1. Introduction
Moesziomyces antarcticus (former
Candida antarctica and
Pseudozyma antarctica) is a yeast strain that is well known as a mannosylerythritol lipid (MELs) producer. MEL is an amphiphilic molecule and a potent biosurfactant, with a wide array of possible industrial applications, ranging from cleaning and bioremediation [
1], to cosmetic and therapeutic uses [
2]. Lowering the cultivation and downstream costs is still required, to enable large-scale production of MEL.
Moesziomyces antarcticus is also a potent producer of extracellular enzymes, mainly lipases. The main lipase produced is
Candida antarctica lipase B (CAL-B), which has a crucial industrial role in the hydrolysis of lipidic materials [
3]. The lipase production by this yeast is crucial for the consolidated production of MEL from lipid-based substrates.
Biotechnological production involves a high consumption of fresh water. Sea water is, however, an abundant resource, which could be utilized in biotechnological production (it is already used for algae cultivation), if the 3.5% of salt present in the medium, as well as marine microbiological contaminants, do not affect the working microorganism’s performance. One of the main issues with this concept is that sea water negatively effects stainless steel bioreactors, by corroding the surfaces in contact with water of over time [
4]. Alternative materials for bioreactor production should be considered, when designing the equipment in a production line of this kind.
The aim of this study was to understand if M. antarcticus can produce lipases and MEL in media with NaCl present in the growth medium, as this could contribute to the use of sea water in the production process, thus increasing the overall sustainability of the process. NaCl, sea water, or wastewater with high salinity (such as that generated in the process of producing drinking water from seawater by reverse osmosis and some food processing facilities) could be also used as a basis for the growth medium. This would give a new perspective for the MELs production process, to compete with other industrially produced biosurfactants. The overall goal was to stimulate beneficial physical changes in the microorganism, which have been reported for other yeasts, to simplify downstream processing, decrease the need for fresh water during production, as well as to enable growth and MEL production in non-sterile conditions. To fully study the interactions of M. antarcticus and NaCl, and to study the physiological effects it has on the microorganism, both short exposures to osmotic stress (addition of pure NaCl) and constant stress (with NaCl present in the media from the very beginning of the fermentation) were examined.
Furthermore, this would possibly enable non-sterile fermentation conditions to be used, as the osmotic stress could deter most common contaminants in the fermentation step, e.g., from the Escherichia and Bacillus genera, which lack the ability to survive in hypertonic conditions [
5].
The effect of sodium chloride in various concentrations on yeasts has been studied and documented in detail in the literature. Decades of research on this topic have mostly focused on baker’s yeast (Saccharomyces cerevisiae), as well as infectious species, such as those of Candida, Cryptococcus, etc. For these genera, the literature sources report that NaCl causes physical changes as a result of the yeast’s reaction to the stressor. NaCl is commonly used to induce an osmotic stress response, activating a mitogen-activated protein (MAP) kinase cascade, the high-osmolarity glycerol (HOG) pathway, as an attempt by the cell to equalize the transmembrane osmotic pressure [
6]. This effect is fast and fully reversible, meaning that the cell can facilitate the release and metabolization of unneeded glycerol in the cell after adaptation or further changes in the medium.
The continuous extensive use of petroleum products results in their accidental release in marine and terrestrial ecosystems during their use and transportation [
7]. Their presence causes extensive environmental harm, and the search for alternatives to invasive remediation efforts has turned towards more eco-friendly alternatives. Bioremediation of oil spills is performed by introducing microorganisms that are able to degrade the spilled hydrocarbons, as well as chemicals of biological origin which facilitate their natural degradation [
8,
9].
Moesziomyces antarcticus is known to metabolize hydrocarbons [
10], and MELs were suggested as a potential bioremediation chemical that accelerates petroleum degradation by marine microorganisms [
11]. With this in mind, the ability of
M. antarcticus to metabolize hydrocarbons and crude oil into MELs was evaluated.
Before being applied in the field of bioremediation, the potential toxic effect of MELs on living organisms must be evaluated. In this work, MELs ecotoxicity was evaluated for the first time, by testing its inhibitory effects on a marine organism used as a marine ecotoxicity model, Artemia franciscana.
2. Materials and Methods
2.1. Microorganisms and Maintenance
Moesziomyces antarcticus PYCC 5048T (CBS 5955) was provided by the Portuguese Yeast Culture Collection (PYCC, Caparica, Portugal), CREM, FCT/UNL, Caparica, Portugal, and maintained following a previously established protocol [
12].
Escherichia coli DH5-α was used to induce contamination when testing the ability of M. antarcticus to grow in non-sterile conditions. A tube with 5 mL of Luria–Bertani broth was inoculated with 10 μL of cryopreserved stock (with 17.5% glycerol), containing approx. 5 × 108 cells/mL. The tubes were incubated for 24 h, at 37 °C and 250 rpm. A sample amount containing approx. 1 × 105 of cells was used to simulate contamination. This strain was acquired from the iBB laboratory strain collection.
2.2. Media and Cultivation Conditions
Erlenmeyer flasks were used for the preparation of inoculum and batch fermentation. The previously described media and conditions were used [
12], with the initial addition of 40 g·L
−1 D-glucose, and 20 g·L
−1 soybean oil addition on day 4. Inoculum flasks were incubated for 48 h, after which they were used as a 10% (
v/v) inoculum for batch cultivation flasks, which were afterwards incubated in an orbital shaker.
In certain conditions, NaCl was added to the media as a stress agent, in amounts of 1%, 3.5%, 5%, 7.5%, and 10% (w/w). Periodical samples were collected to quantify biomass (CDW), monosaccharides, lipids, and MELs, as well as protein content and lipase activity.
For tests with hydrocarbons, light crude oil obtained from Troll field, Norway, was used, as well as a hydrocarbon mixture of n-dodecane, n-tetradecane, and n-hexadecane in equal mass ratios. Hydrocarbon substrates were added at a 5 g·L−1 concentration on day 4, into the culture grown on the previously described media, with an initial addition of 40 g·L−1 of glucose. These tests were performed with fresh water (0% NaCl) and marine-level saline water (3.5%).
2.3. Biomass Content and Viable Cell Determination
Cell growth was quantified by cell dry weight (CDW), using a previously described protocol [
12]. Supernatant from the centrifuged samples was collected and used for various analyses.
To determine the viable cell content, samples were diluted and plated, and the colonies formed were counted.
2.4. Exposure to NaCl as a Stress Agent, and Release of Intracellular Glycerol
When indicated, cells were cultivated with no NaCl present, which was added later, in order to cause osmotic stress in the established cultures of
M. antarcticus. For these experiments, 5 mL samples of fermentation broth were placed in test tubes with pure NaCl (ITW Reagents, Glenview, IL, USA, 99.5% purity), using concentrations (
w/
v) of 1%, 3.5%, 5%, and 7.5%, and slowly mixed on a rotating mixer at 12 rpm, to prevent physiological damage to the culture, for 30 and 120 min. In order to release intracellular glycerol, a method reported in literature was applied [
13]. Namely, after the required time of exposure to NaCl, tubes were placed in boiling water for 10 min and centrifuged for 3 min at 4000 rpm, to remove solid cell debris, and the supernatant was used for analysis.
2.5. Analysis of Fermentable Sugars, Nitrates, and Glycerol Concentrations
In the collected sample supernatants, monosaccharides, nitrates, and glycerol were quantified in a high-performance liquid chromatography (HPLC) system (VWR Hitachi, Darmstadt, Germany) equipped with a RI detector (L-2490, VWR Hitachi, Darmstadt, Germany), UV-detector (L-2420, VWR Hitachi, Darmstadt, Germany), and a RezexTM RHM-Monosaccharide H+ (8%) column (300 mm × 7.8 mm, Phenomenex, Torrance, CA, USA), at 65 °C. Milli-Q water was used as mobile phase at 0.5 mL·min−1.
2.6. Analysis of MEL and Lipid Concentrations
MEL concentrations in the samples were calculated based on the results acquired through gas chromatography of methyl-esters, as previously described [
12]. MELs were quantified using the amount of C8, C10, and C12 fatty acids, considering a molecular weight between 574 and 676 g·mol
−1, depending on the length of the two-acyl chain (C8–C12). The quantification of glycolipids based on a specific moiety was previously described by Faria et al. [
14].
All lipids and lipid derivatives (free fatty acids, mono-, di-, and triglycerides) are represented as lipids in graphs.
In some cases, the solid MELs-rich precipitates (beads) were analyzed separately. When present, the beads were removed from the broth using a metal sieve (opening diameter ~1 mm) and dried at 60 °C for 48 h, to determine their moisture content. Then, the dry bead mass was extracted with ethyl acetate twice (10:1 w/w of solvent to bead was used). The liquid phase of the extract was used for GC analysis after solvent evaporation, while the solid fraction was dried and measured, to determine content of biomass (which included cell debris and some medium fractions).
2.7. Analysis of Crude Oil and Hydrocarbon Concentrations
To extract the hydrocarbons from the fermentation broth, n-hexane (≥95%, HPLC grade, Fisher Chemical, Hampton, NH, USA) was used. After 12 days of fermentation, 0.1 mL of pristane (10 g·L−1 in n-hexane) was added as an internal standard, to quantify the extraction losses. Then, the total content of the flask was transferred to a separation vessel, and a total 50 mL of hexane was added. Another 25 mL of hexane was used to rinse each flask and then added to the vessel. The contents were mixed vigorously for 1 min, left to separate, and the top organic layer was removed. This hexane phase was evaporated on a rotavapor, corrected to 50 mL, and a 1 mL sample was used for analysis.
Samples were analyzed using gas chromatography (Hewlett-Packard, HP5890, Palo Alto, CA, USA). The initial oven temperature was 60 °C and held for 2 min. The temperature was then increased at a rate of 6 °C/min, until reaching 310 °C, where it was kept for 5 min. Injector and detector temperature was 300 °C and 310 °C, respectively. Purge was set to turn on after 0.66 min. Injected sample volume was 1 μL. Calibration curves were made for both crude oil and alkane mixture using a series of dilutions (5 g·L
−1, 2 g·L
−1, 1 g·L
−1, 0.5 g·L
−1, 0.2 g·L
−1 in n-hexane), with 5-α-androstane as a secondary external standard. The relative response factor (RRF) was then calculated using Equation (1).
Equation (1) RRF equation for GC analysis of hydrocarbons.
For the crude oil samples, automatic integration of peaks was used with a minimum area to height ratio of 1, from 6 min to 50 min of the GC-FID chromatogram, from which the area of the internal standard peaks (5-α-androstane, pristane) was subtracted. For the alkane mixture, the three hydrocarbon areas were individually integrated. Based on the initial and measured concentrations of the standards, final values were corrected to account for losses in extraction and measurement errors.
2.8. Extracellular Protein and Lipase Activity Analysis
Extracellular protein content was assessed using a Thermo Scientific™ (Waltham, MA, USA), Pierce™ (Outagamie County, WI, USA), BCA Protein Assay, in a 96 well microplate, with albumin as standard. All runs were performed in triplicate.
The enzymatic assays were performed based on a method acquired from the literature [
15], in a 96 well microplate, with p-nitrophenyl butyrate used as substrate. One unit (U) of lipase activity was defined as the amount of enzyme releasing 1 μmol p-nitrophenol per minute.
2.9. Ecotoxicity Experimental Setup and Protocol
For testing the marine organism ecotoxic effects of MELs, an Artemia Toxicity Screening Test for Estuarine and Marine Waters ARTOXKIT M kit was used, following the standard protocol (Artoxkit, 1990). In a 24 multiwell plate, 10 spores were added in each plate, and incubated at 25 °C for 5 days, with exposure to light. After this period, living shrimp were counted, and the IC50 was determined. Tests were performed in triplicate, with marine-level saline water (3.5% of NaCl).
To compare MELs to other biosurfactants, sophorolipids (provided by Holiferm, Manchester, UK) and rhamnolipids (90% purity, Agae Technologies, Corvallis, OR, USA) were used; as well as Corexit (Nalco Holding Company, Naperville, IL, USA), a chemical oil dispersant used for bioremediation, and Tween 80, a commercial chemical surfactant.
3. Results
Moesziomyces antarcticus culture was exposed to various levels of NaCl, in different groups of experiments, to observe and describe the various effects of saline stress on the microorganism. The results are grouped in four major sections, according to the experimental aim.
In the first group of experiments, M. antarcticus was constantly exposed to NaCl during the whole fermentation. The goal of these experiments was to establish the level of tolerance to osmotic stress of M. antarcticus and the NaCl levels at which it was still able to produce MEL and CAL-B. This opens the possibility of using saltwater as a substitute for freshwater in the formulation of growth medium.
The second group of experiments consisted of short exposures to salt, to study the rapid changes in physiology of the culture, in order to gain knowledge that could be used in engineering alternative production processes.
In the third set of experiments, M. antarcticus was grown in various non-sterile conditions, in the presence of NaCl.
Finally, to enable the use of MELs for oil spill bioremediation in marine and terrestrial environments, the production of MELs in the presence of NaCl with hydrocarbons as a carbon source was estimated, as well as the ecotoxic effects of the biosurfactant.
3.1. Effects of Continuous Exposure to NaCl on M. antarcticus
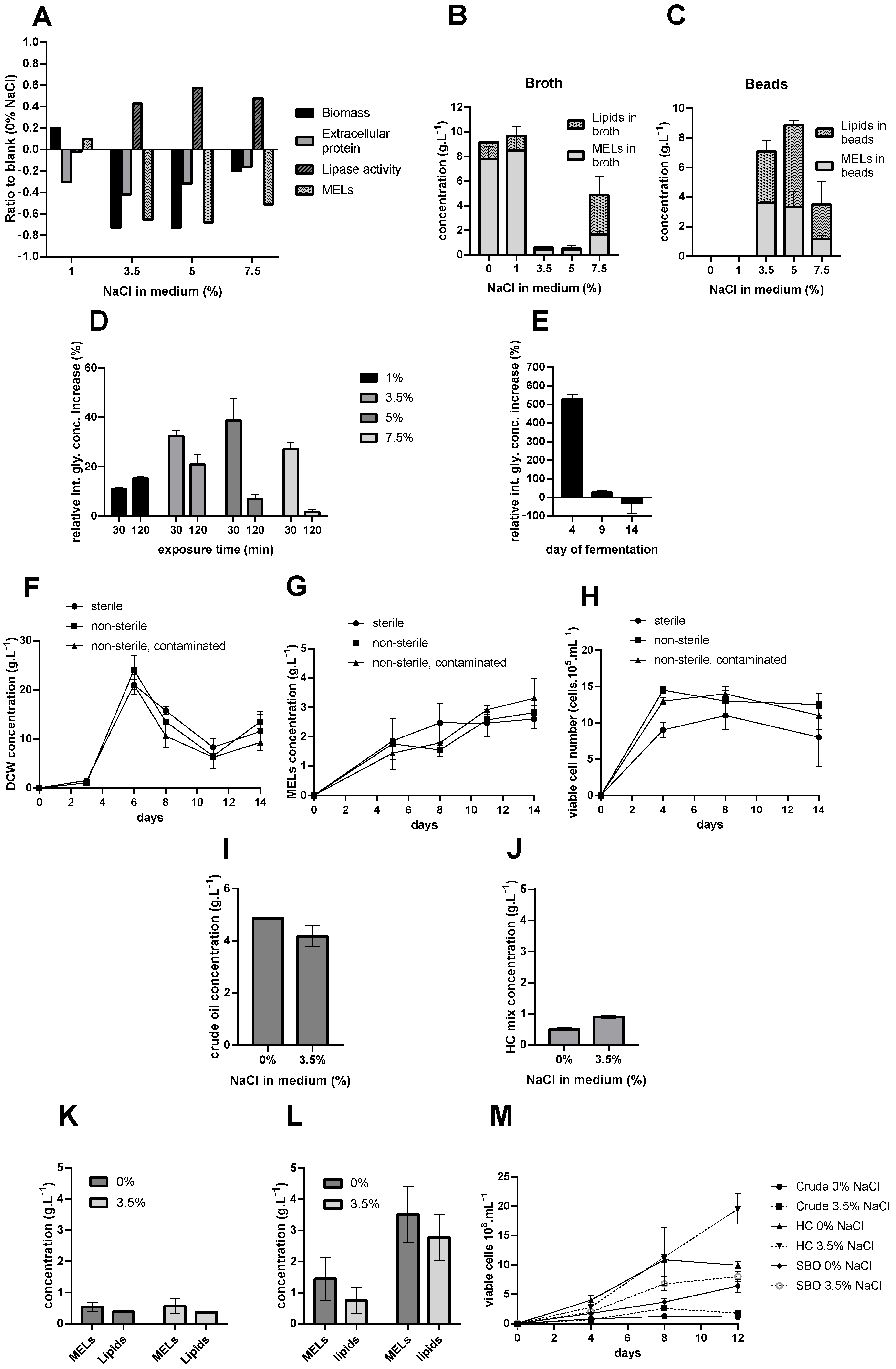
To simplify the manner in which this large set of data is presented, focus is only given to the relation of the data obtained with salt presence in the media compared to the blank cultures (with 0% NaCl). Thus, key values are presented in ratios (%), indicating how the corresponding level of NaCl in the medium deviated from the values for each parameter compared to the blank, as seen in
Figure 1A, while the complete profiles over the cultivation period are presented in
Appendix A Figure A1.
The flasks containing 10% NaCl in the growth medium had no detectable changes in any of the measured parameters, and no observable growth of biomass occurred, meaning that the salt level was high enough to stop all metabolic activity of the cells with added inoculum (as seen in
Figure 2B). This concentration of salt was not used for other experiments, and these results are thus excluded from the remaining part of the paper.
3.2. Effect of NaCl in Growth Media on MEL Production
Finally, to determine whether the production of MEL at an industrial scale with growth media containing salt is promising, we must examine the effect of NaCl on MEL production. The profiles of MEL produced by cells exposed to different levels of salt during growth (
Appendix A) showed high quantities of MEL present in the liquid fraction of the collected cultivation broth for flasks with 0% and 1% of NaCl, with the latter overperforming the blank by roughly 10%. Based on these values alone, it can be concluded that the cultivations with higher NaCl concentration significantly underperformed.
However, due to poor water solubility, the MELs which were not adsorbed to cells had the tendency to form solid beads in the fermentation broth (
Figure 2A), which prevented homogeneous sampling and reliable results being obtained from the liquid fraction of the broth alone. These beads can be harvested using a rough filtration step, for instance with a sieve. They are mainly composed of MELs, however a significant content of free fatty acids and biomass is present. During fermentation with increased titers of MELs, agglomerations form and eventually dissipate.
Flasks with 3.5%, 5%, and 7.5% NaCl content contained significant amounts of solid MEL beads (as seen in
Figure 2B). These beads were recovered using a metal strainer (opening diameter ~1 mm) and extracted with ethyl-acetate twice. MELs and lipid concentrations were then determined for the collected extracts, and their composition was analyzed (
Figure 1B,C and
Table 1).
3.3. Intracellular Glycerol Accumulation by M. antarcticus as a Response to Hyperosmotic Stress
The two main stress response mechanisms observed in
S. cerevisiae and other yeasts are the increase of intracellular glycerol levels and the tendency of cells to flocculate. A study of changes in intracellular glycerol levels as a reaction to the osmotic stress caused by NaCl and sorbitol (another commonly used osmolyte) in
Saccharomyces cerevisiae and
Pichia guillierondii [
16] reported a continuous increase of glycerol levels for up to 40 h after stress agent addition.
To test if M. antarcticus manifested these responses, a set of flasks were prepared, in order to be sacrificed at the end of the fermentation, and the stress response could be quantified as a relative increase in intracellular glycerol concentration.
Intracellular glycerol concentration was measured after exposure of
M. antarcticus to different amounts of salt for 30 and 120 min (
Figure 1D).
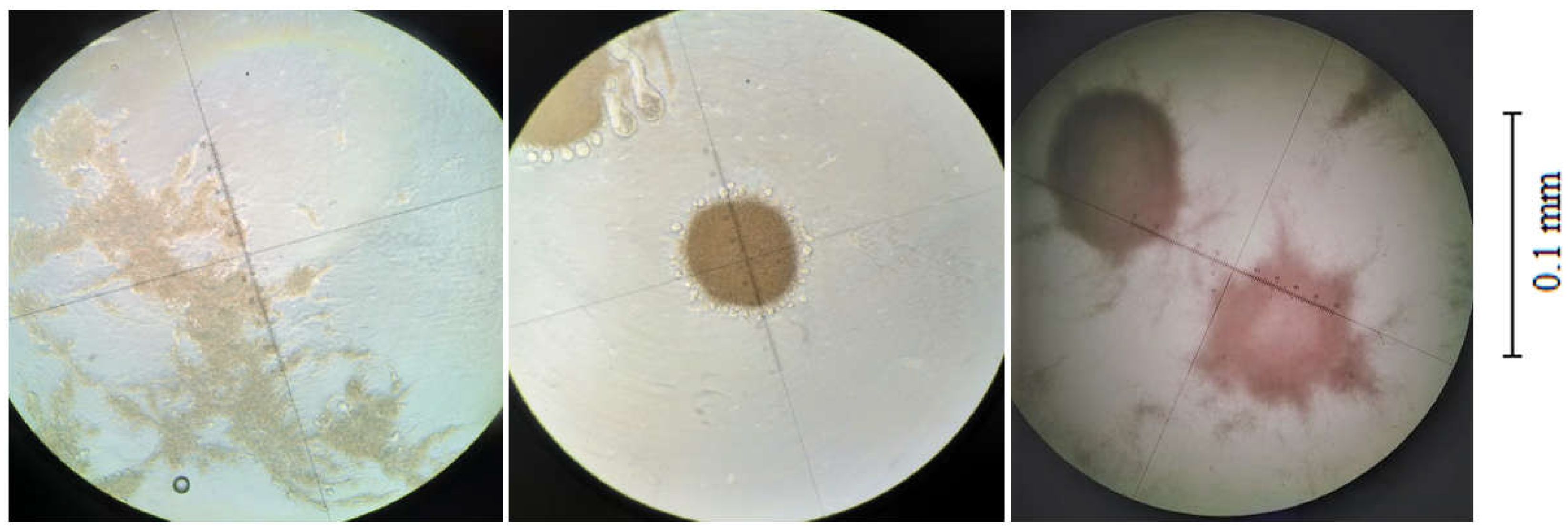
In order to determine what effect a brief exposure to NaCl had on the physiology of
M. antarcticus cells, a test was performed with a sample of the fermentation broth that was mixed with salt for 30 min, with photomicrographs made of cells before and after exposure (
Figure 3).
To test the difference between continuous exposure to NaCl and brief stressor exposure, a set of flasks were incubated. The first group, the blanks, had no salt present in the media, while the others had 3.5% of NaCl. On certain days, flasks were collected and sacrificed, in order to determine the intracellular glycerol concentration in the cultures with NaCl present, relative to the blank (
Figure 1E).
3.4. Non-Sterile MEL Production
In order to test the ability of M. antarcticus to grow in media with NaCl in non-sterile conditions, a set of flasks with 3.5% NaCl were prepared and incubated for 14 days. Initially, growth medium and flasks were sterilized for all runs, to ensure that the initial conditions were identical among different runs. During the experiment, three different conditions were followed:
flask fermentations and sampling were performed in sterile conditions, fermentations were prepared in sterile conditions, sampling/handling in non-sterile conditions.
fermentations prepared in sterile conditions, sampling/handling in non-sterile conditions, with an induced contamination using ~105 E. coli cells after 24 h of fermentation per flask.
this contaminant was selected since it is ubiquitous, mildly resistant to salt, and proven to be a common contaminant. Samples were collected over time, and the biomass (CDW) and viable cell number were measured (
Figure 1F–H).
Viable cells were determined over time by counting the number of colony forming units on a plate after incubation. This allowed clarifying both the composition of the biomass (yeast or bacteria cells) and how the added contaminant affected yeast cell viability (
Figure 1H).
3.5. Crude Oil and Hydrocarbon Tests
To estimate the dual marine bioremediation effect (the hydrocarbon-consuming microorganism and the petroleum-dispersive properties of the biosurfactant),
M. antarcticus was grown on media with and without NaCl present, following the previously described protocol, where the vegetable oil was substituted by 5 g·L
−1 of crude oil and a mix of linear alkanes and added on day 4. The residual MELs, FFAs, crude oil, and alkanes were extracted from the fermentation broth and their concentrations were analyzed. The results for residual hydrocarbons are presented in
Figure 1I,J.
The results for MEL and FFA concentrations extracted from flasks after 12 days of cultivation are presented in
Figure 1K,L.
To further study the toxicity effects of the hydrocarbons on the cells, viable cell concentrations were determined and are presented in
Figure 1M.
3.6. Ecotoxicity of MELs
MELs potential role in bioremediation has already been hypothesized in the literature [
11]. Here, we established that
M. antarcticus has the potential to produce MELs in the presence of a marine level of NaCl. However, before using MELs for bioremediation, ecotoxicity concerns need to be addressed. In order to estimate MELs toxic effects in a marine setting, a model organism for marine ecotoxicity, the brine shrimp
Artemia franciscana, was used. Cysts were immersed in solutions of several compounds with a range of concentrations, and IC50 values were estimated based on cyst mortality. MELs were tested in their crude form, as extracted from the fermentation broth, as well as using a cell-free supernatant containing 2.32 g·L
−1 of MELs. To compare its performance with other glycolipid biosurfactants, tests were also performed using sophorolipids and rhamnolipids in crude form. Corexit, a commercial chemical product used for bioremediation after marine oil spills, was also tested. Pure marine salt water and potassium dichromate were used as the negative and positive blank, respectively. Data for IC 50 values are presented in
Table 2.
4. Discussion
The results of this study of the effect of NaCl on
M. antarcticus biomass growth (
Figure 1A) are consistent with the effects reported in literature for
S. cerevisiae [
17], indicating that the increased levels of NaCl in the medium affected
M. antarcticus by negatively affecting biomass development and extending the lag-period. All cultures performed worse than the blank (with 0% NaCl) in terms of DCW, except the culture with 1% NaCl, which was better than the blank, by roughly 20%.
However, one of the main preconditions for MEL production is that there is a sufficient amount of active lipases. They have a key importance in MEL production, as they play a role in the consumption of lipid-based substrates, which is one of the major steps in the biotransformation chain towards MEL generation. The results for extracellular protein and lipase activity (
Figure 1A), as well as the specific activity of the lipases calculated from these two values (
Appendix A), show some difference among the various growth conditions. Although the increasing levels of salt in the media decreased the amount of extracellular protein produced, the activities of the lipases in cultures in the presence of 3.5 and 5% NaCl were over 40% higher than the blank. Although the growth and basic metabolic activity of the cells was hindered by the presence of salt in the media, it is common for yeast and fungal strains to react to stressors with stimulated extracellular protein secretion [
18]. Furthermore, osmotic stress causes a drop in gene expression for proteins which have a role in amino acid metabolism, cell wall maintenance, nucleosome structure, DNA synthesis, and nucleotide metabolism [
6], as they play a key role in cell proliferation. In hyperosmotic stress conditions, cells tend to promote the production of osmoregulatory and other enzymes, which can increase survival rates under such conditions. This may be the explanation for the unexpectedly high lipase secretion. Although the overall metabolism of the strain might be affected, the increase of lipase production could be a cell response to the stress caused by increased osmolarity.
As for the profiles of the MEL concentrations, some variance was present due to inhomogeneous sampling in the presence of solids (
Appendix A Figure A1). However, this bead appearance did not depend solely on the profile of the MELs in the broth. Based on MEL and lipid profiles (
Appendix A Figure A1), as well as data from
Figure 1B,C, it seems that free fatty acids played an integral role in the formation of these beads, as they were present in the beads in a significant quantity. The presence of beads rich in MELs during the final stages of fermentation is beneficial, as this would improve the efficiency and sustainability of the downstream processing through simpler bead harvesting and lower organic solvent requirements for MEL separation using liquid–liquid extractions.
One mechanism of stress response reported in hypersaline fermentation conditions in yeasts is the rapid accumulation of intracellular glycerol. The results of the experiments where cells were exposed to short-term osmotic stress (
Figure 1D) showed that the intracellular glycerol accumulation was higher in
M. antarcticus samples exposed to higher concentrations of salt, while such accumulation ceased after some time, dropping significantly by the end of two hours of exposure. Either the cells started to adapt to the osmotic stress, recovered, and released some of the produced glycerol, or the cells response mechanisms were exhausted and the high-osmolarity glycerol (HOG) pathway was reversed. The reaction was similar in all cases, with a glycerol spike in the first sample measured and a later drop, except for the flask with 1%, where cells responded more slowly, continuously accumulating glycerol over the course of 2 h.
In this work, the cultures continuously exposed to osmotic stress had significantly higher intracellular glycerol concentrations (
Figure 1E). However, in the later stages of the fermentation, probably due to slower growth and compromised metabolic activity due to constant exposure to the stressor, the culture with salt present had less accumulated glycerol inside the cells. This further validates that the presence of NaCl negatively influenced the basic metabolism of the yeast, since it weakened the ability of the cells to respond to osmotic stress with time.
Physiological changes in
M. antarcticus caused my hyperosmotic stress were observed in the form of rapid cell flocculation (
Figure 3). Other studies have reported that NaCl causes disruption of the floccules of brewer’s yeast [
19], due to the antagonistic effect of Na ions compared to Ca ions, and this phenomenon is widely used in analytical practice for deflocculation prior to analysis [
20]. Namely, sodium ions tend to substitute calcium, which normally enables the formation of ionic bridges and causes cell flocculation with great efficiency [
21]. However, this was not observed with
M. antarcticus, as the floccules did not completely dissipate after exposure to EDTA, which should have inhibited the effect of the ions on the cells. This means that the cells did not aggregate due to salt bridges alone, but due to other additional stimuli.
The use of marine-level saline water for medium preparation was studied for production of MELs in non-sterile conditions. From the relevant parameters presented in
Figure 1F–H, it can be observed that there was no significant difference in biomass concentration and viable cell number among the different conditions tested. The presence of the intentionally introduced, or any other, contaminant did not compromise the
M. antarcticus cultivation. Petri plates with samples from all flasks showed only colonies of
M. antarcticus, with the set of flasks with the induced contamination not showing any increase of contaminant colony number over time; i.e., the contaminant culture did not develop. Furthermore, the MEL production was not affected.
The fermentations performed using hydrocarbons as a carbon source explored the possible dual bioremediation effect: hydrocarbon consumed and removed from a contaminated area and the in-situ MEL production. The hydrocarbon consumption was not significantly different for cultures cultivated in media with 0% and 3.5% of NaCl. These results indicate that the presence of salt did not inhibit the culture metabolism. When using crude oil instead of pure hydrocarbons, the MELs and lipid concentration were similar in both conditions and the presence of salt did not inhibit the MELs production. Nevertheless, the MELs production was higher when using linear alkanes. The complex composition, impurities, and toxic compounds present in crude oil, such as aromatic compounds, asphaltenes, and heavy metals, which are not present in the alkane mix, had inhibitory effects on the culture.
Hydrocarbons and the use of salt were shown to have effects on cell proliferation (
Figure 1M). It seems that stress, caused by the salinity and moderate hydrocarbon toxicity, had a positive effect on cell proliferation. The effect was, however, opposite when crude oil was used, and the intense toxicity inhibited culture development. Although stress normally negatively affects yeast cell proliferation, Tamás and Hohmann [
22] reported that the cell proliferation and regulation of protein kinase A are tightly connected in
S. cerevisiae. Although the induction of cell proliferation caused by osmotic and toxic stress in
M. antarcticus seems evident from the presented data, this requires further investigation.
Finally, the ecotoxic effect of MELs was estimated using brine shrimp, a model organism for marine ecotoxicity determination,
Artemia franciscana. The chemical oil dispersant Corexit proved to be highly toxic to the brine shrimp, based on the IC50 values presented in
Table 2. Among the tested glycolipid biosurfactants, the MELs outperformed sophorolipids and rhamnolipids, whose toxicity values matched those reported in the literature [
23,
24,
25].
5. Conclusions
Moesziomyces antarcticus was grown in media with different concentrations of NaCl present, biological samples collected at different stages of the fermentation were exposed to various amounts of osmotic stressor (NaCl), and different tests and analyses were performed, in order to determine cell reactions.
First, it can be concluded that the presence of NaCl negatively affects the growth and basic metabolism (such as sugar and lipid consumption) of Moesziomyces spp. Conversely, lipase production was not affected, or even induced, with the presence of NaCl.
Osmotic stress responses, such as intracellular glycerol accumulation and surface changes, which cause agglomeration, as reported for Saccharomyces cerevisiae and other yeasts, were also seemingly observed in M. antarcticus.
NaCl promoted formation of MEL-rich beads. The physiological effects of NaCl probably caused surface changes to the cells, and the MELs adhesion to the cell walls was lowered, which promoted bead formation, which can bring advantages in downstream processing. Salt can thus be considered an interesting parameter to study in MEL bead formation and disappearance in Moesziomyces spp. cultivation.
MEL was produced in both non-sterile conditions in 3.5% NaCl medium (equivalent to seawater salt levels), and in the same medium spiked with a common contaminating microorganism, which prevented yeast development and MEL production in the regular medium (with no NaCl). This could enable production in unsterile conditions, thus contributing to the final goal of lowering production costs of CAL-B and MELs, increasing the sustainability of the production process, and making MELs a more competitive biosurfactant on the market.
In addition, M. antarcticus’ ability to successfully metabolize hydrocarbons and produce MELs in saline water was explored. Although crude oil was shown to be toxic to the culture, MELs were successfully produced from a mix of linear alkanes, especially in 3.5% NaCl medium. Osmotic stress was observed as promoting cell proliferation, which is unusual compared to data from the literature relating osmotic stress with proliferation arrest in other yeasts.
Finally, the ecotoxicity of the biosurfactant was explored using the model marine organisms Artemia franciscana. This was tested with MELs, which was compared to other biosurfactants and a commercial oil dispersant used for bioremediation. Here, MELs proved to be less toxic than the other biosurfactants and commercial oil dispersants, with a slightly lower toxicity observed in MELs in supernatant form. These results indicate that MELs can be safely used for marine oil spill bioremediation.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}