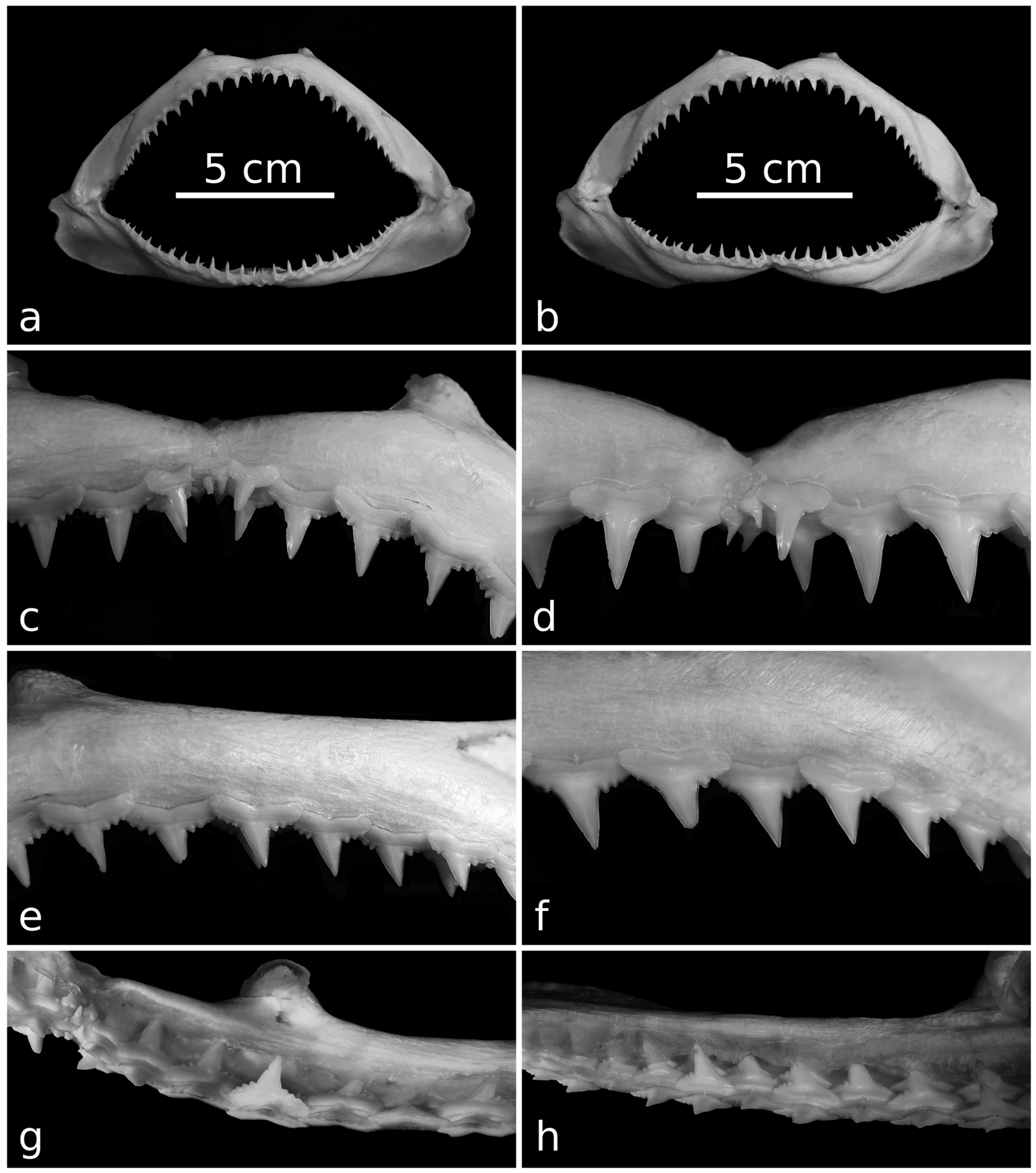
- (1)
- The cusp is transversely narrower and, especially, more narrowly based.
- (2)
- The cusp exhibits a sigmoid profile (which is only incipient in C. macloti) and a distinct coronal twist.
- (3)
- The labial crown face is weakly convex transversely (rather than almost flat as in C. macloti).
- (4)
- In labial/lingual view, the cusp of the lateral teeth is characteristically “hook-shaped”, with the crown apex bending distinctly toward the commissure. This is due to the mesial and distal cutting edges of the cusp becoming progressively more convex and concave, respectively.
- (5)
- The cusp is labiolingually thick at the base and semicylindrical, whereas the distal and mesial heels are remarkably thinner. In apical view, the cusp is clearly divided from the distal heel by means of an abrupt variation of the labiolingual thickness of the crown. In C. macloti, the cusp is proportionally thinner at the base and not semicylindrical, and the thickness of the crown decreases regularly towards its distobasal end.
- (1)
- The mesial and distal cutting edges of the cusp are completely smooth (rather than being finely serrated to approximately their mid-points);
- (2)
- The distal heel bears one to five, coarse, angularly lobate serrae that become more prominent toward the cusp (rather than finer, more abundant serrations).
4. Discussion
4.1. Palaeoecology
4.2. Palaeobiogeography
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- White, W.T.; Kyne, P.M.; Harris, M. Lost before found: A new species of whaler shark Carcharhinus obsolerus from the Western Central Pacific known only from historic records. PLoS ONE 2019, 14, e0209387. [Google Scholar] [CrossRef] [PubMed]
- Compagno, L.J.V. FAO species catalogue. Sharks of the world. In An Annotated and Illustrated Catalogue of Shark Species Known to Date. Part 1. Hexanchiformes to Lamniformes; FAO: Rome, Italy, 1984; Volume 4, pp. 1–258. [Google Scholar]
- Garrick, J.A.F. Sharks of the Genus Carcharhinus; NOAA Technical Report NMFS Circular; NOAA Fisheries: Silver Spring, MD, USA, 1982; Volume 405, pp. 1–194. [Google Scholar]
- Garrick, J.A.F. Additions to a Revision of the Shark Genus Carcharhinus: Synonymy of Aprionodon and Hypoprion, and Description of a New Species of Carcharhinus (Carcharhinidae); NOAA Technical Report NMFS; NOAA Fisheries: Silver Spring, MD, USA, 1985; Volume 34, pp. 1–26. [Google Scholar]
- Voigt, M.; Weber, D. Field Guide for Sharks of the Genus Carcharhinus, 1st ed.; Dr. Friedrich Pfeil: Munich, Germany, 2011; pp. 1–151. [Google Scholar]
- Iqbal, M.; Saputra, R.F.; Setiawan, A.; Yustian, I. First photographic inland record of blacktip reef sharks Carcharhinus melanopterus (Carcharhiniformes: Carcharhinidae) in Indonesian waters. Ecol. Montenegrina 2019, 24, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Gausmann, P. Synopsis of global fresh and brackish water occurrences of the bull shark Carcharhinus leucas Valenciennes, 1839 (Pisces: Carcharhinidae), with comments on distribution and habitat use. Integr. Syst. 2021, 4, 55–213. [Google Scholar]
- Cappetta, H. Handbook of Paleoichtyology; Dr. Friedrich Pfeil: Munich, Germany, 2012; Volume 3E, pp. 1–512. [Google Scholar]
- Underwood, C.J.; Ward, D.J. Sharks of the Order Carcharhiniformes from the British Coniacian, Santonian and Campanian (Upper Cretaceous). Palaeontology 2008, 51, 509–536. [Google Scholar] [CrossRef] [Green Version]
- Sorenson, L.; Santini, F.; Alfaro, M.E. The effect of habitat on modern shark diversification. J. Evol. Biol. 2014, 27, 1536–1548. [Google Scholar] [CrossRef] [PubMed]
- Underwood, C.J.; Gunter, G.C. The shark Carcharhinus sp. from the Middle Eocene of Jamaica and the Eocene record of Carcharhinus. Caribb. J. Earth Sci. 2012, 44, 25–30. [Google Scholar]
- Adnet, S.; Antoine, P.O.; Baqri, S.H.; Crochet, J.Y.; Marivaux, L.; Welcomme, J.L.; Métais, G. New tropical carcharhinids (Chondrichthyes, Carcharhiniformes) from the late Eocene–early Oligocene of Balochistan, Pakistan: Paleoenvironmental and paleogeographic implications. J. Asian Earth Sci. 2007, 30, 303–323. [Google Scholar] [CrossRef]
- Adnet, S.; Marivaux, L.; Cappetta, H.; Charruault, A.L.; El Mabrouk, E.; Jiquel, S.; Ammar, H.K.; Marandat, B.; Marzougui, W.; Merzeraud, G.; et al. Diversity and renewal of tropical elasmobranchs around the Middle Eocene Climatic Optimum (MECO) in North Africa: New data from the lagoonal deposits of Djebel el Kébar, Central Tunisia. Palaeontol. Electron. 2020, 23, a38. [Google Scholar] [CrossRef]
- Ralte, V.Z.; Tiwari, R.P.; Malsawma, J. Selachian fishes from Bhuban Formation, Surma Group, Aizawl, Mizoram. J. Geol. Soc. India 2011, 77, 328–348. [Google Scholar] [CrossRef]
- Carrillo-Briceño, J.D.; Maxwell, E.; Aguilera, O.A.; Sànchez, R.; Sànchez-Villagra, M.R. Sawfishes and other elasmobranch assemblages from the Mio-Pliocene of the South Caribbean (Urumaco sequence, northwestern Venezuela). PLoS ONE 2015, 10, e0139230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebersole, J.A.; Cicimurri, D.J.; Stringer, G.L. Taxonomy and biostratigraphy of the elasmobranchs and bony fishes (Chondrichthyes and Osteichthyes) of the lower-to-middle Eocene (Ypresian to Bartonian) Claiborne Group in Alabama, USA, including an analysis of otoliths. Eur. J. Taxon. 2019, 585, 1–274. [Google Scholar] [CrossRef]
- Samonds, K.E.; Andrianavalona, T.H.; Wallett, L.A.; Zalmout, I.S.; Ward, D.J. A middle-late Eocene neoselachian assemblage from nearshore marine deposits, Mahajanga Basin, northwestern Madagascar. PLoS ONE 2019, 14, e0211789. [Google Scholar] [CrossRef] [PubMed]
- Canevet, J.-M.; Lebrun, P. Des dents de requins fossiles! 10. Nouvelles espèces de Carcharhinidae du Miocène de l’Ouest de la France. Foss.–Rev. Fr. Paleontol. 2021, 46, 5–22. [Google Scholar]
- Cicimurri, D.J.; Ebersole, J.A. New Paleogene elasmobranch (Chondrichthyes) records from the Gulf Coastal Plain of the United States, including a new species of Carcharhinus de Blainville, 1816. Cainozoic Res. 2021, 21, 147–164. [Google Scholar]
- Marramà, G.; Carnevale, G.; Kriwet, J. New observations on the anatomy and paleobiology of the Eocene requiem shark †Eogaleus bolcensis (Carcharhiniformes, Carcharhinidae) from Bolca Lagerstätte, Italy. C. R. Palevol 2018, 17, 443–459. [Google Scholar] [CrossRef]
- de Muizon, C.; DeVries, T.J. Geology and paleontology of late Cenozoic marine deposits in the Sacaco area (Peru). Geol. Rundsch. 1985, 74, 547–563. [Google Scholar] [CrossRef]
- de Muizon, C. Les vertebres fossiles de la formation Pisco (Perou): Biostratigraphie, correlations et paleoenvironnement. Geodynamique 1988, 3, 21–24. [Google Scholar]
- Brand, L.R.; Esperante, R.; Chadwick, A.V.; Porras, O.P.; Alomía, M. Fossil whale preservation implies high diatom accumulation rate in the Miocene–Pliocene Pisco Formation of Peru. Geology 2004, 32, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.A.; Ksepka, D.T.; Salas-Gismondi, R.; Altamirano, A.J.; Shawkey, M.D.; D’Alba, L.; Vinther, J.; DeVries, T.J.; Baby, P. Fossil evidence for evolution of the shape and color of penguin feathers. Science 2010, 330, 954–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, J.A.; Ksepka, D.T.; Stucchi, M.; Urbina, M.; Giannini, N.; Bertelli, S.; Narváez, Y.; Boyd, C.A. Paleogene equatorial penguins challenge the proposed relationship between biogeography, diversity, and Cenozoic climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 11545–11550. [Google Scholar] [CrossRef] [Green Version]
- Esperante, R.; Brand, L.; Nick, K.E.; Poma, O.; Urbina, M. Exceptional occurrence of fossil baleen in shallow marine sediments of the Neogene Pisco Formation, Southern Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 257, 344–360. [Google Scholar] [CrossRef]
- Esperante, R.; Brand, L.R.; Chadwick, A.V.; Poma, O. Taphonomy and paleoenvironmental conditions of deposition of fossil whales in the diatomaceous sediments of the Miocene/Pliocene Pisco Formation, southern Peru—A new fossil-lagerstätte. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 417, 337–370. [Google Scholar] [CrossRef]
- Lambert, O.; Bianucci, G.; Post, K.; de Muizon, C.; Salas-Gismondi, R.; Urbina, M.; Reumer, J. The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nature 2010, 466, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Lambert, O.; Bianucci, G.; Urbina, M. Huaridelphis raimondii, a new early Miocene Squalodelphinidae (Cetacea, odontoceti) from the Chilcatay Formation, Peru. J. Vertebr. Paleontol. 2014, 34, 987–1004. [Google Scholar] [CrossRef] [Green Version]
- Lambert, O.; Collareta, A.; Landini, W.; Post, K.; Ramassamy, B.; Di Celma, C.; Urbina, M.; Bianucci, G. No deep diving: Evidence of predation on epipelagic fish for a stem beaked whale from the Late Miocene of Peru. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151530. [Google Scholar] [CrossRef] [PubMed]
- Lambert, O.; de Muizon, C.; Bianucci, G. A new archaic homodont toothed cetacean (Mammalia, Cetacea, Odontoceti) from the early Miocene of Peru. Geodiversitas 2015, 37, 79–108. [Google Scholar] [CrossRef] [Green Version]
- Lambert, O.; Bianucci, G.; de Muizon, C. Macroraptorial sperm whales (Cetacea, Odontoceti, Physeteroidea) from the Miocene of Peru. Zool. J. Linn. Soc. 2017, 179, 404–474. [Google Scholar] [CrossRef]
- Lambert, O.; Martínez-Cáceres, M.; Bianucci, G.; Di Celma, C.; Salas-Gismondi, R.; Steurbaut, E.; Urbina, M.; de Muizon, C. Earliest mysticete from the Late Eocene of Peru sheds new light on the origin of baleen whales. Curr. Biol. 2017, 27, 1535–1541. [Google Scholar]
- Lambert, O.; de Muizon, C.; Malinverno, E.; Di Celma, C.; Urbina, M.; Bianucci, G. A new odontocete (toothed cetacean) from the Early Miocene of Peru expands the morphological disparity of extinct heterodont dolphins. J. Syst. Palaeontol. 2018, 16, 981–1016. [Google Scholar] [CrossRef]
- Lambert, O.; Bianucci, G.; Salas-Gismondi, R.; Di Celma, C.; Steurbaut, E.; Urbina, M.; de Muizon, C. An amphibious whale from the middle Eocene of Peru reveals early South Pacific dispersal of quadrupedal cetaceans. Curr. Biol. 2019, 29, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Lambert, O.; Collareta, A.; Benites-Pallomino, A.; Di Celma, C.; de Muizon, C.; Urbina, M.; Bianucci, G. A new small, mesorostrine inioid (Cetacea, Odontoceti, Delphinida) from four upper Miocene localities in the Pisco Basin, Peru. Pap. Palaeontol. 2021, 7, 1043–1064. [Google Scholar] [CrossRef]
- Lambert, O.; de Muizon, C.; Varas-Malca, R.M.; Urbina, M.; Bianucci, G. Eurhinodelphinids from the early Miocene of Peru: First unambiguous records of these hyper-longirostrine dolphins outside the North Atlantic realm. Riv. Ital. Paleontol. Stratigr. 2021, 127, 17–32. [Google Scholar]
- Parham, J.F.; Pyenson, N.D. New sea turtle from the Miocene of Peru and the iterative evolution of feeding ecomorphologies since the Cretaceous. J. Paleontol. 2010, 84, 231–247. [Google Scholar] [CrossRef]
- Martínez-Cáceres, M.; de Muizon, C. A new basilosaurid (Cetacea, Pelagiceti) from the late Eocene to early Oligocene Otuma Formation of Peru. C. R. Palevol 2011, 10, 517–526. [Google Scholar] [CrossRef]
- Uhen, M.D.; Pyenson, N.D.; DeVries, T.J.; Urbina, M.; Renne, P.R. New middle Eocene whales from the Pisco Basin of Peru. J. Paleontol. 2011, 85, 955–969. [Google Scholar] [CrossRef]
- Collareta, A.; Landini, W.; Lambert, O.; Post, K.; Tinelli, C.; Di Celma, C.; Panetta, D.; Tripodi, M.; Salvadori, P.A.; Caramella, D.; et al. Piscivory in a Miocene Cetotheriidae of Peru: First record of fossilized stomach content for an extinct baleen-bearing whale. Sci. Nat. 2015, 102, 70. [Google Scholar] [CrossRef] [PubMed]
- Collareta, A.; Landini, W.; Chacaltana-Budiel, C.A.; Valdivia-Vera, W.; Altamirano-Sierra, A.; Urbina, M.; Bianucci, G. A well preserved skeleton of the fossil shark Cosmopolitodus hastalis from the late Miocene of Peru, featuring fish remains as fossilized stomach contents. Riv. Ital. Paleontol. Stratigr. 2017, 123, 11–22. [Google Scholar]
- Collareta, A.; Lambert, O.; de Muizon, C.; Benites-Palomino, A.M.; Urbina, M.; Bianucci, G. A new physeteroid from the late Miocene of Peru expands the diversity of extinct dwarf and pygmy sperm whales (Cetacea: Odontoceti: Kogiidae). C. R. Palevol 2020, 19, 79–100. [Google Scholar] [CrossRef]
- Collareta, A.; Di Celma, C.; Bosio, G.; Pierantoni, P.P.; Malinverno, E.; Lambert, O.; Marx, F.G.; Landini, W.; Urbina, M.; Bianucci, G. Distribution and paleoenvironmental framework of middle Miocene marine vertebrates along the western side of the lower Ica Valley (East Pisco Basin, Peru). J. Maps 2021, 17, 7–17. [Google Scholar] [CrossRef]
- Gariboldi, K.; Gioncada, A.; Bosio, G.; Malinverno, G.; Di Celma, C.; Tinelli, C.; Cantalamessa, G.; Landini, W.; Urbina, M.; Bianucci, G. The dolomite nodules enclosing fossil marine vertebrates in the East Pisco Basin, Peru: Field and petrographic insights into the Lagerstätte formation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 438, 81–95. [Google Scholar] [CrossRef]
- Bianucci, G.; Di Celma, C.; Collareta, A.; Landini, W.; Klaas, P.; Tinelli, C.; de Muizon, C.; Bosio, G.; Gariboldi, K.; Gioncada, A.; et al. Fossil marine vertebrates of Cerro Los Quesos: Distribution of cetaceans, seals, crocodiles, seabirds, sharks, and bony fish in a late Miocene locality of the Pisco Basin, Peru. J. Maps 2016, 12, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Bianucci, G.; Di Celma, C.; Landini, W.; Post, K.; Tinelli, C.; de Muizon, C.; Gariboldi, K.; Malinverno, E.; Cantalamessa, G.; Gioncada, A.; et al. Distribution of fossil marine vertebrates in Cerro Colorado, the type locality of the giant raptorial sperm whale Livyatan melvillei (Miocene, Pisco Formation, Peru). J. Maps 2016, 12, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Bianucci, G.; Di Celma, C.; Urbina, M.; Lambert, O. New beaked whales from the late Miocene of Peru and evidence for convergent evolution in stem and crown Ziphiidae (Cetacea, Odontoceti). PeerJ 2016, 4, e2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianucci, G.; Bosio, G.; Malinverno, E.; de Muizon, C.; Villa, I.M.; Urbina, M.; Lambert, O. A new large squalodelphinid (Cetacea, Odontoceti) from Peru sheds light on the Early Miocene platanistoid disparity and ecology. R. Soc. Open Sci. 2018, 5, 172302. [Google Scholar] [CrossRef] [Green Version]
- Bianucci, G.; Collareta, A.; Bosio, G.; Landini, W.; Gariboldi, K.; Gioncada, A.; Lambert, O.; Malinverno, E.; de Muizon, C.; Varas-Malca, R.; et al. Taphonomy and palaeoecology of the lower Miocene marine vertebrate assemblage of Ullujaya (Chilcatay Formation, East Pisco Basin, southern Peru). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 511, 256–279. [Google Scholar] [CrossRef]
- Bianucci, G.; Marx, F.G.; Collareta, A.; Di Stefano, A.; Landini, W.; Morigi, C.; Varola, A. Rise of the titans: Baleen whales became giants earlier than thought. Biol. Lett. 2019, 15, 20190175. [Google Scholar] [CrossRef] [Green Version]
- Bianucci, G.; de Muizon, C.; Urbina, M.; Lambert, O. Extensive diversity and disparity of the Early Miocene platanistoids (Cetacea, Odontoceti) in the southeastern Pacific (Chilcatay Formation, Peru). Life 2020, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Gioncada, A.; Collareta, A.; Gariboldi, K.; Lambert, O.; Di Celma, C.; Bonaccorsi, E.; Urbina, M.; Bianucci, G. Inside baleen: Exceptional microstructure preservation in a late Miocene whale skeleton from Peru. Geology 2016, 44, 839–842. [Google Scholar] [CrossRef]
- Gioncada, A.; Gariboldi, K.; Collareta, A.; Di Celma, C.; Bosio, G.; Malinverno, E.; Lambert, O.; Pike, J.; Urbina, M.; Bianucci, G. Looking for the key to preservation of fossil marine vertebrates in the Pisco Formation of Peru: New insights from a small dolphin skeleton. Andean Geol. 2018, 45, 379–398. [Google Scholar] [CrossRef]
- Gioncada, A.; Petrini, R.; Bosio, G.; Gariboldi, K.; Collareta, A.; Malinverno, E.; Bonaccorsi, E.; Di Celma, C.; Pasero, M.; Urbina, M.; et al. Insights into the diagenetic environment of fossil marine vertebrates of the Pisco Formation (late Miocene, Peru) from mineralogical and Sr-isotope data. J. South Am. Earth Sci. 2018, 81, 141–152. [Google Scholar] [CrossRef]
- Di Celma, C.; Malinverno, E.; Bosio, G.; Collareta, A.; Gariboldi, K.; Gioncada, A.; Molli, G.; Landini, W.; Sarti, G.; Bianucci, G. Sequence stratigraphy and paleontology of the Upper Miocene Pisco Formation along the western side of the lower Ica Valley (Ica Desert, Peru). Riv. Ital. Paleontol. Stratigr. 2017, 123, 255–273. [Google Scholar]
- Di Celma, C.; Malinverno, E.; Collareta, A.; Bosio, G.; Gariboldi, K.; Lambert, O.; Landini, W.; Pierantoni, P.P.; Gioncada, A.; Villa, I.M.; et al. Facies analysis, stratigraphy and marine vertebrate assemblage of the lower Miocene Chilcatay Formation at Ullujaya (Pisco basin, Peru). J. Maps 2018, 14, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Di Celma, C.; Pierantoni, P.P.; Malinverno, E.; Collareta, A.; Lambert, O.; Landini, W.; Bosio, G.; Gariboldi, K.; Gioncada, A.; de Muizon, C.; et al. Allostratigraphy and paleontology of the lower Miocene Chilcatay Formation in the Zamaca area, East Pisco basin, southern Peru. J. Maps 2019, 15, 393–405. [Google Scholar] [CrossRef] [Green Version]
- Marx, F.G.; Collareta, A.; Gioncada, A.; Post, K.; Lambert, O.; Bonaccorsi, E.; Urbina, M.; Bianucci, G. How whales used to filter: Exceptionally preserved baleen in a Miocene cetotheriid. J. Anat. 2017, 231, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, F.G.; Lambert, O.; de Muizon, C. A new Miocene baleen whale from Peru deciphers the dawn of cetotheriids. R. Soc. Open Sci. 2017, 4, 170560. [Google Scholar] [CrossRef] [Green Version]
- Ramassamy, B.; Lambert, O.; Collareta, A.; Urbina, M.; Bianucci, G. Description of the skeleton of the fossil beaked whale Messapicetus gregarius: Searching potential proxies for deep-diving abilities. Foss. Rec. 2018, 21, 11–32. [Google Scholar] [CrossRef] [Green Version]
- Bosio, G.; Gioncada, A.; Gariboldi, K.; Bonaccorsi, E.; Collareta, A.; Pasero, M.; Di Celma, C.; Malinverno, E.; Urbina, M.; Bianucci, G. Mineralogical and geochemical characterization of fossil bones from a Miocene marine Konservat-Lagerstätte. J. South Am. Earth Sci. 2021, 105, 102924. [Google Scholar] [CrossRef]
- Bosio, G.; Collareta, A.; Di Celma, C.; Lambert, O.; Marx, F.G.; de Muizon, C.; Gioncada, A.; Gariboldi, K.; Malinverno, E.; Varas-Malca, R.; et al. Taphonomy of marine vertebrates of the Pisco Formation (Miocene, Peru): Insights into the origin of an outstanding Fossil-Lagerstätte. PLoS ONE 2021, 16, e0254395. [Google Scholar] [CrossRef]
- Boskovic, D.S.; Vidal, U.L.; Nick, K.E.; Esperante, R.; Brand, L.R.; Wright, K.R.; Sandberg, L.B.; Siviero, B.C.T. Structural and protein preservation in fossil whale bones from the Pisco Formation (Middle-Upper Miocene), Peru. Palaios 2021, 36, 155–164. [Google Scholar] [CrossRef]
- Benites-Palomino, A.; Vélez-Juarbe, J.; Altamirano-Sierra, A.; Collareta, A.; Carrillo-Briceño, J.D.; Urbina, M. Sperm whales (Physeteroidea) from the Pisco Formation, Peru, and their trophic role as fat sources for Late Miocene sharks. Proc. R. Soc. B Biol. Sci. 2022, 289, 20220774. [Google Scholar] [CrossRef] [PubMed]
- Bianucci, G.; Collareta, A. An overview of the fossil record of cetaceans from the East Pisco Basin (Peru). Boll. Soc. Paleontol. Ital. 2022, 61, 19–60. [Google Scholar]
- Collareta, A.; Tejada-Medina, L.M.; Chacaltana-Budiel, C.A.; Landini, W.; Altamirano-Sierra, A.; Urbina, M.; Bianucci, G. A rhinopristiform sawfish (genus Pristis) from the middle Eocene (Lutetian) of southern Peru and its regional implications. Carnets Geol. 2020, 20, 91–105. [Google Scholar] [CrossRef]
- Shimada, K.; Chandler, R.E.; Lam, O.L.T.; Tanaka, T.; Ward, D.J. A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade. Hist. Biol. 2017, 29, 704–714. [Google Scholar] [CrossRef]
- Landini, W.; Altamirano-Sierra, A.; Collareta, A.; Di Celma, C.; Urbina, M.; Bianucci, G. The late Miocene elasmobranch assemblage from Cerro Colorado (Pisco Formation, Peru). J. S. Am. Earth Sci. 2017, 73, 168–190. [Google Scholar] [CrossRef]
- Landini, W.; Collareta, A.; Di Celma, C.; Malinverno, E.; Urbina, M.; Bianucci, G. The early Miocene elasmobranch assemblage from Zamaca (Chilcatay Formation, Peru). J. S. Am. Earth Sci. 2019, 91, 352–371. [Google Scholar] [CrossRef]
- Landini, W.; Colareta, A.; Pesci, F.; Di Celma, C.; Urbina, M.; Bianucci, G. A secondary nursery area for the copper shark Carcharhinus brachyurus from the late Miocene of Peru. J. S. Am. Earth Sci. 2017, 78, 164–174. [Google Scholar] [CrossRef]
- Sibert, E.C.; Rubin, L.D. An early Miocene extinction in pelagic sharks. Science 2021, 372, 1105–1107. [Google Scholar] [CrossRef]
- Feichtinger, I.; Adnet, S.; Cuny, G.; Guinot, G.; Kriwet, J.; Neubauer, T.A.; Pollerspöck, J.; Shimada, K.; Straube, N.; Underwood, C.; et al. Comment on “An early Miocene extinction in pelagic sharks”. Science 2021, 374, eabk0632. [Google Scholar] [CrossRef]
- Naylor, G.J.; de Lima, A.; Castro, J.I.; Hubbell, G.; de Pinna, M.C. Comment on “An early Miocene extinction in pelagic sharks”. Science 2021, 374, eabj8723. [Google Scholar] [CrossRef]
- Bengtson, P. Open nomenclature. Palaeontology 1988, 31, 223–227. [Google Scholar]
- Marocco, R.; de Muizon, C. Los vertebrados del Neogeno de la costa sur del Perú: Ambiente sedimentario y condiciones de fosilización. Bull. Inst. Français Ét. Andin. 1988, 17, 105–117. [Google Scholar]
- Dunbar, R.B.; Marty, R.C.; Baker, P.A. Cenozoic marine sedimentation in the Sechura and Pisco basins, Peru. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 77, 235–261. [Google Scholar] [CrossRef]
- DeVries, T.J.; Jud, N.A. Lithofacies patterns and paleogeography of the Miocene Chilcatay and lower Pisco depositional sequences (East Pisco Basin, Peru). Bol. Soc. Geol. Perú 2018, 8, 124–167. [Google Scholar]
- DeVries, T.J.; Barron, J.A.; Urbina, M.; Ochoa, D.; Esperante, R.; Snee, L.W. The Miocene stratigraphy of the Laberinto area (Río Ica Valley) and its bearing on the geological history of the East Pisco Basin (south-central Peru). J. S. Am. Earth Sci. 2021, 111, 103458. [Google Scholar] [CrossRef]
- Bosio, G.; Bianucci, G.; Collareta, A.; Landini, W.; Urbina, M.; Di Celma, C. Ultrastructure, composition, and 87Sr/86Sr dating of shark teeth from lower Miocene sediments of southwestern Peru. J. S. Am. Earth Sci. 2022, 118, 103909. [Google Scholar] [CrossRef]
- Travis, R.B.; Gonzales, G.; Pardo, A. Hydrocarbon potential of coastal basins of Peru: Hydrocarbons. In Circum-Pacific Energy and Mineral Resources, 1st ed.; American Association of Petroleum Geologists: Tulsa, OK, USA, 1976; pp. 331–338. [Google Scholar]
- Thornburg, T.M.; Kulm, L.D. Sedimentary basins of the Peru continental margin: Structure, stratigraphy, and Cenozoic tectonics from 6 S to 16 S latitude. Nazca Plate Crustal Form. Andean Converg. 1981, 154, 393–422. [Google Scholar]
- DeVries, T.J.; Schrader, H. Middle Miocene marine sediments in the Pisco basin (Peru). Bol. Soc. Geol. Perú 1997, 87, 1–13. [Google Scholar]
- Di Celma, C.; Malinverno, E.; Bosio, G.; Gariboldi, K.; Collareta, A.; Gioncada, A.; Landini, W.; Pierantoni, P.P.; Bianucci, G. Intraformational unconformities as a record of late Miocene eustatic falls of sea level in the Pisco Formation (southern Peru). J. Maps 2018, 14, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Di Celma, C.; Pierantoni, P.P.; Volatili, T.; Molli, G.; Mazzoli, S.; Sarti, G.; Ciattoni, S.; Bosio, G.; Malinverno, E.; Collareta, A.; et al. Towards deciphering the Cenozoic evolution of the East Pisco Basin (southern Peru). J. Maps 2022, 1–16. [Google Scholar] [CrossRef]
- Lambert, O.; de Muizon, C.; Urbina, M.; Bianucci, G. A new longirostrine sperm whale (Cetacea, Physeteroidea) from the lower Miocene of the Pisco Basin (southern coast of Peru). J. Syst. Palaeontol. 2020, 18, 1707–1742. [Google Scholar] [CrossRef]
- Bianucci, G.; Urbina, M.; Lambert, O. A new record of Notocetus vanbenedeni (Squalodelphinidae, Odontoceti, Cetacea) from the Early Miocene of Peru. Comptes. Rendus. Palevol. 2015, 14, 5–13. [Google Scholar] [CrossRef]
- Acosta-Hospitalache, C.; Stucchi, M. Nuevos restos terciarios de Spheniscidae (Aves, Sphenisciformes) procedentes de la costa del Perú. Span. J. Palentol. 2005, 20, 1–5. [Google Scholar] [CrossRef]
- Wood, R.C.; Johnson-Gove, J.; Gaffney, E.S.; Maley, K.F. Evolution and phylogeny of the leatherback turtles (Dermochelyidae), with descriptions of new fossil taxa. Chelonian Conserv. Biol. 1996, 2, 266–286. [Google Scholar]
- Alván de la Cruz, A. Geología de Ocucaje: Aportes en la sedimentología y paleontología de Lomas de Ullujaya (Ica, Perú). Rev. Investig. Fac. Ing. Geol. Min. Met. Geogr. 2008, 11, 51–59. [Google Scholar]
- Renz, M. Desert Sharks; PaleoPress: Lehigh Acres, FL, USA, 2009; pp. 1–197. [Google Scholar]
- Collareta, A.; Lambert, O.; Marx, F.G.; de Muizon, C.; Varas-Malca, R.; Landini, W.; Bosio, G.; Malinverno, E.; Gariboldi, K.; Gioncada, A.; et al. Vertebrate Palaeoecology of the Pisco Formation (Miocene, Peru): Glimpses into the ancient Humboldt Current Ecosystem. J. Mar. Sci. Eng. 2021, 9, 1188. [Google Scholar] [CrossRef]
- DeVries, T.J.; Frassinetti, D. Range extensions and biogeographic implications of Chilean Neogene mollusks found in Peru. Mus. Nac. Hist. Nat. Chile 2003, 52, 119–135. [Google Scholar]
- Coletti, G.; Bosio, G.; Collareta, A.; Buckeridge, J.S.; Consani, S.; El Kateb, A. Palaeoenvironmental analysis of the Miocene barnacle facies: Case studies from Europe and South America. Geol. Carpathica 2018, 69, 573–592. [Google Scholar] [CrossRef] [Green Version]
- Coletti, G.; Collareta, A.; Bosio, G.; Urbina-Schmitt, M.; Buckeridge, J.S. Perumegabalanus calziai gen. et sp. nov., a new intertidal megabalanine barnacle from the early Miocene of Peru. Neues Jahrb. Geol. Paläontol. Abh. 2019, 294, 197–212. [Google Scholar] [CrossRef]
- Collareta, A.; Coletti, G.; Bosio, G.; Buckeridge, J.S.; de Muizon, C.; DeVries, T.J.; Varas-Malca, R.M.; Altamirano-Sierra, A.; Urbina, M.; Bianucci, G. A new barnacle (Cirripedia: Neobalanoformes) from the early Miocene of Peru: Palaeoecological and palaeobiogeographical implications. Neues Jahrb. Geol. Paläontol. Abh. 2019, 292, 321–338. [Google Scholar] [CrossRef]
- Kočí, T.; Bosio, G.; Collareta, A.; Sanfilippo, R.; Ekrt, B.; Urbina, M.; Malinverno, E. First report on the cirratulid (Annelida, Polychaeta) reefs from the Miocene Chilcatay and Pisco Formations (East Pisco Basin, Peru). J. S. Am. Earth Sci. 2021, 107, 103042. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Kočí, T.; Bosio, G.; Collareta, A.; Ekrt, B.; Malinverno, E.; Di Celma, C.; Urbina, M.; Bianucci, G. An investigation of vermetid reefs from the Miocene of Peru, with the description of a new species. J. S. Am. Earth Sci. 2021, 108, 103233. [Google Scholar] [CrossRef]
- Bosio, G.; Malinverno, E.; Collareta, A.; Di Celma, C.; Gioncada, A.; Parente, M.; Berra, F.; Marx, F.G.; Vertino, A.; Urbina, M.; et al. Strontium Isotope Stratigraphy and the thermophilic fossil fauna from the middle Miocene of the East Pisco Basin (Peru). J. S. Am. Earth Sci. 2020, 97, 102399. [Google Scholar] [CrossRef]
- Bosio, G.; Malinverno, E.; Villa, I.M.; Di Celma, C.; Gariboldi, K.; Gioncada, A.; Barberini, V.; Urbina, M.; Bianucci, G. Tephrochronology and chronostratigraphy of the Miocene Chilcatay and Pisco formations (East Pisco basin, Peru). Newsl. Stratigr. 2020, 53, 213–247. [Google Scholar] [CrossRef]
- Compagno, L.J.V. Interrelationships of living elasmobranchs. In Interrelationships of Fishes; Greenwood, P.H., Miles, R.S., Patterson, C., Eds.; Academic Press: London, UK, 1973; pp. 15–61. [Google Scholar]
- Jordan, D.S.; Evermann, B.W. A Check-List of the Fishes and Fish-Like Vertebrates of North and Middle America; Report of the Commissioner; Government Printing Office: Washington, DC, USA, 1896; pp. 207–584. [Google Scholar]
- De Blainville, H.M.D. Prodrome d’une nouvelle distribution systématique du règne animal. Bull. Soc. Philomath. 1816, 8, 105–124. [Google Scholar]
- Quoy, J.R.C.; Gaimard, J.P. Description des poissons. In Voyage Autour du Monde, Entrepris par Ordre du Roi, Execute Sur les les Corvettes de L.M. “L’Uranie” et “La Physicienne”, Pendant les Années 1817, 1818, 1819 et 1820; Chapter IX; de Freycinet, M.L., Ed.; Pillet Aîne: Paris, France, 1824; pp. 192–401. [Google Scholar]
- Carrillo-Briceño, J.D.; Aguilera, O.A.; De Gracia, C.; Aguirre-Fernàndez, G.; Kindlimann, R.; Sànchez-Villagra, M.R. An early Neogene elasmobranch fauna from the southern Caribbean (western Venezuela). Palaeontol. Electron. 2016, 19.2.28A. [Google Scholar] [CrossRef]
- Richter, M.; Ward, D.J. Fish remains from the Santa Marta Formation (Late Cretaceous) of James Ross Island, Antarctica. Antarct. Sci. 1990, 2, 67–76. [Google Scholar] [CrossRef]
- Mannering, A.A.; Hiller, N. An early Cenozoic neoselachian shark fauna from the Southwest Pacific. Palaeontology 2008, 51, 1341–1365. [Google Scholar] [CrossRef]
- Carrillo-Briceño, J.D.; Aguilera, O.A.; Rodriguez, F. Fossil Chondrichthyes from the central eastern Pacific Ocean and their paleoceanographic significance. J. S. Am. Earth Sci. 2014, 51, 76–90. [Google Scholar] [CrossRef]
- Naylor, G.J.; Marcus, L.F. Identifying isolated shark teeth of the genus Carcharhinus to species: Relevance for tracking phyletic change through the fossil record. Am. Mus. Novit. 1994, 3109, 1–53. [Google Scholar]
- Bourdon, J. The Life and Times of Long Dead Sharks. 1999. Available online: www.elasmo.com (accessed on 27 August 2022).
- Whitenack, L.B.; Motta, P.J. Performance of shark teeth during puncture and draw: Implications for the mechanics of cutting. Biol. J. Linn. Soc. 2010, 100, 271–286. [Google Scholar] [CrossRef]
- Mollen, F.H. A new species of Abdounia (Elasmobranchii, Carcharhinidae) from the base of the Boom Clay Formation (Oligocene) in northwest Belgium. Geol. Belg. 2007, 10, 69–79. [Google Scholar]
- Perez, V.J. The chondrichthyan fossil record of the Florida Platform (Eocene–Pleistocene). Paleobiology, 2022; ahead-of-print. [Google Scholar] [CrossRef]
- Perez, V.J.; Pimiento, C.; Hendy, A.; González-Barba, G.; Hubbell, G.; MacFadden, B.J. Late Miocene chondrichthyans from Lago Bayano, Panama: Functional diversity, environment and biogeography. J. Paleontol. 2017, 91, 512–547. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, D.K.; Haney, T.A.; Louie, K.D. Genes, diversity, and geological process on the Pacific coast. Annu. Rev. Earth Planet. Sci. 2004, 32, 601–652. [Google Scholar] [CrossRef] [Green Version]
- Coates, A.G.; Stallard, R.F. How old is the Isthmus of Panama? Bull. Mar. Sci. 2013, 89, 801–813. [Google Scholar] [CrossRef]
- Bianucci, G.; Collareta, A.; Post, K.; Varola, A.; Lambert, O. A new record of Messapicetus from the Pietra Leccese (Late Miocene, Southern Italy): Antitropical distribution in a fossil beaked whale (Cetacea, Ziphiidae). Riv. Ital. Paleontol. Stratigr. 2016, 122, 63–74. [Google Scholar]
- Bianucci, G.; Llàcer, S.; Cardona, J.Q.; Collareta, A.; Florit, A.R. A new beaked whale record from the upper Miocene of Menorca, Balearic Islands, based on CT-scan analysis of limestone slabs. Acta Palaeontol. Pol. 2019, 64, 63–74. [Google Scholar] [CrossRef]
- O’Dea, E.; Furner, R.; Wakelin, S.; Siddorn, J.; While, J.; Sykes, P.; King, R.; Holt, J.; Hewitt, H. The CO5 configuration of the 7 km Atlantic Margin Model: Large-scale biases and sensitivity to forcing, physics options and vertical resolution. Geosci. Model Dev. 2017, 10, 2947–2969. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo, C.; Montes, C.; Cardona, A.; Silvestro, D.; Antonelli, A.; Bacon, C.D. Comment (1) on “Formation of the Isthmus of Panama” by O’Dea et al. Sci. Adv. 2017, 3, e1602321. [Google Scholar] [CrossRef] [Green Version]
- Peri, E.; Collareta, A.; Insacco, G.; Bianucci, G. An Inticetus-like (Cetacea: Odontoceti) postcanine tooth from the Pietra leccese (Miocene, southeastern Italy) and its palaeobiogeographical implications. Neues Jahrb. Geol. Paläontologie Abh. 2019, 291, 221–228. [Google Scholar] [CrossRef]
- Landini, W.; Collareta, A.; Bianucci, G. The origin of biogeographic segregation in the copper shark (Carcharhinus brachyurus): An integrative reconstruction based on neontological and paleontological data. Vie Milieu 2020, 70, 117–132. [Google Scholar]
- Collareta, A.; Landini, W.; Bianucci, G.; Di Celma, C. Until Panama do us part: New finds from the Pliocene of Ecuador provide insights into the origin and palaeobiogeographic history of the extant requiem sharks Carcharhinus acronotus and Nasolamia velox. Neues Jahrb. Geol. Paläontol. Abh. 2021, 300, 103–115. [Google Scholar] [CrossRef]
- Purdy, R.W.; Schneider, V.P.; Applegate, S.P.; McLellan, J.H.; Meyer, R.L.; Slaughter, R.T. The Neogene sharks, rays, and bony fishes from Lee Creek Mine, Aurora, North Carolina. Smithson. Contrib. Paleobiol. 2001, 90, 71–202. [Google Scholar]
- Kent, B.W. The Cartilaginous fishes (chimaeras, sharks, and rays) of Calvert Cliffs, Maryland, USA. Smithson. Contrib. Paleobiol. 2018, 100, 45–160. [Google Scholar]
- Laurito Mora, C.A. Los Seláceos Fósiles de la Localidad de Alto Guayacán (y Otros Ictiolitos Asociados): Mioceno Superior-Plioceno Inferior de la Formación Uscari; ScienceOpen: San José, CA, USA, 1999; p. 567. [Google Scholar]
- Maisey, J.G. What is an ‘elasmobranch’? The impact of palaeontology in understanding elasmobranch phylogeny and evolution. J. Fish Biol. 2012, 80, 918–951. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collareta, A.; Kindlimann, R.; Baglioni, A.; Landini, W.; Sarti, G.; Altamirano, A.; Urbina, M.; Bianucci, G. Dental Morphology, Palaeoecology and Palaeobiogeographic Significance of a New Species of Requiem Shark (Genus Carcharhinus) from the Lower Miocene of Peru (East Pisco Basin, Chilcatay Formation). J. Mar. Sci. Eng. 2022, 10, 1466. https://doi.org/10.3390/jmse10101466
Collareta A, Kindlimann R, Baglioni A, Landini W, Sarti G, Altamirano A, Urbina M, Bianucci G. Dental Morphology, Palaeoecology and Palaeobiogeographic Significance of a New Species of Requiem Shark (Genus Carcharhinus) from the Lower Miocene of Peru (East Pisco Basin, Chilcatay Formation). Journal of Marine Science and Engineering. 2022; 10(10):1466. https://doi.org/10.3390/jmse10101466
Chicago/Turabian StyleCollareta, Alberto, René Kindlimann, Alessio Baglioni, Walter Landini, Giovanni Sarti, Alí Altamirano, Mario Urbina, and Giovanni Bianucci. 2022. "Dental Morphology, Palaeoecology and Palaeobiogeographic Significance of a New Species of Requiem Shark (Genus Carcharhinus) from the Lower Miocene of Peru (East Pisco Basin, Chilcatay Formation)" Journal of Marine Science and Engineering 10, no. 10: 1466. https://doi.org/10.3390/jmse10101466