
Exploratory Study of Sex Identification for Chicken Embryos Based on Blood Vessel Images and Deep Learning


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.1.1. Materials and Instruments
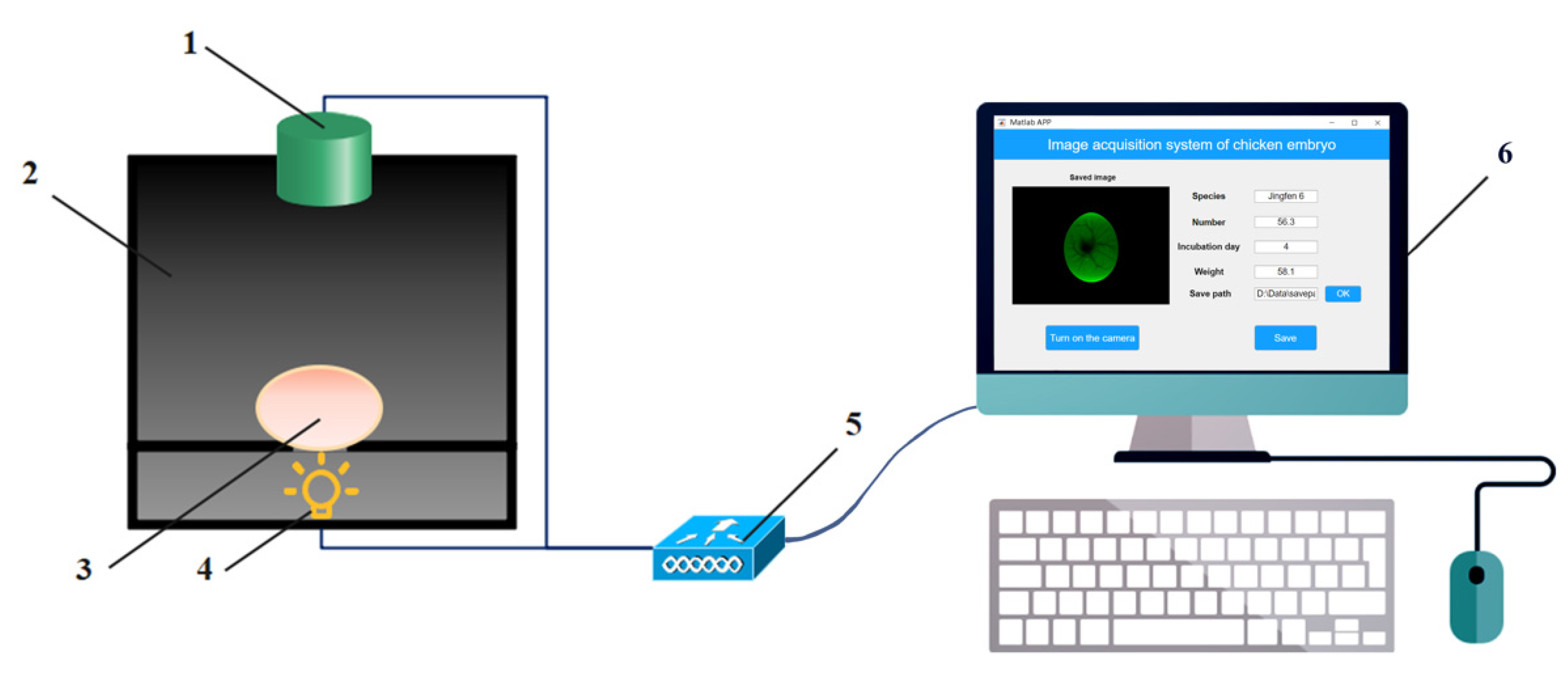
2.1.2. Image Acquisition System
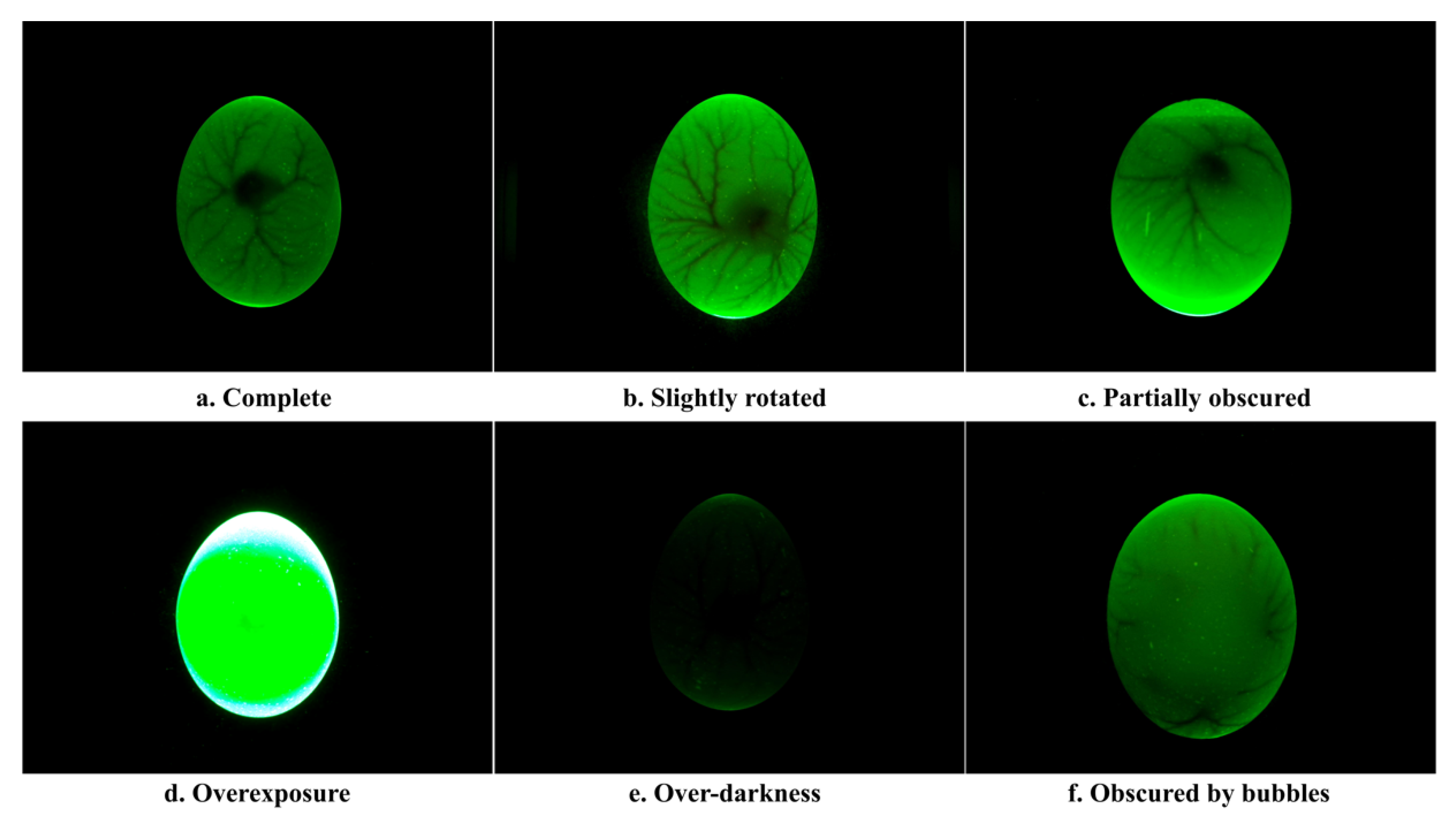
2.1.3. Data Acquisition Experiment
2.2. Dataset Construction
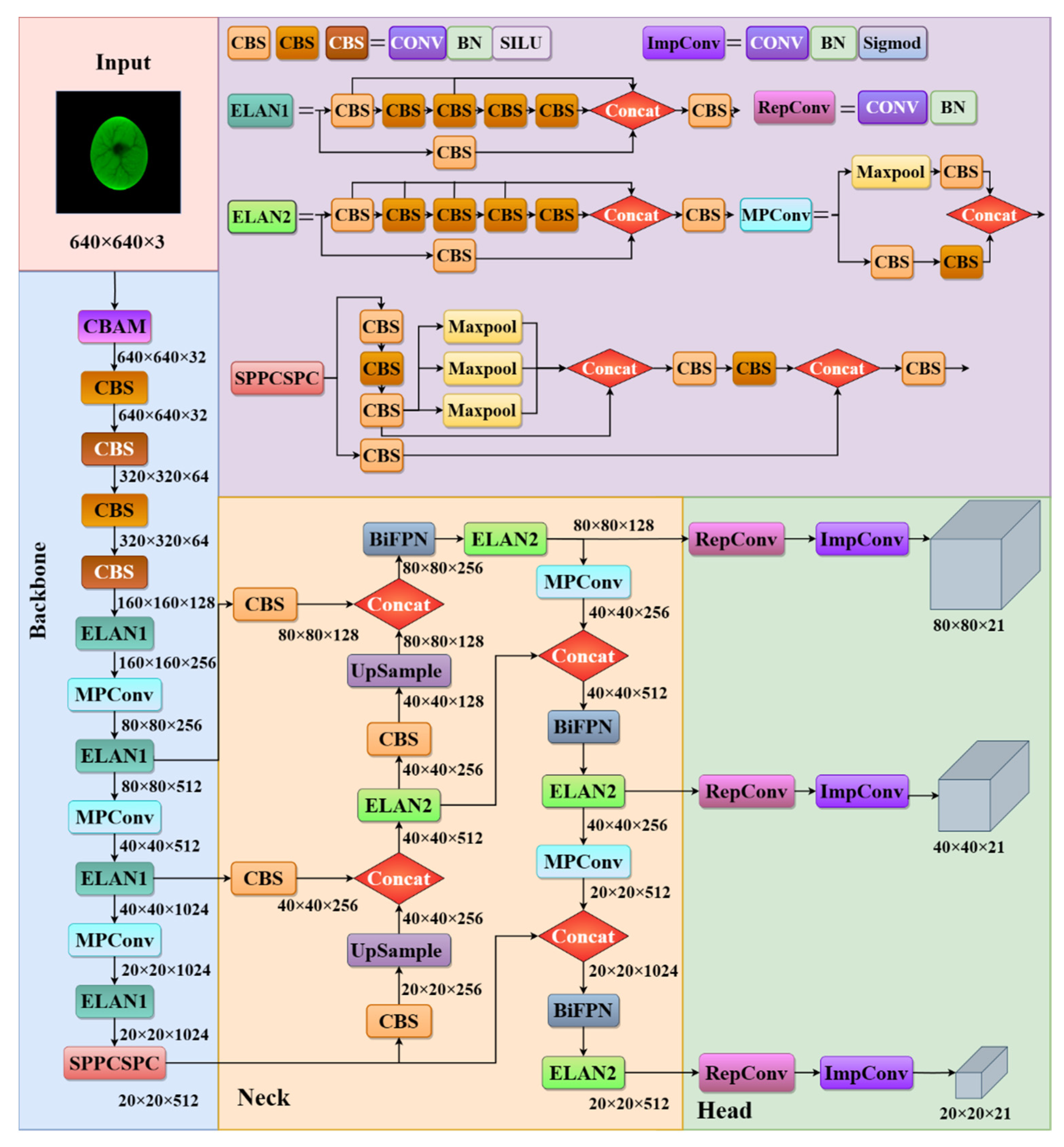
2.3. YOLOv7 Deep Learning Network
2.4. Improvement of the YOLOv7 Algorithm
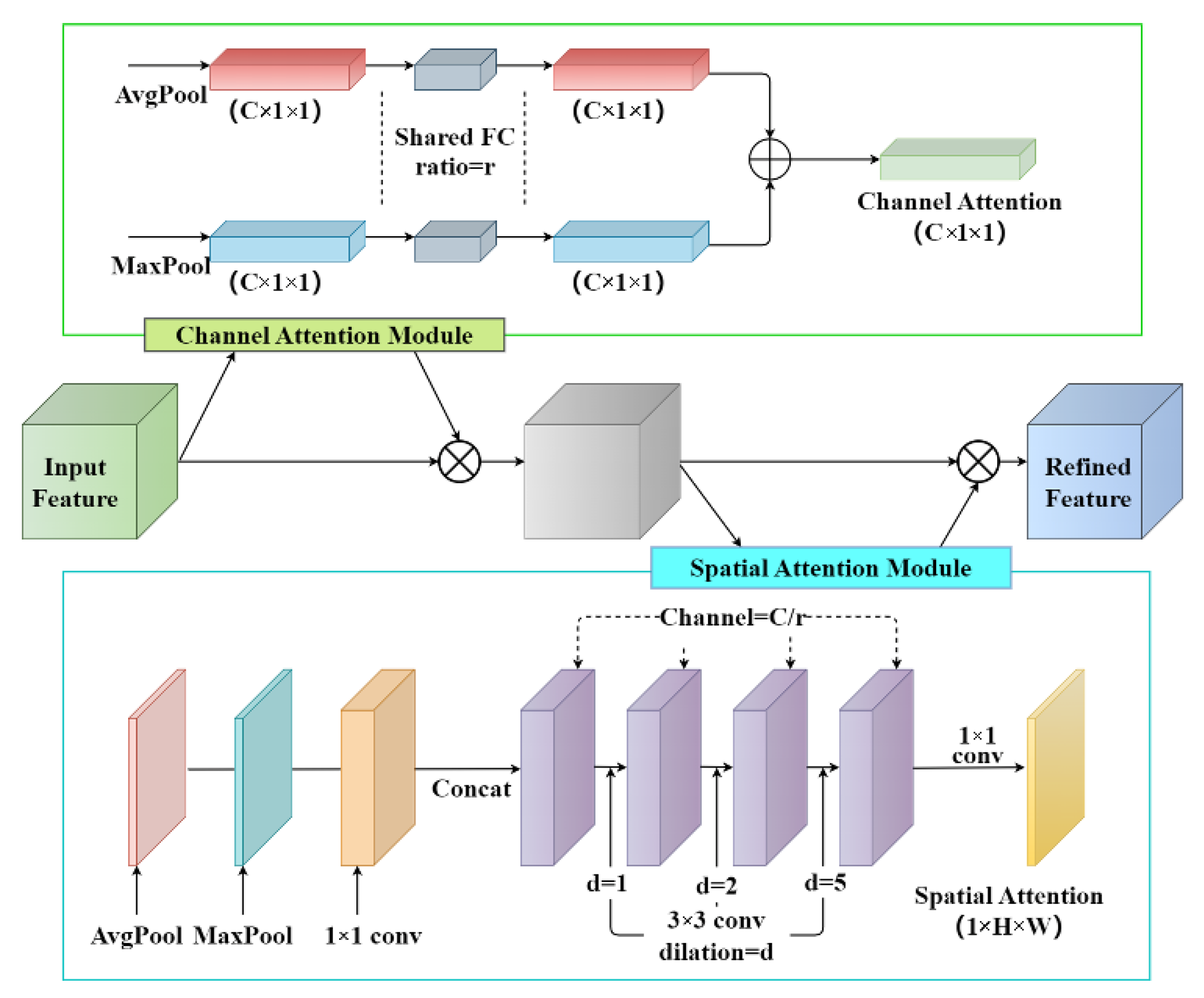
2.4.1. Backbone Attention Module
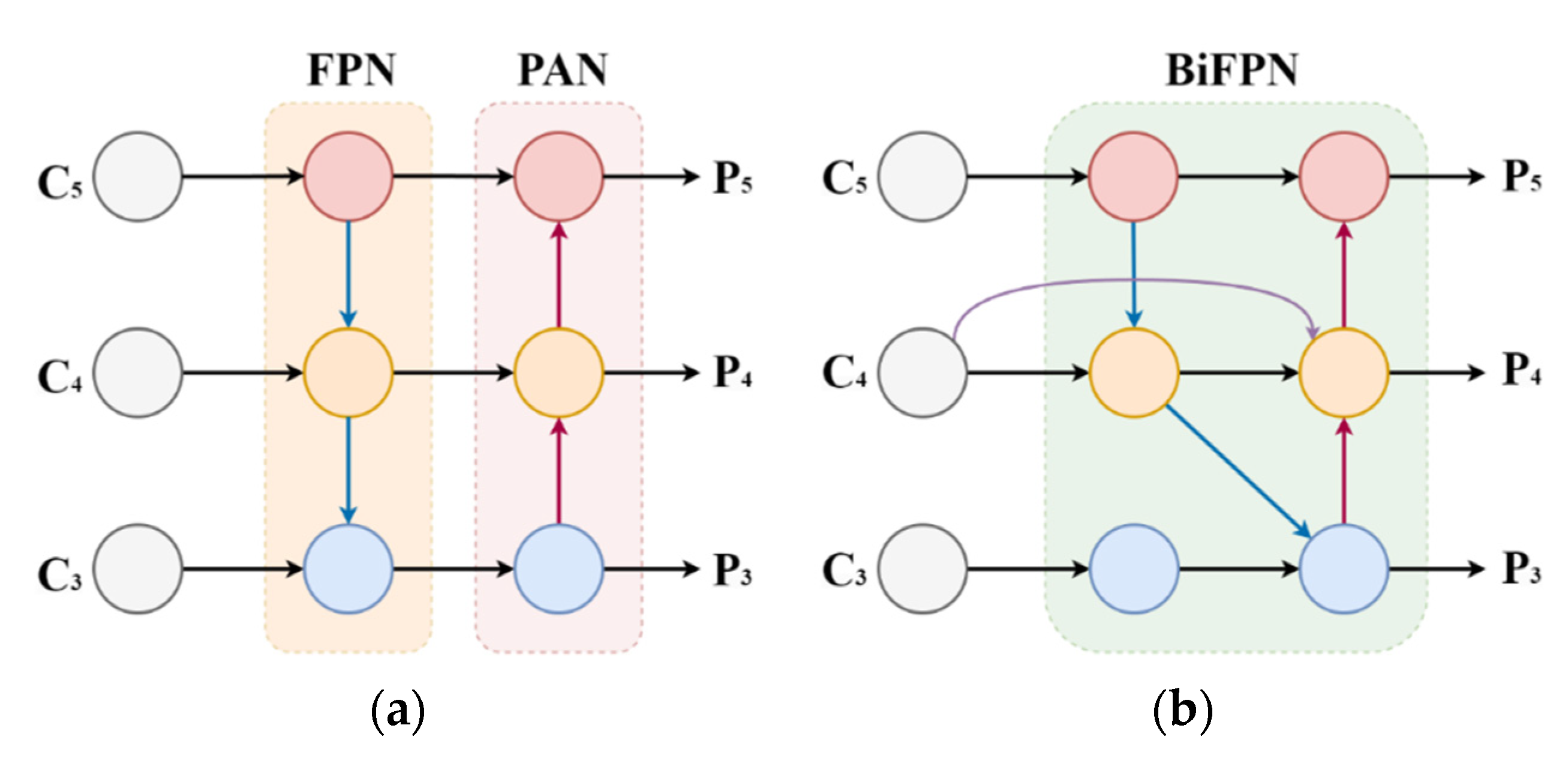
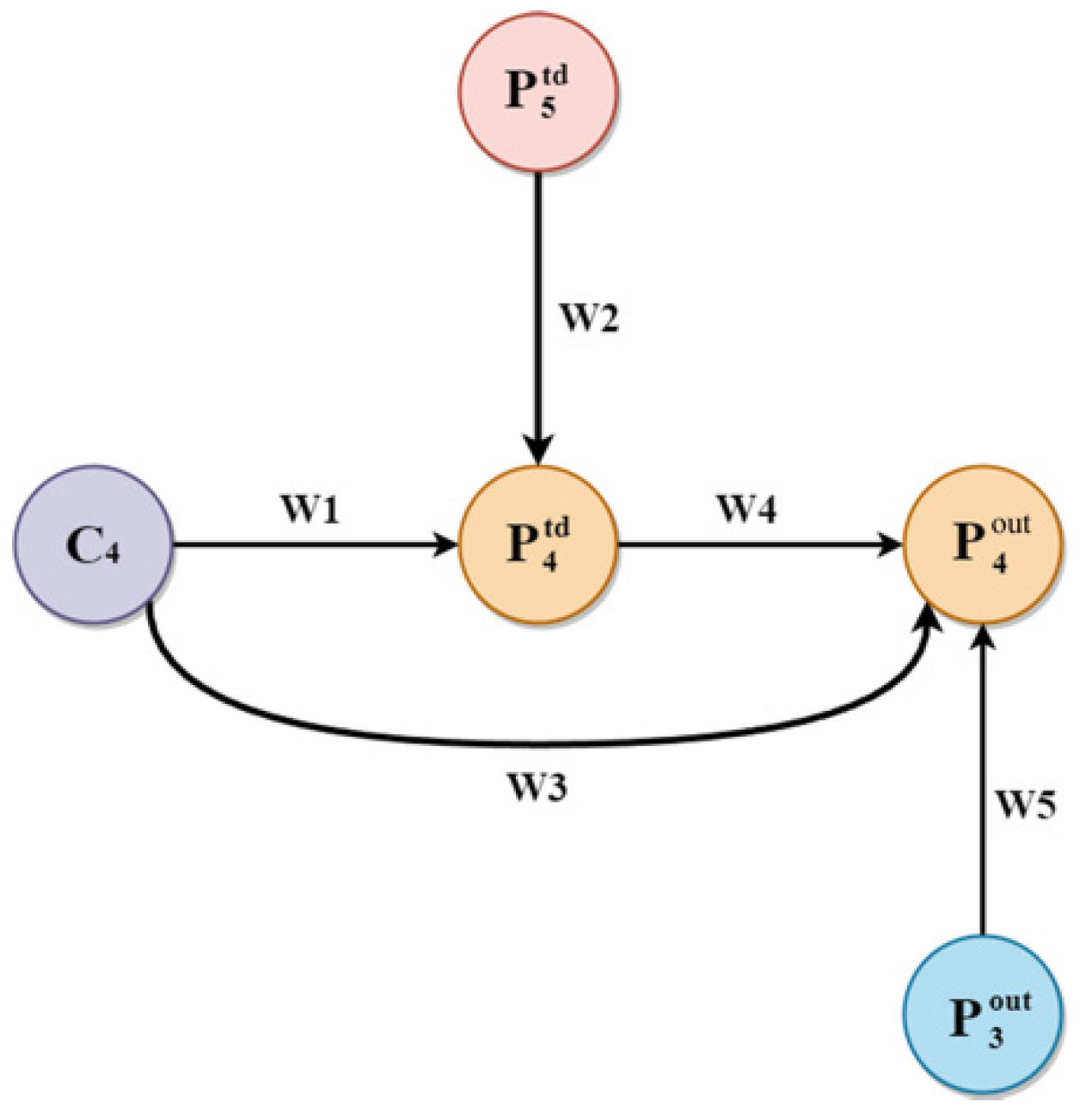
2.4.2. Multi-Scale Features Fusion
2.4.3. Loss Algorithm
2.5. Experimental Setup and Evaluation Metrics
3. Results and Discussion
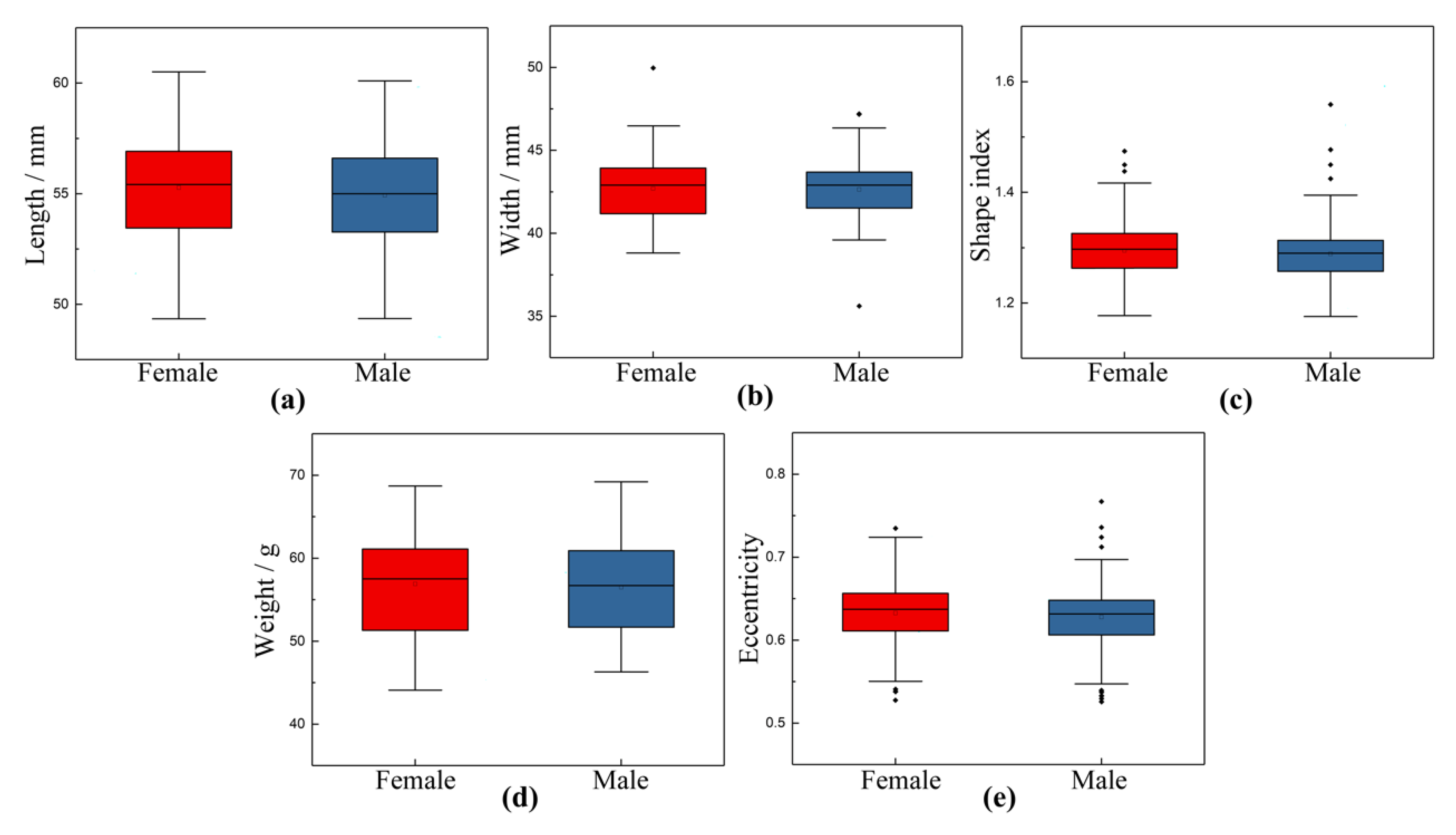
3.1. Relationship between External Morphological Characteristics and Sex of Chicken Eggs
3.2. Algorithm Performance Evaluation
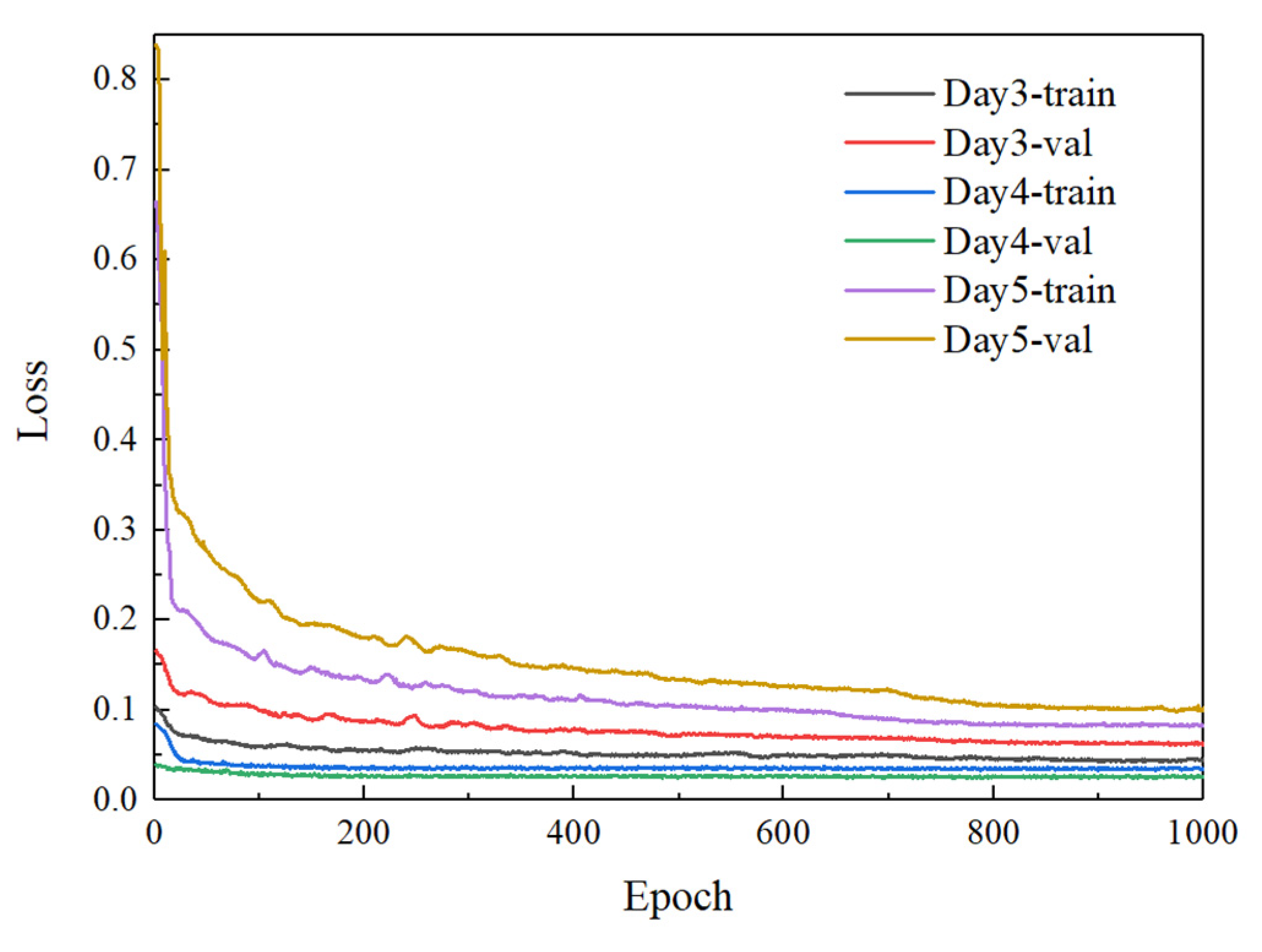
3.2.1. Results of Different Incubation Days
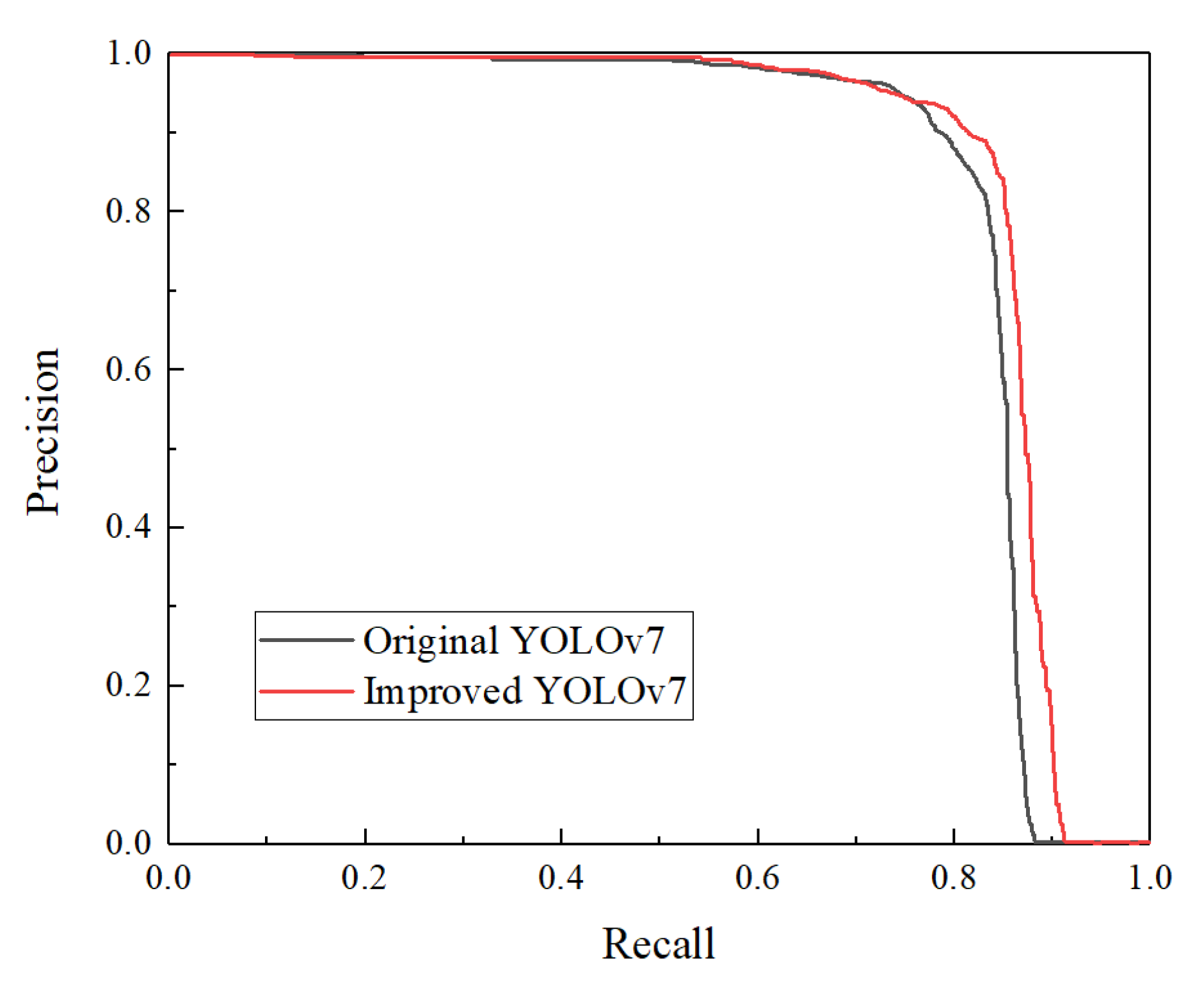
3.2.2. Impact of YOLOv7 Improvements on Sex Identification of Chicken Eggs
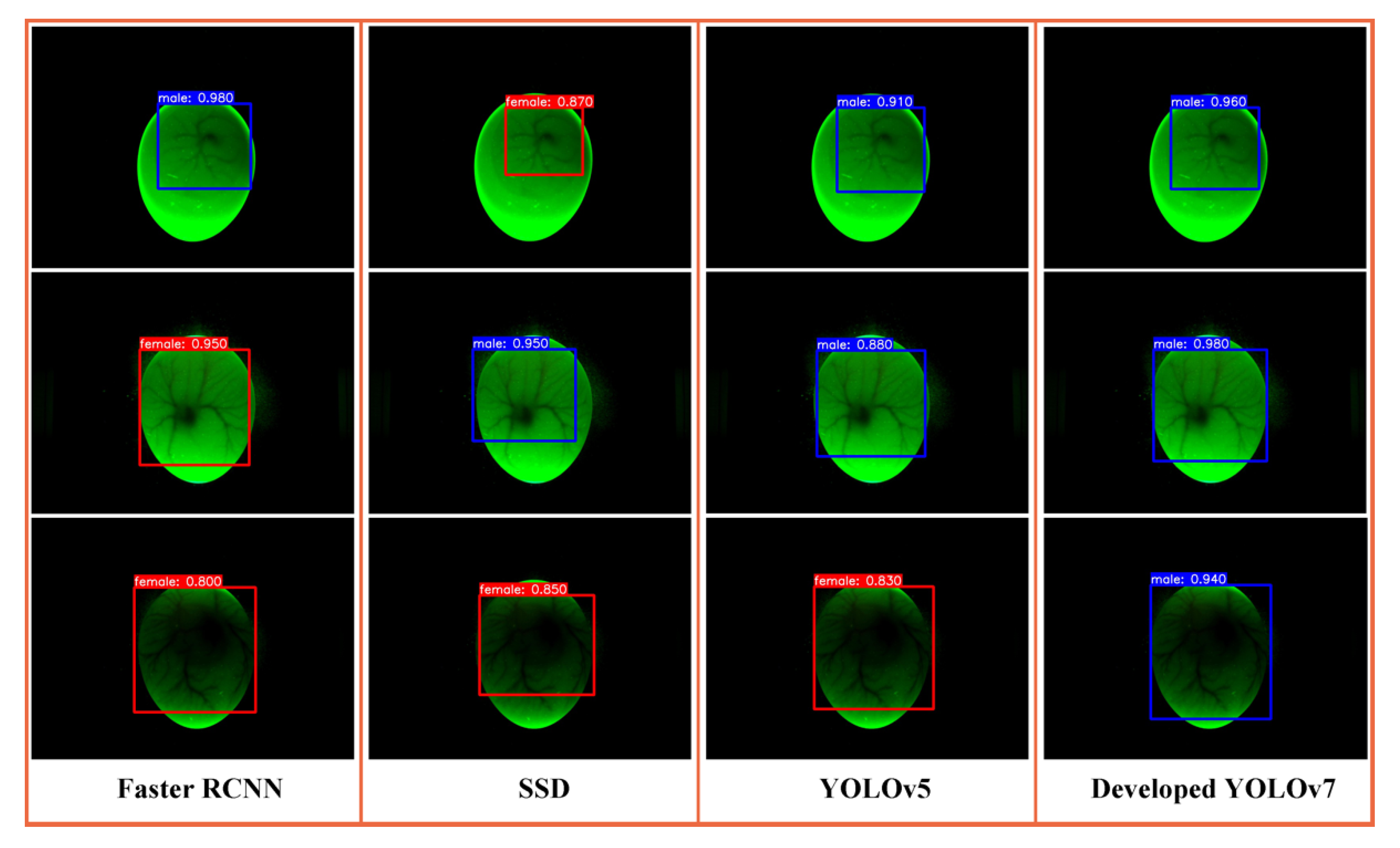
3.3. Comparison of Different Object Detection Algorithms
3.4. Comparison with Existing Research
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krautwald-Junghanns, M.E.; Cramer, K.; Fischer, B.; Förster, A.; Galli, R.; Kremer, F.; Mapesa, E.U.; Meissner, S.; Preisinger, R.; Preusse, G.; et al. Current approaches to avoid the culling of day-old male chicks in the layer industry, with special reference to spectroscopic methods. Poult. Sci. 2018, 97, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.; Bartels, T.; Stelling, A.; Krautwald-Junghanns, M.-E.; Fuhrmann, H.; Sablinskas, V.; Koch, E. Gender determination of fertilized unincubated chicken eggs by infrared spectroscopic imaging. Anal. Bioanal. Chem. 2011, 400, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Steiner, G.; Koch, E.; Krautwald-Junghanns, M.E.; Bartels, T. Method and Device for Determining the Sex of Fertilized, Non-Incubated Bird Eggs. U.S. Patent 8,624,190, 7 January 2014. [Google Scholar]
- Galli, R.; Preusse, G.; Uckermann, O.; Bartels, T.; Krautwald-Junghanns, M.E.; Koch, E.; Steiner, G. In Ovo Sexing of Domestic Chicken Eggs by Raman Spectroscopy. Anal. Chem. 2016, 88, 8657–8663. [Google Scholar] [CrossRef] [PubMed]
- Galli, R.; Koch, E.; Preusse, G.; Schnabel, C.; Bartels, T.; Krautwald-Junghanns, M.-E.; Steiner, G. Contactless in ovo sex determination of chicken eggs. Curr. Dir. Biomed. Eng. 2017, 3, 131–134. [Google Scholar] [CrossRef]
- Galli, R.; Preusse, G.; Schnabel, C.; Bartels, T.; Cramer, K.; Krautwald-Junghanns, M.E.; Koch, E.; Steiner, G. Sexing of chicken eggs by fluorescence and Raman spectroscopy through the shell membrane. PLoS ONE 2018, 13, e0192554. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Zhang, W.; Yu, M.; Sun, Y.; Gu, X.; Ma, L.; Li, Z.; Hu, P.; Tu, K. Gender determination of early chicken hatching eggs embryos by hyperspectral imaging. Trans. Chin. Soc. Agric. Eng. 2016, 32, 181–186. [Google Scholar]
- Göhler, D.; Fischer, B.; Meissner, S. In-ovo sexing of 14-day-old chicken embryos by pattern analysis in hyperspectral images (VIS/NIR spectra): A non-destructive method for layer lines with gender-specific down feather color. Poult. Sci. 2017, 96, 1–4. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Q.; Ma, M.; Xiao, S.; Shi, H. Non-Destructive Detection of Male and Female Information of Early Duck Embryos Based on Visible/Near Infrared Spectroscopy and Deep Learning. Spectrosc. Spectr. Anal. 2021, 41, 1800–1805. [Google Scholar]
- Alin, K.; Fujitani, S.; Kashimori, A.; Suzuki, T.; Ogawa, Y.; Kondo, N. Non-invasive broiler chick embryo sexing based on opacity value of incubated eggs. Comput. Electron. Agric. 2019, 158, 30–35. [Google Scholar] [CrossRef]
- Rahman, A.; Syduzzaman, M.; Khaliduzzaman, A.; Fujitani, S.; Kashimori, A.; Suzuki, T.; Ogawa, Y.; Kondo, N. Nondestructive sex-specific monitoring of early embryonic development rate in white layer chicken eggs using visible light transmission. Br. Poult. Sci. 2020, 61, 209–216. [Google Scholar] [CrossRef]
- Xiang, X.; Wang, Y.; Yu, Z.; Ma, M.; Zhu, Z.; Jin, Y. Non-destructive characterization of egg odor and fertilization status by SPME/GC-MS coupled with electronic nose. J. Sci. Food Agric. 2019, 99, 3264–3275. [Google Scholar] [CrossRef]
- Corion, M.; De Ketelaere, B.; Hertog, M.; Lammertyn, J. Profiling the emission of volatile organic compounds from chicken hatching eggs in the first half of incubation. In Proceedings of the IFRG eMeeting 2021, Online, 14–15 October 2021. [Google Scholar]
- Xiang, X.; Hu, G.; Jin, Y.; Jin, G.; Ma, M. Nondestructive characterization gender of chicken eggs by odor using SPME/GC-MS coupled with chemometrics. Poult. Sci. 2022, 101, 101619. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Redmann, T. Approaches to determine the sex prior to and after incubation of chicken eggs and of day-old chicks. Worlds. Poult. Sci. J. 2008, 64, 391–399. [Google Scholar] [CrossRef]
- Tang, Y.; Hong, Q.; Wang, Q.; Zhu, Z. Sex identification of chicken eggs based on blood line texture features and GA-BP neural network. J. Huazhong Agric. Univ. 2018, 37, 130–135. [Google Scholar]
- Zhu, Z.; Tang, Y.; Hong, Q.; Huang, P.; Wang, Q.; Ma, M. Female and male identification of early chicken embryo based on blood line features of hatching egg image and deep belief networks. Trans. Chin. Soc. Agric. Eng. 2018, 34, 197–203. [Google Scholar]
- Zhu, Z.; Zifan, Y.; Yuting, H.; Kai, Y.; Qiaohua, W.; Meihu, M. Gender identification of early chicken embryo based on RF-DS information fusion of spectroscopy and machine vision. Trans. Chin. Soc. Agric. Eng. 2022, 38, 308–315. [Google Scholar]
- Li, W.; Feng, X.S.; Zha, K.; Li, S.; Zhu, H.S. Summary of Target Detection Algorithms. J. Phys. Conf. Ser. 2021, 1757, 012003. [Google Scholar] [CrossRef]
- Girshick, R.; Donahue, J.; Darrell, T.; Malik, J. Rich Feature Hierarchies for Accurate Object Detection and Semantic Segmentation. In Proceedings of the 2014 IEEE Conference on Computer Vision and Pattern Recognition, Columbus, OH, USA, 23–28 June 2014; pp. 580–587. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Spatial pyramid pooling in deep convolutional networks for visual recognition. IEEE Trans. Pattern Anal. Mach. Intell. 2015, 37, 1904–1916. [Google Scholar] [CrossRef] [Green Version]
- Girshick, R. Fast R-CNN. In Proceedings of the IEEE International Conference on Computer Vision (ICCV), Santiago, Chile, 7–13 December 2015; pp. 1440–1448. [Google Scholar]
- Ren, S.; He, K.; Girshick, R.; Sun, J. Faster R-CNN: Towards Real-Time Object Detection with Region Proposal Networks. IEEE Trans. Pattern Anal. Mach. Intell. 2017, 39, 1137–1149. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Anguelov, D.; Erhan, D.; Szegedy, C.; Reed, S.; Fu, C.-Y.; Berg, A.C. SSD: Single Shot MultiBox Detector BT-Computer Vision–ECCV 2016; Leibe, B., Matas, J., Sebe, N., Welling, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 21–37. [Google Scholar]
- Redmon, J.; Divvala, S.; Girshick, R.; Farhadi, A.; Impiombato, D.; Giarrusso, S.; Mineo, T.; Catalano, O.; Gargano, C.; La Rosa, G.; et al. You Only Look Once: Unified, Real-Time Object Detection. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2016, 794, 185–192. [Google Scholar]
- Bochkovskiy, A.; Wang, C.-Y.; Liao, H.-Y.M. YOLOv4: Optimal speed and accuracy of object detection. arXiv 2020, arXiv:2004.10934. [Google Scholar]
- Wang, C.Y.; Bochkovskiy, A.; Liao, H.Y.M. Scaled-yolov4: Scaling cross stage partial network. In Proceedings of the IEEE/cvf Conference on Computer Vision and Pattern Recognition, Nashville, TN, USA, 20–25 June 2021; pp. 13024–13033. [Google Scholar] [CrossRef]
- Long, X.; Deng, K.; Wang, G.; Zhang, Y.; Dang, Q.; Gao, Y.; Shen, H.; Ren, J.; Han, S.; Ding, E.; et al. PP-YOLO: An effective and efficient implementation of object detector. arXiv 2020, arXiv:2007.12099. [Google Scholar]
- Ge, Z.; Liu, S.; Wang, F.; Li, Z.; Sun, J. YOLOX: Exceeding YOLO series in 2021. arXiv 2021, arXiv:2107.08430. [Google Scholar]
- Paulguerrie Ultralytics.YOLOv5. Available online: https://github.com/ultralytics/yolov5 (accessed on 19 September 2022).
- Wang, C.-Y.; Yeh, I.-H.; Liao, H.-Y.M. You Only Learn One Representation: Unified Network for Multiple Tasks. arXiv 2021, arXiv:2105.04206. [Google Scholar]
- Wang, C.; Bochkovskiy, A.; Liao, H. YOLOv7: Trainable bag-of-freebies sets new stateof-the-art for real-time object detectors. arXiv 2022, arXiv:2207.02696. [Google Scholar]
- Shafey, T.M.; Al-Mohsen, T.H. Embryonic growth, hatching time and hatchability performance of meat breeder eggs incubated under continuous green light. Asian-Australas. J. Anim. Sci. 2002, 15, 1702–1707. [Google Scholar] [CrossRef]
- Rozenboim, I.; Piestun, Y.; Mobarkey, N.; Barak, M.; Hoyzman, A.; Halevy, O. Monochromatic light stimuli during embryogenesis enhance embryo development and posthatch growth. Poult. Sci. 2004, 83, 1413–1419. [Google Scholar] [CrossRef]
- Sobolewska, A.; Elminowska-Wenda, G.; Bogucka, J.; Szpinda, M.; Walasik, K.; Dankowiakowska, A.; Jóźwicki, W.; Wiśniewska, H.; Bednarczyk, M. The effect of two different green lighting schedules during embryogenesis on myogenesis in broiler chickens. Eur. Poult. Sci. 2019, 83, 1–14. [Google Scholar] [CrossRef]
- Abdulateef, S.; Farhan, S.M.; Awad, M.M.; Mohammed, T.T.; Al-Ani, M.Q.; Al-Hamdani, A. LBMON172 Impact Of Photoperiodic Green Light During Incubation On Change Hormone In The Embryo Of Chicken. J. Endocr. Soc. 2022, 6, A470–A471. [Google Scholar] [CrossRef]
- Woo, S.; Park, J.; Lee, J.Y.; Kweon, I.S. CBAM: Convolutional Block Attention Module; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Pang, R.; Le, Q.V. EfficientDet: Scalable and efficient object detection. In Proceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition (CVPR), Seattle, WA, USA, 13–19 June 2020; pp. 10778–10787. [Google Scholar] [CrossRef]
- He, J.; Erfani, S.; Ma, X.; Bailey, J.; Chi, Y.; Hua, X.S. Alpha-IoU: A Family of Power Intersection over Union Losses for Bounding Box Regression. Adv. Neural Inf. Process. Syst. 2021, 24, 20230–20242. [Google Scholar]
- Nyalala, I.; Okinda, C.; Kunjie, C.; Korohou, T.; Nyalala, L.; Chao, Q. Weight and volume estimation of poultry and products based on computer vision systems: A review. Poult. Sci. 2021, 100, 101072. [Google Scholar] [CrossRef]
- Yilmaz-Dikmen, B.; Dikmen, S. A morphometric method of sexing white layer Eggs. Rev. Bras. Cienc. Avic. 2013, 15, 203–210. [Google Scholar] [CrossRef]
- Mappatao, G. Duck Egg Sexing by Eccentricity Determination Using Image Processing. J. Telecommun. Electron. Comput. Eng. 2018, 10, 71–75. [Google Scholar]
- Dioses, J.L.; Medina, R.P.; Fajardo, A.C.; Hernandez, A.A. Performance of Classification Models in Japanese Quail Egg Sexing. In Proceedings of the 2021 IEEE 17th International Colloquium on Signal Processing & Its Applications (CSPA), Langkawi, Malaysia, 5–6 March 2021; pp. 29–34. [Google Scholar] [CrossRef]
- Pardo, J.I.S.; González, F.J.N.; Ariza, A.G.; Arbulu, A.A.; Jurado, J.M.L.; Bermejo, J.V.D.; Vallejo, M.E.C. Traditional sexing methods and external egg characteristics combination allow highly accurate early sex determination in an endangered native turkey breed. Front. Vet. Sci. 2022, 9, 948502. [Google Scholar] [CrossRef]
- Imholt, D. Morphometrische Studien an Eiern von Hybrid-und Rassehühnern mit Versuchen zur Detektion einer Beziehung zwischen der Form von Eiern und dem Geschlecht der darin befindlichen Küken: Eine oologische und mathematische Studie. VVB Laufersweiler. 2010. Available online: http://geb.uni-giessen.de/geb/volltexte/2010/7760/ (accessed on 6 March 2023).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation Day | Class | No. of Eggs | No. of Images | |||
|---|---|---|---|---|---|---|
| Complete | Slightly Rotated | Partially Obscured | Total | |||
| Day 3 | Female | 1418 | 2246 | 457 | 34 | 2737 |
| Male | 1426 | 2259 | 473 | 30 | 2762 | |
| Day 4 | Female | 1418 | 2096 | 685 | 185 | 2966 |
| Male | 1426 | 2105 | 690 | 199 | 2994 | |
| Day 5 | Female | 1418 | 1628 | 692 | 312 | 2632 |
| Male | 1426 | 1634 | 707 | 329 | 2670 | |
| Total | 2844 | 11,968 | 3704 | 1089 | 16,761 | |
| Incubation Day | Class | Training Set | Testing Set | ||
|---|---|---|---|---|---|
| No. of Eggs | No. of Images | No. of Eggs | No. of Images | ||
| Day 3 | Female | 1000 | 1928 | 418 | 809 |
| Male | 1000 | 1944 | 426 | 818 | |
| Day 4 | Female | 1000 | 2124 | 418 | 842 |
| Male | 1000 | 2127 | 426 | 847 | |
| Day 5 | Female | 1000 | 1870 | 418 | 762 |
| Male | 1000 | 1884 | 426 | 786 | |
| Class | Length | Width | Shape Index | Weight | Eccentricity |
|---|---|---|---|---|---|
| Female | 55.2755 ± 2.3654 | 42.7008 ± 1.5772 | 1.2948 ± 0.0465 | 56.9026 ± 5.7508 | 0.6326 ± 0.0343 |
| Male | 54.9282 ± 2.1884 | 42.6404 ± 1.5810 | 1.2890 ± 0.0494 | 56.4981 ± 5.1345 | 0.6279 ± 0.0362 |
| Incubation Day | Class | Acc (%) | P (%) | R (%) | AP (%) | mAP (%) |
|---|---|---|---|---|---|---|
| Day 3 | Female | 87.68 | 88.60 | 89.05 | 86.05 | 85.81 |
| Male | 86.43 | 87.17 | 88.23 | 85.57 | ||
| Day 4 | Female | 90.32 | 93.21 | 83.39 | 89.67 | 88.79 |
| Male | 88.18 | 95.36 | 82.01 | 87.91 | ||
| Day 5 | Female | 85.23 | 83.60 | 81.39 | 84.73 | 82.29 |
| Male | 84.12 | 78.27 | 80.64 | 79.85 |
| Module | Exp No. 1 | Exp No. 2 | Exp No. 3 | Exp No. 4 |
|---|---|---|---|---|
| CBAM | × | √ | × | √ |
| BiFPN | × | × | √ | √ |
| mAP (%) | 84.82 | 87.11 | 86.65 | 88.28 |
| Algorithm | P (%) | R (%) | AP (%) | mAP (%) | Processing Time (ms) | Model Size (MB) |
|---|---|---|---|---|---|---|
| Faster R-CNN | 45.62 | 89.73 | 73.88 | 71.62 | 52.63 | 118.60 |
| SSD | 36.10 | 82.41 | 58.49 | 56.30 | 8.40 | 97.30 |
| YOLOv5 | 92.23 | 82.58 | 84.66 | 82.83 | 31.25 | 15.90 |
| Ours | 94.05 | 85.12 | 89.61 | 88.79 | 23.90 | 11.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, N.; Li, B.; Zhao, Y.; Fan, S.; Zhu, J.; Wang, H.; Zhao, W. Exploratory Study of Sex Identification for Chicken Embryos Based on Blood Vessel Images and Deep Learning. Agriculture 2023, 13, 1480. https://doi.org/10.3390/agriculture13081480
Jia N, Li B, Zhao Y, Fan S, Zhu J, Wang H, Zhao W. Exploratory Study of Sex Identification for Chicken Embryos Based on Blood Vessel Images and Deep Learning. Agriculture. 2023; 13(8):1480. https://doi.org/10.3390/agriculture13081480
Chicago/Turabian StyleJia, Nan, Bin Li, Yuliang Zhao, Shijie Fan, Jun Zhu, Haifeng Wang, and Wenwen Zhao. 2023. "Exploratory Study of Sex Identification for Chicken Embryos Based on Blood Vessel Images and Deep Learning" Agriculture 13, no. 8: 1480. https://doi.org/10.3390/agriculture13081480