
Characterisation of Pectinolytic Bacillus pumilus and Paenibacillus amyloliticus Strains, New Pathogens of Potato in Tunisia
 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Potato Sampling and Isolation of Bacterial Strains and Their Growth Conditions
2.2. Isolation of Pectinolytic Bacteria
2.3. MALDI-TOF MS Analysis
2.4. Sequencing of 16S rDNA
2.5. Phenotypic Characterisation of the Isolated Strains
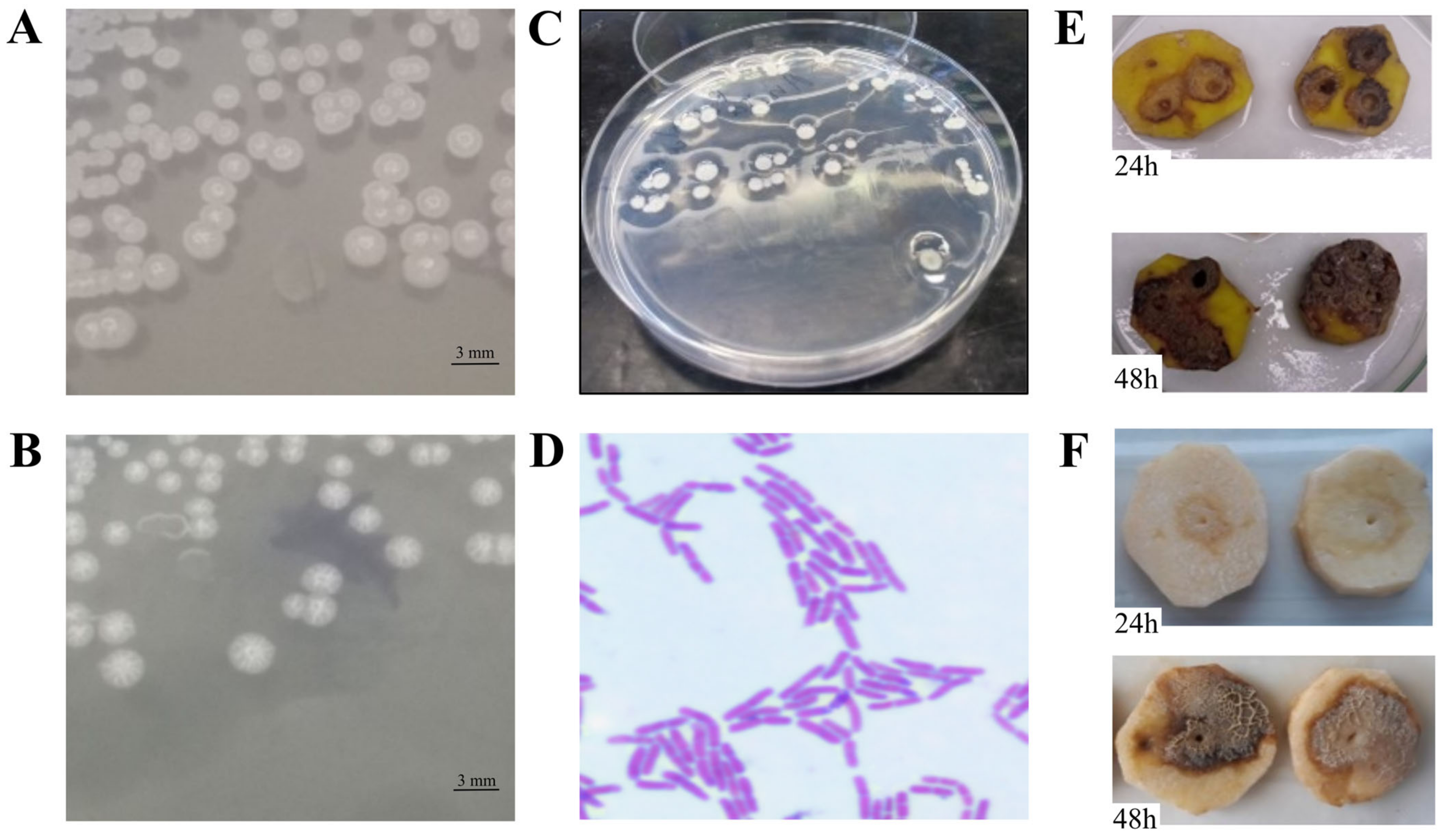
2.5.1. Bacterial Ability to Macerate Potato Tuber Tissue
2.5.2. Production of Extracellular Enzymes and Bacterial Motility
2.5.3. Biochemical Profiling
2.6. Statistical Analysis
3. Results
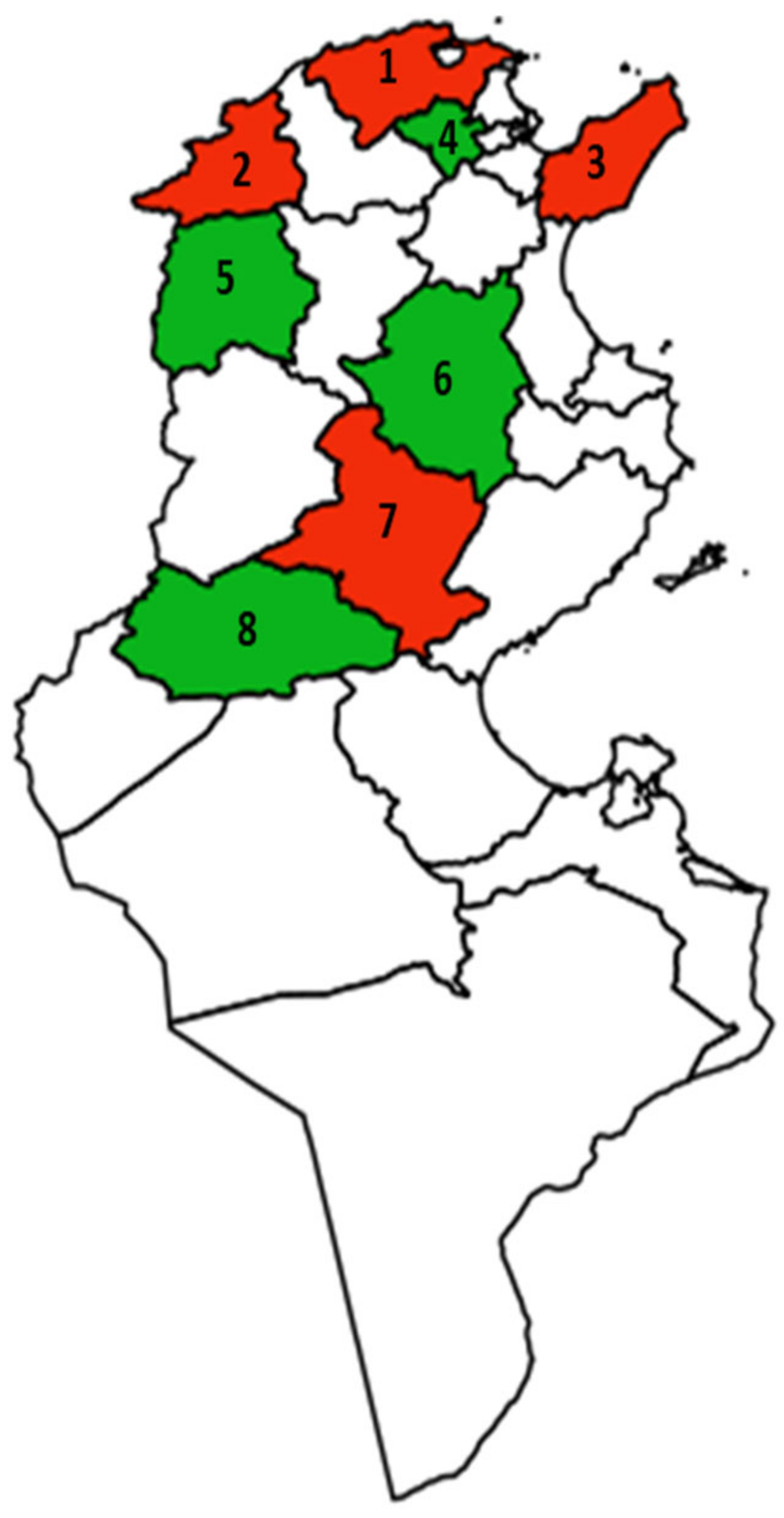
3.1. Occurrence of Pectinolytic Bacteria in Potato Fields in the Territory of Tunisia
3.2. Identification of the Collected Pectinolytic Isolates
3.3. Phenotypic Characterisation of the Collected Pectinolytic Isolates
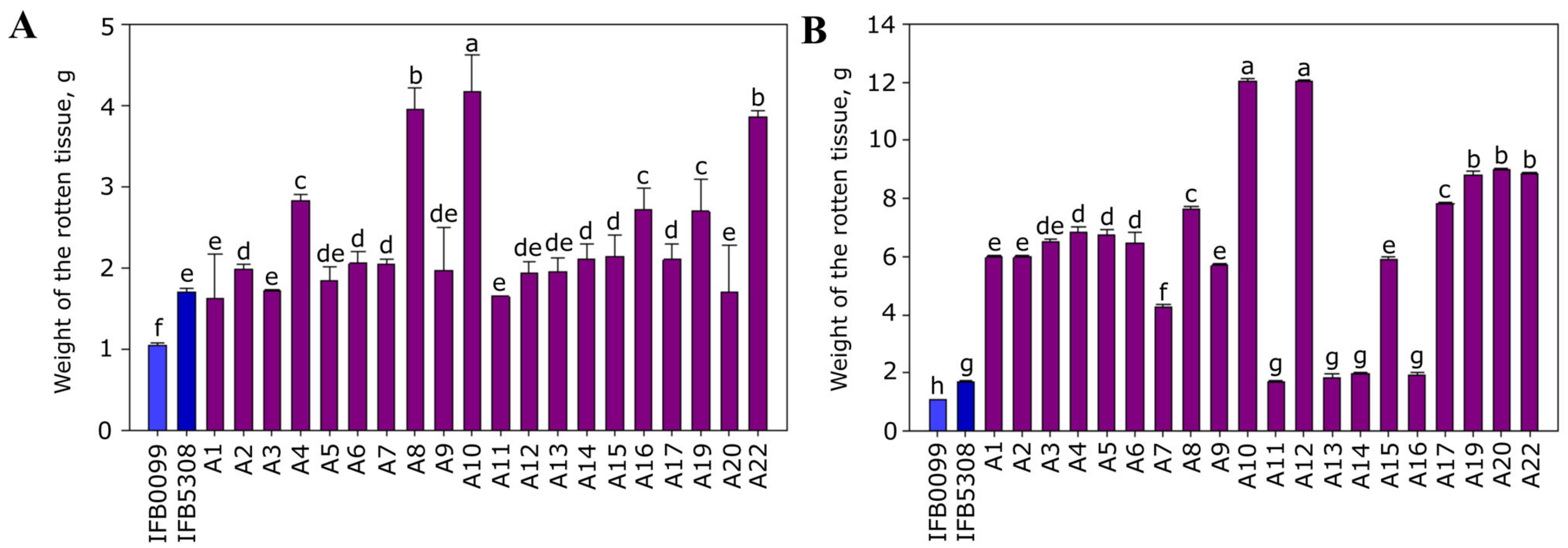
3.3.1. Bacterial Ability to Macerate Potato Tuber Tissue
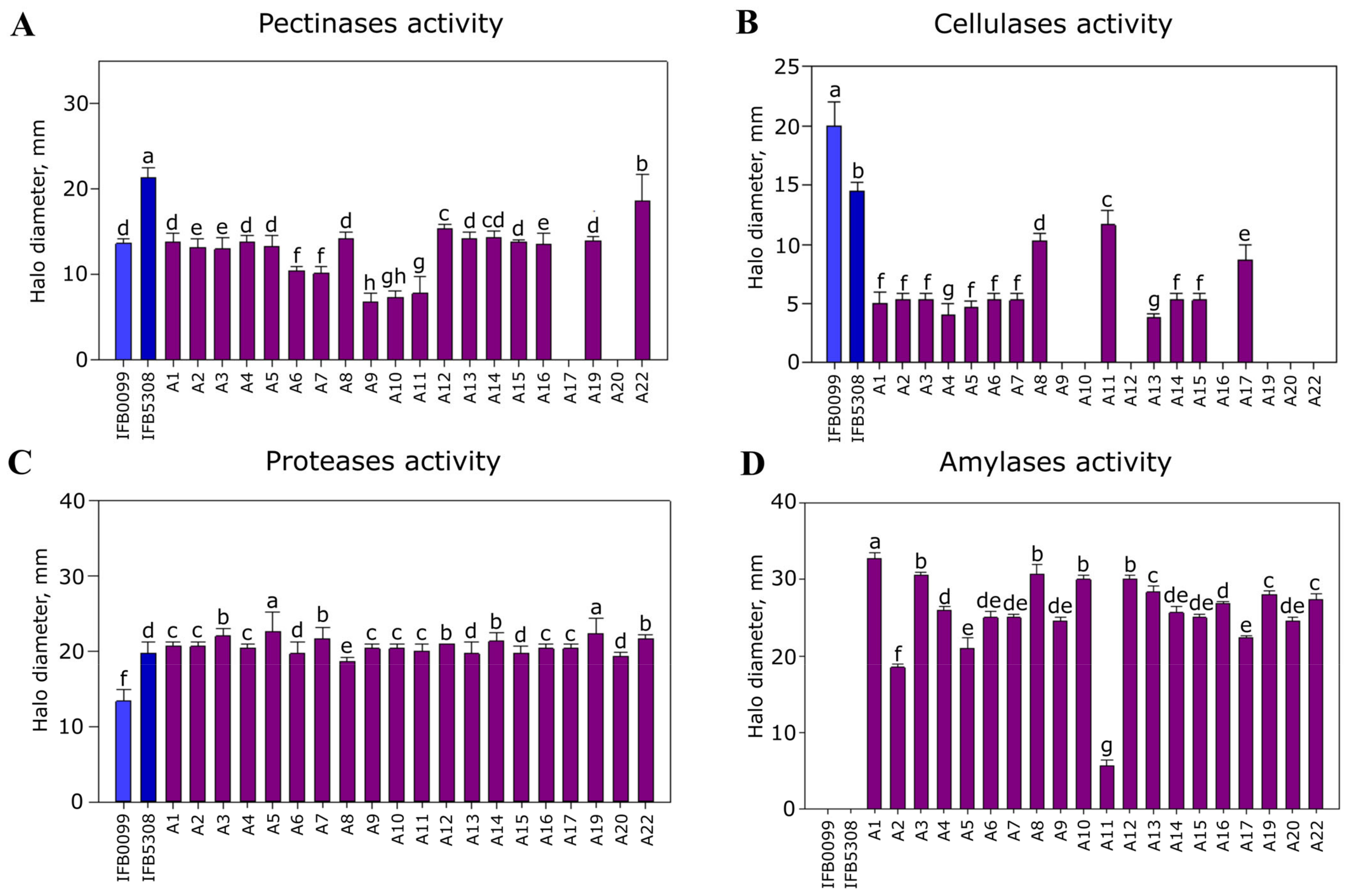
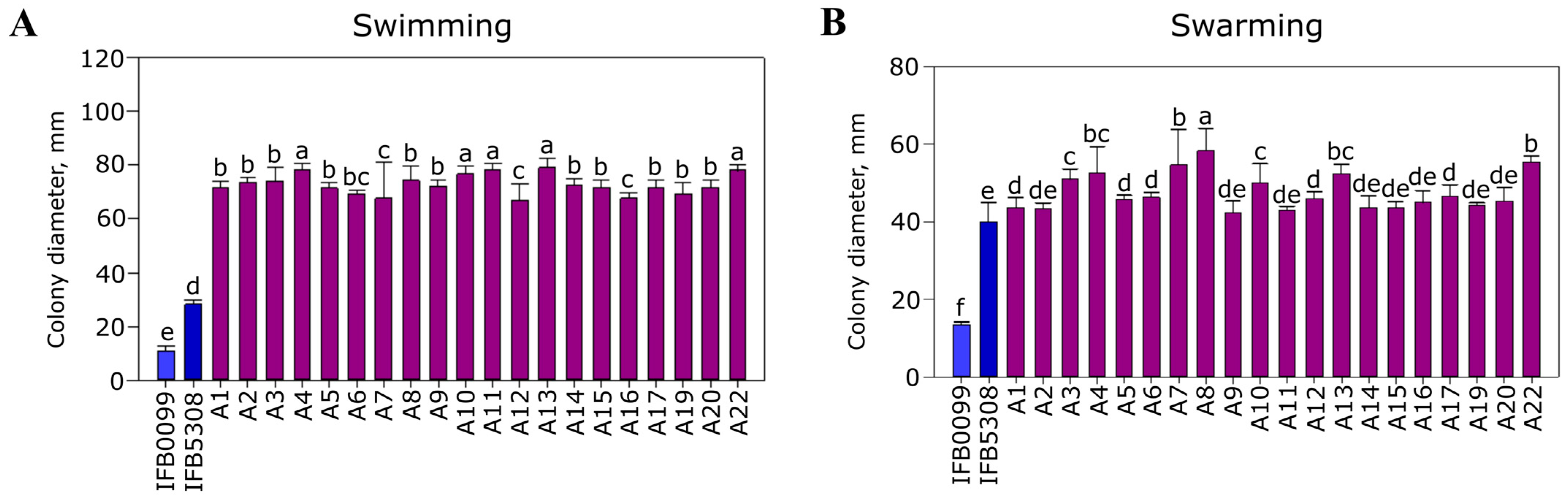
3.3.2. Major Pathogenicity-Associated Features
3.3.3. Biochemical Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chemak, F.; Allagui, L.; Ali, Y. Analyse des performances techniques des producteurs de la pomme de terre en Tunisie. Une approche non paramétrique. New Medit. 2014, 13, 72–80. [Google Scholar]
- Perombelon, M.C.M.; Kelman, A. Ecology of the soft rot Erwinias. Ann. Rev. Phytopathol. 1980, 18, 361–387. [Google Scholar] [CrossRef]
- Toth, I.K.; Bell, K.S.; Holeva, M.C.; Birch, P.R.J. Soft rot erwiniae: From genes to genomes. Mol. Plant Pathol. 2003, 4, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.K.; van der Wolf, J.M.; Saddler, G.; Lojkowska, E.; Helias, V.; Pirhonen, M.; Tsror, L.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe. Plant. Pathol 2011, 60, 385–399. [Google Scholar] [CrossRef]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Hélias, V.; Hamon, P.; Huchet, E.; van der Wolf, J.M.; Andrivon, D. Two new effective semiselective crystal violet pectate media for isolation of Pectobacterium and Dickeya. Plant Pathol. 2012, 61, 339–345. [Google Scholar] [CrossRef]
- De Boer, S.H.; Li, X.; Ward, L.J. Pectobacterium spp. associated with bacterial stem rot syndrome of potato in Canada. Phytopathology 2012, 102, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Galal, A.A.; El-Bana, A.A.; Janse, J. Bacillus pumilus, a new pathogen on mango plants. Egypt. J. Phytopathol. 2006, 34, 17–29. [Google Scholar]
- Bathily, H.; Babana, A.H.; Samaké, F. Bacillus pumilus, a new pathogen on potato tubers in storage in Mali. Afr. J. Microbiol. Res. 2010, 4, 2067–2071. [Google Scholar]
- Peng, Q.; Yuan, Y.; Gao, M. Bacillus pumilus, a novel ginger rhizome rot pathogen in China. Plant Dis. 2013, 97, 1308–1315. [Google Scholar] [CrossRef] [Green Version]
- Kovaleva, V.A.; Shalovylo, Y.I.; Gorovik, Y.N.; Lagonenko, A.L.; Evtushenkov, A.N.; Gout, R.T. Bacillus pumilus—A new phytopathogen of Scots pine. J. Forest Sci. 2015, 61, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Saleh, O.I.; Huang, P.-Y.; Huang, J.-S. Bacillus pumilus, the cause of bacterial blotch of immature balady peach in Egypt. J. Phytopathol. 1997, 145, 447–453. [Google Scholar] [CrossRef]
- Song, J.H.; Wu, Z.R.; Zhang, L.X.; Tan, G.J.; Wang, S.; Wang, J.J. First report of Bacillus pumilus causing fruit rot on muskmelon (Cucumis melo) in China. Plant Dis. 2018, 102, 439. [Google Scholar] [CrossRef]
- Hakim, M.; Liaquat, F.; Gul, S.; Chaudhary, H.J.; Munis, M.F.H. Presence of Bacillus pumilus causing fruit rot of Ficus lacor in Pakistan. J. Plant Pathol. 2015, 97, 543. [Google Scholar]
- Ahmadi, E.; Kowsari, M.; Azadfar, D.; Jouzani, G.S. Bacillus pumilus and Stenotrophomonas maltophilia as two potentially causative agents involved in Persian oak decline in Zagros forests (Iran). Forest Pathol. 2019, 49, e12541. [Google Scholar] [CrossRef]
- Mazlan, S.; Zulperi, D.; Wahab, A.; Jaafar, N.M.; Sulaiman, Z.; Rajandas, H. First report of Bacillus pumilus causing trunk bulges of rubber tree (Hevea brasiliensis) in Malaysia. Plant Dis. 2019, 103, 1016. [Google Scholar] [CrossRef]
- Font, M.I.; Bassimba, D.D.M.; Cebrián, M.C.; Molina, L.M.; Jordá, C. First report of Bacillus pumilus on Phaseolus vulgaris in Spain. Plant Pathol. 2010, 59, 2. [Google Scholar] [CrossRef]
- Evdokimova, O.V.; Miamin, V.E.; Valentovich, L.N. Biochemical and molecular genetic characteristics of Bacillus pumilus bacteria, isolated on the territory of Belarus. J. Belarusian State Univ. Biol. 2018, 1, 39–47. [Google Scholar]
- Priou, S.; El-Mahjoub, M. Bacterial and fungal diseases in the major potato-growing areas of Tunisia. EPPO Bull. 1999, 29, 167–171. [Google Scholar] [CrossRef]
- Yahiaoui-Zaidi, R.; Ladjouzi, R.; Benallaoua, S. Pathogenic variability within biochemical groups of Pectobacterium carotovorum isolated in Algeria from seed potato tubers. Int. J. Biotech. Mol. Biol. Res. 2010, 1, 1–9. [Google Scholar]
- El Arbia, A.; Rochexa, A.; Chataignéa, G.; Bécheta, M.; Lecouturiera, D.; Arnaulda, S.; Gharsallah, N.; Jacques, P. The Tunisian oasis ecosystem is a source of antagonistic Bacillus spp. producing diverse antifungal lipopeptides. Res. Microbiol. 2015, 167, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, M.; Golanowska, M.; Hugouvieux-Cotte-Pattat, N.; Lojkowska, E. Regulators involved in Dickeya solani virulence, genetic conservation, and functional variability. Mol. Plant Microbe Interact. 2014, 27, 700–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoledowska, S.; Motyka-Pomagruk, A.; Sledz, W.; Mengoni, A.; Lojkowska, E. High genomic variability in the plant pathogenic bacterium Pectobacterium parmentieri deciphered from de novo assembled complete genomes. BMC Genom. 2018, 19, 751. [Google Scholar] [CrossRef] [Green Version]
- Golanowska, M.; Kielar, J.; Lojkowska, E. The effect of temperature on the phenotypic features and the maceration ability of Dickeya solani strains isolated in Finland, Israel and Poland. Eur. J. Plant Pathol. 2017, 147, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Zoledowska, S.; Motyka, A.; Zukowska, D.; Sledz, W.; Lojkowska, E. Population structure and biodiversity of Pectobacterium parmentieri isolated from potato fields in temperate climate. Plant Dis. 2018, 102, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Reverchon, S.; Van Gijsegem, F.; Rouve, M.; Kotoujansky, A.; Robert-Baudouy, J. Organization of a pectate lyase gene family in Erwinia chrysanthemi. Gene 1986, 49, 215–224. [Google Scholar] [CrossRef]
- Moran, F.; Nasuno, S.; Starr, M.P. Extracellular and Intracellular Polygalacturonic Acid h-arts-Eliminases of Erwinia carotovora. Arch. Biochem. Biophys. 1968, 123, 298–306. [Google Scholar] [CrossRef]
- Ji, J.; Hugouvieux-Cotte-Pattat, N.; Robert-Baudouy, J. Use of Mu-lac insertions to study the secretion of pectate lyases by Erwinia chrysanthemi. Microbiology 1987, 133, 793–802. [Google Scholar] [CrossRef] [Green Version]
- Carrim, A.J.I.; Barbosa, E.C.; Vieira, J.D.G. Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrents cham.(Carobinha-do-campo). Braz. Archiv. Biol. Technol. 2006, 49, 353–359. [Google Scholar] [CrossRef]
- Harshey, R.M. Bacterial motility on a surface: Many ways to a common goal. Ann. Rev. Microbiol. 2003, 57, 249–273. [Google Scholar] [CrossRef]
- Borisov, V.B.; Forte, E.; Davletshin, A.; Mastronicola, D.; Sarti, P.; Giuffrè, A. Cytochrome bd oxidase from Escherichia coli displays high catalase activity: An additional defence against oxidative stres. FEBS Lett. 2013, 587, 2214–2218. [Google Scholar] [CrossRef]
- Romdhani, M.S.; El-Mahjoub, M. Latent infection of potato tubers with Erwinia spp. in Tunisia. Meded. Van Fac. Landbouwwet. Rijksuniv. Gent 1990, 55, 1119–1123. [Google Scholar]
- Motyka-Pomagruk, A.; Zoledowska, S.; Sledz, W.; Lojkowska, E. The occurrence of bacteria from different species of Pectobacteriaceae on seed potato plantations in Poland. Eur. J. Plant Pathol. 2021, 159, 309–325. [Google Scholar] [CrossRef]
- Potrykus, M.; Golanowska, M.; Sledz, W.; Zoledowska, S.; Motyka, A.; Kolodziejska, A.; Butrymowicz, J.; Lojkowska, E. Biodiversity of Dickeya spp. isolated from potato plants and water sources in temperate climate. Plant Dis. 2016, 100, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Leifert, C.; Li, H.; Chidburee, S.; Hampson, S.; Workman, S.; Sigee, D.; Epton, H.A.; Harbour, A. Antibiotic production and biocontrol activity by Bacillus subtilis CL27 and Bacillus pumilus CL45. J. Appl. Bacteriol. 1995, 78, 97–108. [Google Scholar] [CrossRef]
- Gutiérrez-Mañero, F.J.; Ramos-Solano, F.; Probanza, A.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiologia Plantarum. 2001, 111, 206–211. [Google Scholar] [CrossRef]
- Kimouli, M.; Vrioni, M.; Papadopoulou, M.; Koumaki, V.; Petropoulou, D.; Gounaris, A.; Friedrich, A.W.; Tsakris, A. Two cases of severe sepsis caused by Bacillus pumilus in neonatal infants. J. Med. Microbiol. 2012, 61, 596–599. [Google Scholar] [CrossRef] [Green Version]
- Tena, D.; Martínez-Torres, J.A.; Pérez-Pomata, M.T.; Sáez-Nieto, J.A.; Rubio, V.; Bisquert, J. Cutaneous infection due to Bacillus pumilus: Report of 3 cases. Clin. Infect. Dis. 2007, 44, e40–e42. [Google Scholar] [CrossRef]
- From, C.; Hormazabal, V.; Granum, P.E. Food poisoning associated with pumilacidin-producing Bacillus pumilus in rice. Inter. J. Food Microbiol. 2007, 115, 319–324. [Google Scholar] [CrossRef]
- Vockler, C.J.; Greenfield, P.; Tran-Dinh, N.; Midgley, D.J. Draft genome sequence of Bacillus pumilus Fairview, an isolate recovered from a microbial methanogenic enrichment of coal seam gas formation water from Queensland, Australia. Genome Announc. 2014, 2, e00279-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, L.; Sawyer, J.; Venkateswaran, K.; Nicholson, W. Extreme spore UV resistance of Bacillus pumilus isolates obtained from an ultraclean spacecraft assembly facility. Microb. Ecol. 2004, 47, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Fact. 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Wolf, J.M.; Acuña, I.; de Boer, S.H.; Brurberg, M.B.; Cahill, G.; Charkowski, A.O.; Coutinho, T.; Davey, T.; Dees, M.W.; Degefu, Y.; et al. Diseases caused by Pectobacterium and Dickeya species around the world. In Plant Diseases Caused by Dickeya and Pectobacterium Species; van Gijsegem, F., van der Wolf, J.M., Toth, I.K., Eds.; Springer Nature: Cham, Switzerland, 2021; pp. 215–262. [Google Scholar]
- Slovareva, O.Y. Detection and identification of wheat and barley phytopathogens in Russia. Microbiol. Ind. Res. J. 2020, 7, 13–23. [Google Scholar]
- Zenelt, W.; Krawczyk, K.; Borodynko-Filas, N. Biodiversity and scope of endophytic and phytopathogenic bacterial species identified in plant samples investigated in the Plant Disease Clinic laboratory. J. Plant Prot. Res. 2021, 61, 63–82. [Google Scholar]
- Motyka, A.; Zoledowska, S.; Sledz, W.; Lojkowska, E. Molecular methods as tools to control plant diseases caused by Dickeya and Pectobacterium spp: A minireview. New Biotech. 2017, 39, 181–189. [Google Scholar] [CrossRef]
- Huang, Q.; Yong, P.; Xin, L.; Haifeng, W.; Yizheng, Z. Purification and characterization of an extracellular alkaline serine protease with dehairing function from Bacillus pumilus. Curr. Microbiol. 2003, 46, 169–173. [Google Scholar] [CrossRef]
- Basu, S.; Manabendra, N.S.; Dhrubajyoti, C.; Krishanu, C. Large-scale degumming of ramie fibre using a newly isolated Bacillus pumilus DKS1 with high pectate lyase activity. J. Ind. Microbiol. Biotech. 2009, 36, 239–245. [Google Scholar] [CrossRef]
- Yan, H.; Hui, W.; Bo, Y.; ShengMou, H.; YunJie, L.; JinJu, W. Isolation and identification of Bacillus pumilus HB-3 and purification characteristics of amylase. Storage Proc. 2019, 2, 62–68. [Google Scholar]
- Ariffin, H.; Abdullah, H.; Umi Kalsom, M.S.; Shirai, Y.; Hassan, M.A. Production and characterization of cellulase by Bacillus pumilus EB3. Int. J. Eng. Technol. 2006, 3, 47–53. [Google Scholar]
- Kayath, C.A.; Zamba, A.I.; Mokembiabeka, S.N.; Opa-Iloy, M.; Wilson, P.S.E.; Kaya-Ongoto, M.D.; Mouellet Maboulou, R.J.; Nguimbi, E. Synergic involvements of microorganisms in the biomedical increase of polyphenols and flavonoids during the fermentation of ginger juice. Int. J. Microbiol. 2020, 2020, 8417693. [Google Scholar] [CrossRef]
- McSpadden Gardener, B.B. Ecology of Bacillus and Paenibacillus spp. in agricultural systems. Phytopathology 2004, 94, 1252–1258. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code Strain | Governorate/ Region/Locality | GPS Location | Year/Month | Irrigation Type | Fertilisation | Cultivar | Organ/Symptom |
|---|---|---|---|---|---|---|---|
| A1 | Bizerte/ Ras Djebel/Kichi | 37.216273, 10.109030 | 2019/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A2 | Bizerte/ Ras Djebel/Kichi | 37.216273, 10.109030 | 2019/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A3 | Nabeul/ Beni Khiar/Melliti | 36.478755, 10.770850 | 2019/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A4 | Bizerte/Ras Djebel/ Kichi | 37.216273, 10.109030 | 2019/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A5 | Bizerte/ Ghar Milh/Mohamed | 37.169613, 10.147375 | 2019/June | Pond water | Chemical | Spunta | Tuber/soft rot |
| A6 | Bizerte/ Ras Djebel/Kichi | 37.216273, 10.109030 | 2019/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A7 | Jandouba/ Bousalem/Bousalem1 | 36.612511, 9.000860 | 2019/February | Drip irrigation | Biological and chemical | Daifla | Tuber/soft rot |
| A8 | Bizerte/ Ras Djebal/Kichi | 37.216273, 10.109030 | 2020/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A9 | Bizerte/ Ras Djebal/Kichi | 37.216273, 10.109030 | 2020/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A10 | Sidi Bouzid/ Bir El Hafey/Anonym | 34.930545, 9.226859 | 2019/June | Drip irrigation | Biological | Spunta | Tuber/soft rot |
| A11 | Nabeul/ Beni Khiar/Melliti | 36.478755, 10.770850 | 2019/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A12 | Jandouba/ Bousalem/Bousalem1 | 36.612511, 9.000860 | 2019/February | Drip irrigation | Biological and chemical | Daifla | Tuber/soft rot |
| A13 | Bizerte/ Ras Djebel/Kichi | 37.216273, 10.109030 | 2020/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A14 | Bizerte/ Ras Djebel/Kichi | 37.216273, 10.109030 | 2020/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A15 | Jandouba/ Bousalem/Bousalem 2 | 36.623775, 8.958674 | 2020/February | Drip irrigation | Biological and chemical | Daifla | Tuber/soft rot |
| A16 | Jandouba/ Bousalem/Bousalem2 | 36.623775, 8.958674 | 2020/February | Drip irrigation | Biological and chemical | Daifla | Tuber/soft rot |
| A17 | Jandouba/ Bousalem/Bousalem2 | 36.623775, 8.958674 | 2020/February | Drip irrigation | Biological and chemical | Daifla | Tuber/soft rot |
| A19 | Bizerte/ Ras Djebel/Kichi | 37.216273, 10.109030 | 2020/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| A20 | Bizerte/ Ghar Milh/Mohamed | 37.169613, 10.147375 | 2020/June | Pond water | Chemical | Spunta | Tuber/soft rot |
| A22 | Jandouba/ Bousalem/Bousalem2 | 36.623775, 8.958674 | 2020/June | Drip irrigation | Biological and chemical | Spunta | Tuber/soft rot |
| Number | Isolate | MALDI-TOF MS-Based Classification | Log (Score) MALDI-TOF MS | Accession Number for the Sequenced 16S rDNA Gene Fragments | Final Identification |
|---|---|---|---|---|---|
| 1 | A1 | Bacillus pumilus | 2.02 | ON898611 | Bacillus pumilus |
| 2 | A2 | Bacillus pumilus | 2.03 | ON899811 | Bacillus pumilus |
| 3 | A3 | Bacillus pumilus | 1.82 | ON899941 | Bacillus pumilus |
| 4 | A4 | Bacillus pumilus | 1.99 | ON899942 | Bacillus pumilus |
| 5 | A5 | Bacillus pumilus | 1.96 | ON900081 | Bacillus pumilus |
| 6 | A6 | Bacillus pumilus | 2.03 | ON900082 | Bacillus pumilus |
| 7 | A7 | Bacillus pumilus | 1.86 | ON900083 | Bacillus pumilus |
| 8 | A8 | Bacillus pumilus | 1.82 | ON900084 | Bacillus pumilus |
| 9 | A9 | Bacillus pumilus | 2.04 | ON900085 | Bacillus pumilus |
| 10 | A10 | Bacillus pumilus | 1.90 | ON900086 | Bacillus pumilus |
| 11 | A11 | Bacillus altitudinis | 1.98 | ON922531 | Bacillus pumilus |
| 12 | A12 | Paenibacillus amyloliticus | 2.18 | ON922535 | Paenibacillus amyloliticus |
| 13 | A13 | Bacillus pumilus | 2.03 | ON922532 | Bacillus pumilus |
| 14 | A14 | Bacillus pumilus | 2.06 | ON922533 | Bacillus pumilus |
| 15 | A15 | Bacillus pumilus | 1.88 | ON922534 | Bacillus pumilus |
| 16 | A16 | Bacillus pumilus | 1.72 | ON921248 | Bacillus pumilus |
| 17 | A17 | Bacillus pumilus | 1.85 | ON921249 | Bacillus pumilus |
| 18 | A19 | Bacillus pumilus | 2.00 | ON921250 | Bacillus pumilus |
| 19 | A20 | Bacillus pumilus | 1.91 | ON921251 | Bacillus pumilus |
| 20 | A22 | Bacillus pumilus | 2.05 | ON921252 | Bacillus pumilus |
| Isolate | ONPG | ADH | LDC | ODC | CIT | H2S | URE | TDA | IND | VP | GEL | GLU | MAN | IN | SOR | RHA | SAC | MEL | AMY | ARA | OX |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A1 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A2 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A3 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A4 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A5 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A6 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A7 | + | + | − | + | + | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | − |
| A8 | + | + | − | + | + | − | − | − | − | + | + | + | + | + | + | + | + | + | + | − | − |
| A9 | + | + | − | + | + | − | − | − | − | + | + | + | + | + | + | + | + | + | + | − | − |
| A10 | + | − | − | + | + | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | − |
| A11 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | + | + | − | + | − | − |
| A12 | + | + | − | + | + | − | − | − | − | + | + | − | − | − | − | + | + | + | + | + | − |
| A13 | + | + | − | + | + | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | − |
| A14 | + | + | − | + | + | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | − |
| A15 | + | + | − | + | + | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | − |
| A16 | + | + | − | + | + | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | − |
| A17 | + | + | − | + | + | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | − |
| A19 | + | − | − | − | − | − | − | − | − | + | + | + | + | − | − | − | + | − | + | − | − |
| A20 | + | + | − | + | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | + | − |
| A22 | + | + | − | + | + | − | − | − | − | − | + | + | + | + | + | + | + | + | + | + | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahyaoui, A.; Oueslati, M.; Motyka-Pomagruk, A.; Kaczynska, N.; Sledz, W.; Tarhouni, B.; Hajlaoui, M.R.; Lojkowska, E.; Sadfi-Zouaoui, N. Characterisation of Pectinolytic Bacillus pumilus and Paenibacillus amyloliticus Strains, New Pathogens of Potato in Tunisia. Agriculture 2023, 13, 1275. https://doi.org/10.3390/agriculture13061275
Yahyaoui A, Oueslati M, Motyka-Pomagruk A, Kaczynska N, Sledz W, Tarhouni B, Hajlaoui MR, Lojkowska E, Sadfi-Zouaoui N. Characterisation of Pectinolytic Bacillus pumilus and Paenibacillus amyloliticus Strains, New Pathogens of Potato in Tunisia. Agriculture. 2023; 13(6):1275. https://doi.org/10.3390/agriculture13061275
Chicago/Turabian StyleYahyaoui, Anissa, Maroua Oueslati, Agata Motyka-Pomagruk, Natalia Kaczynska, Wojciech Sledz, Belhassen Tarhouni, Mohamed Rabeh Hajlaoui, Ewa Lojkowska, and Najla Sadfi-Zouaoui. 2023. "Characterisation of Pectinolytic Bacillus pumilus and Paenibacillus amyloliticus Strains, New Pathogens of Potato in Tunisia" Agriculture 13, no. 6: 1275. https://doi.org/10.3390/agriculture13061275