
Effects of Variations in Soil Moisture and Phosphorus Concentrations on the Diversity of the Arbuscular Mycorrhizal Fungi Community in an Agricultural Ecosystem
Abstract
:1. Introduction
2. Methods
2.1. Study Site
2.2. Sample Collection and Experiment Method
2.3. DNA Extraction, Gene Amplification, and Sequencing
2.4. Data Analysis
3. Results
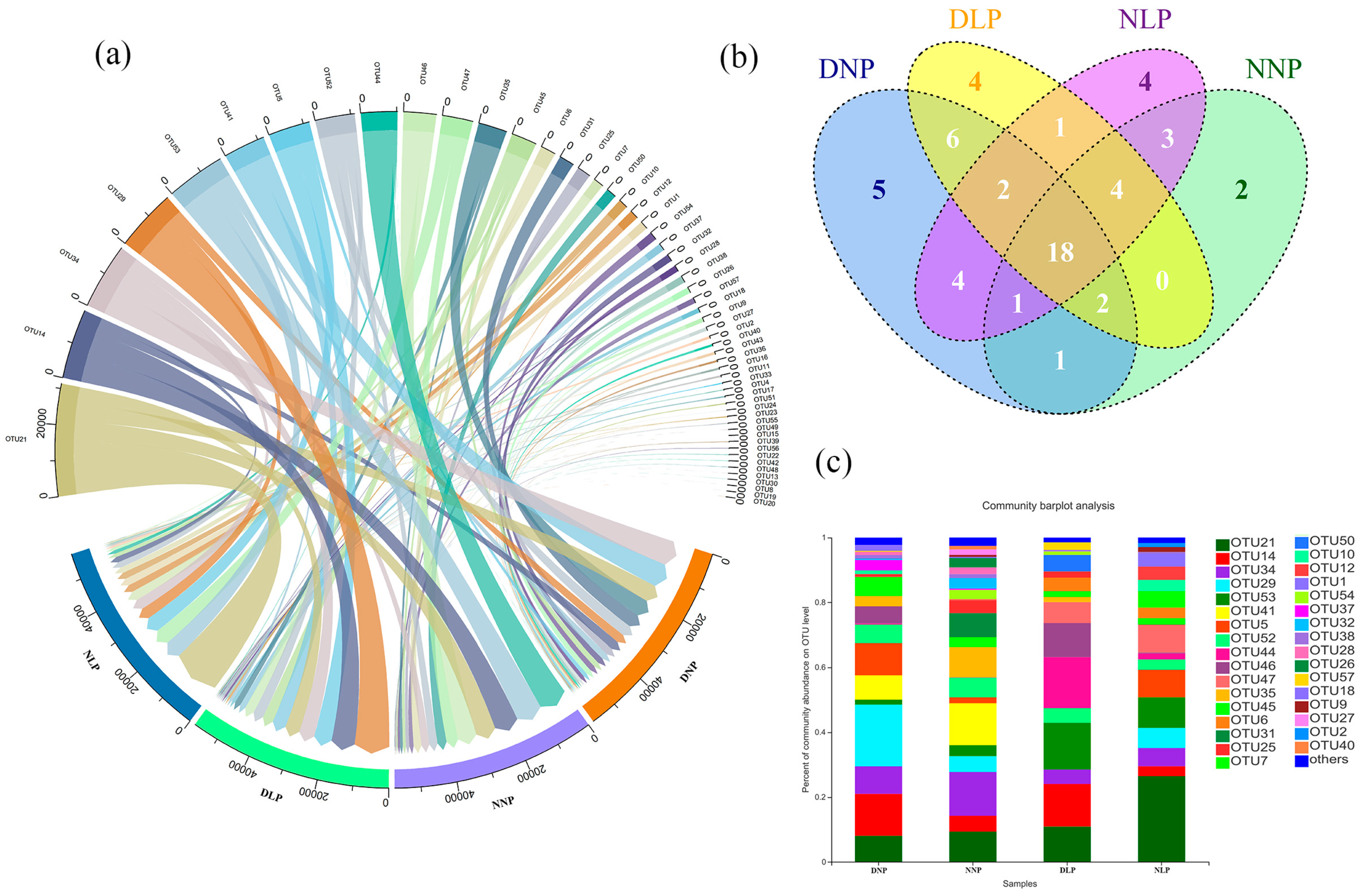
3.1. Distribution Patterns of AMF in Different Habitats
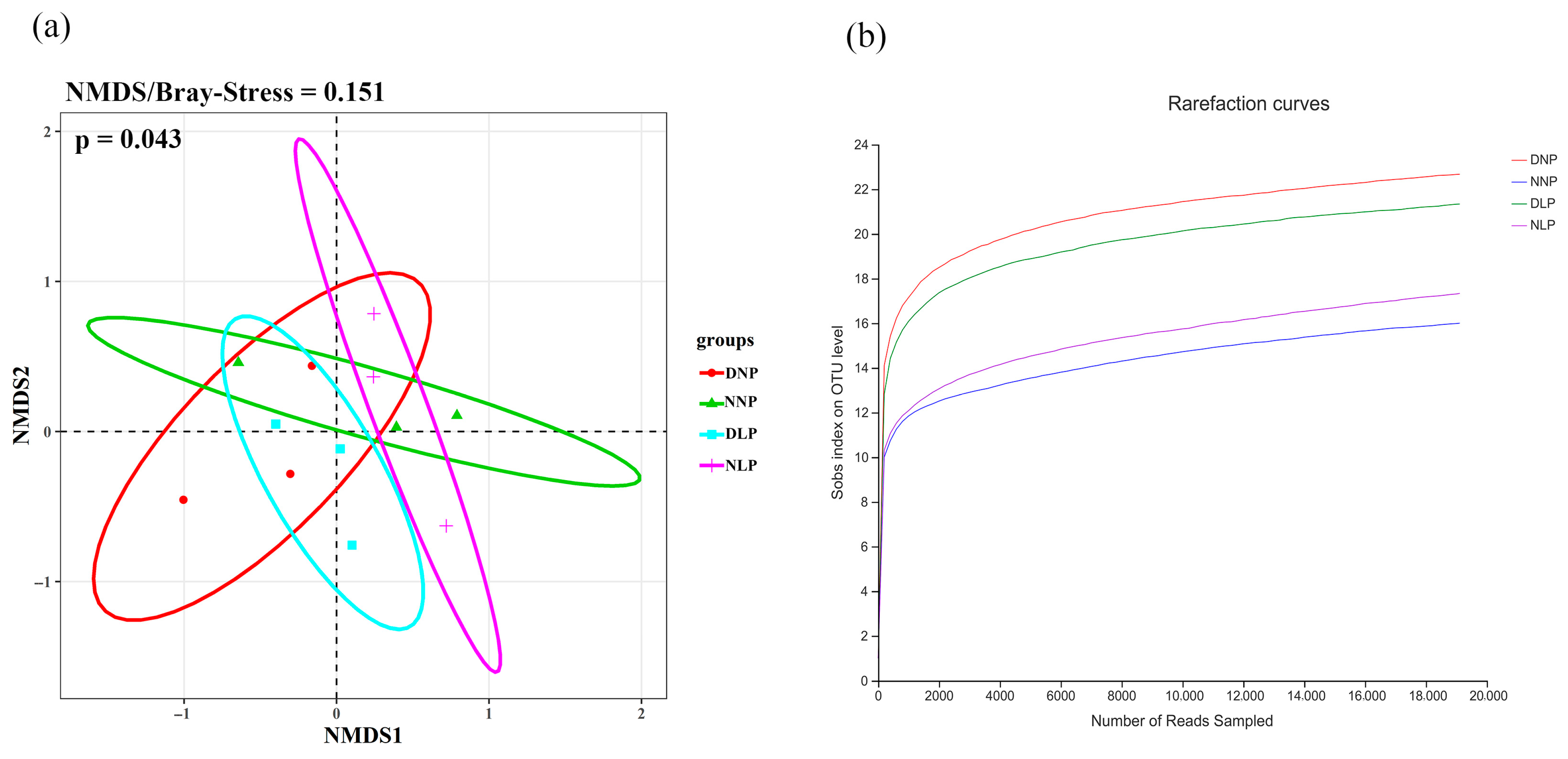
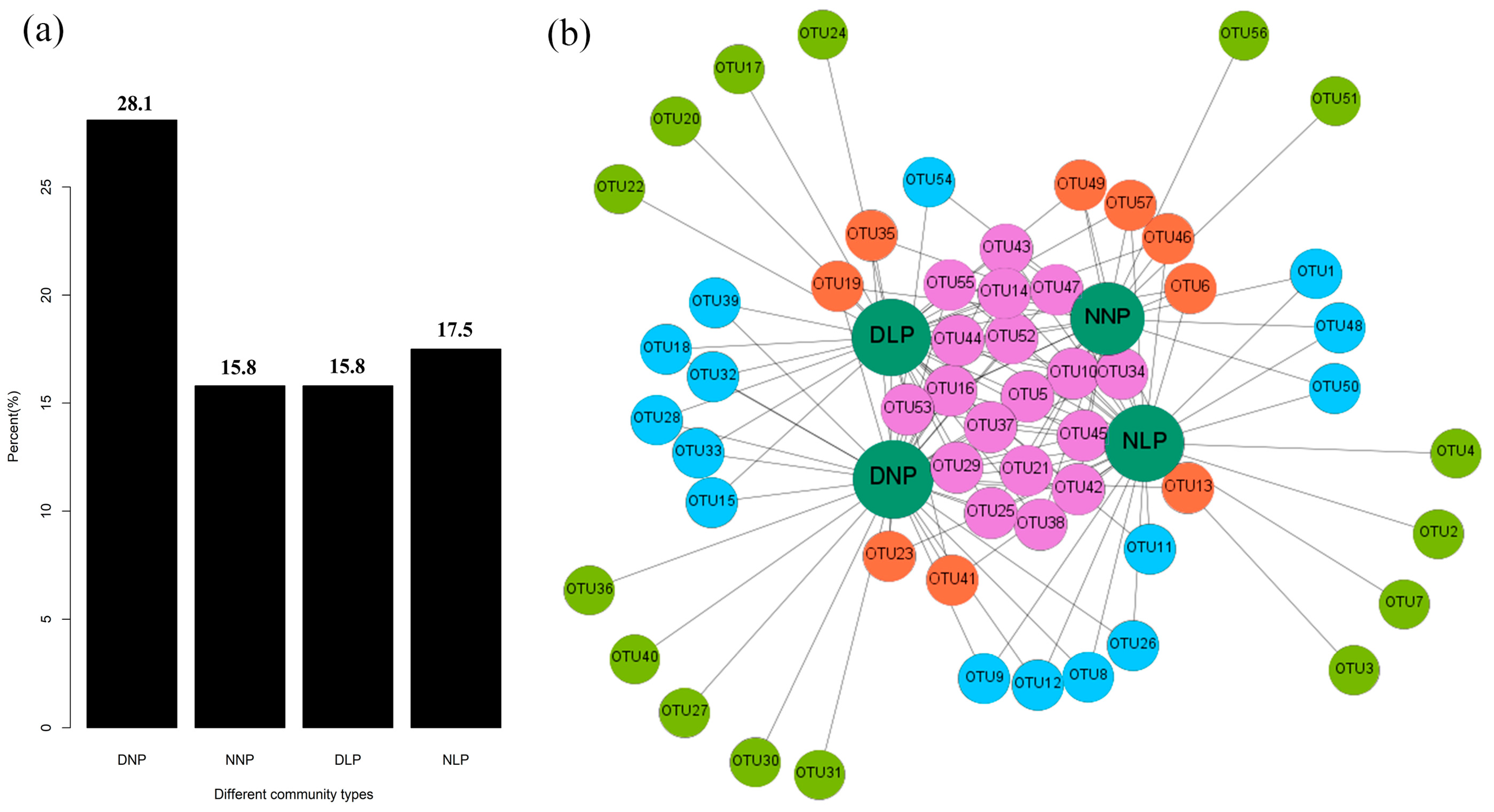
3.2. Differences of AMF in Different Habitats
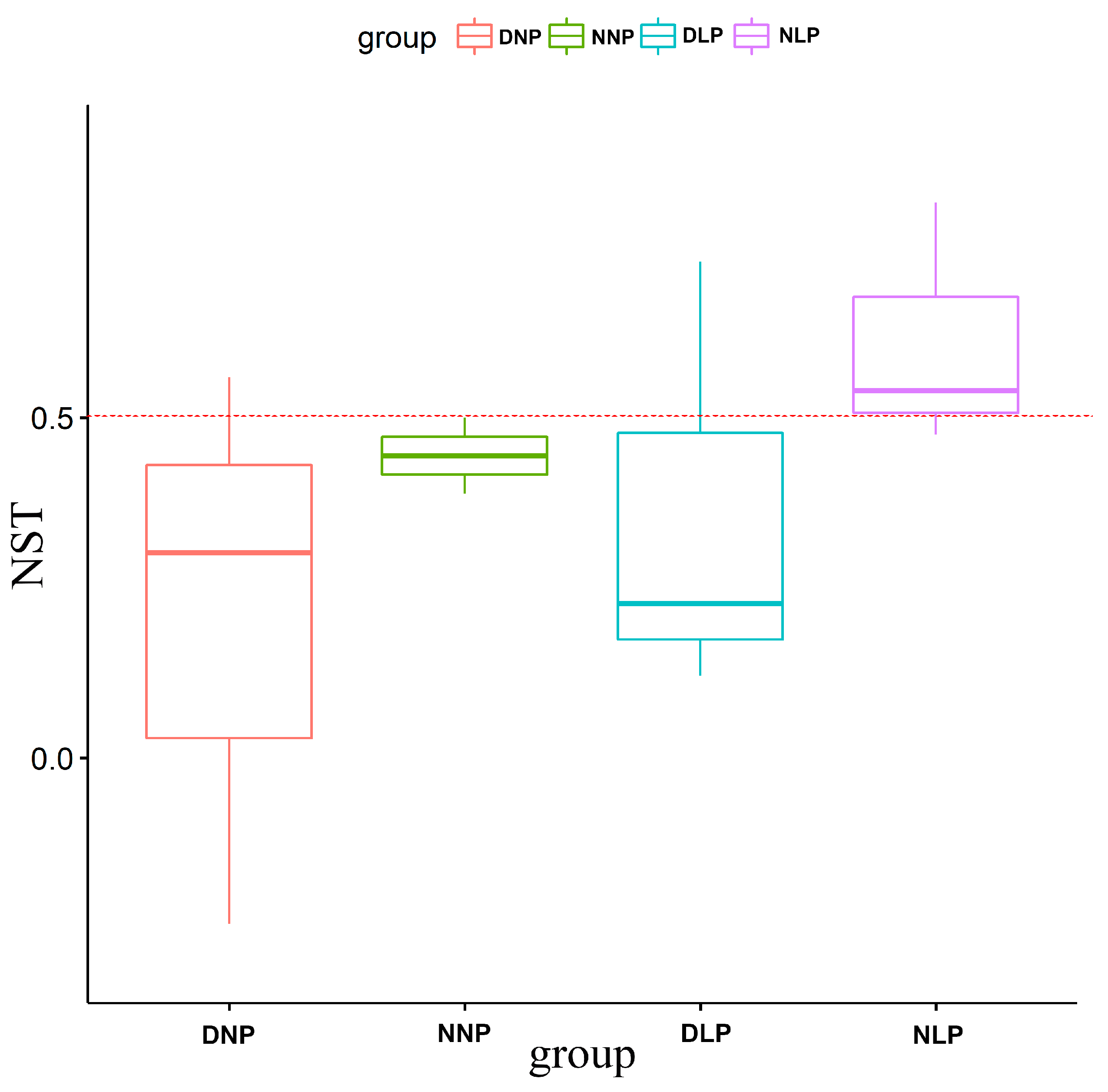
3.3. Relevance of AMF and Different Habitats
4. Discussion
4.1. Relationship between AMF and Different Moisture Contents in Farmland Soil
4.2. Effects of Phosphorus Application on AMF Diversity in Farmland Soil
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, K.; Zai, S.; Wu, F.; Bai, M. Effects of returning wheat or corn straw on soil water infiltration. J. Northeast Agric. Univ. 2022, 53, 35–42, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Tian, X.; Ma, S.; Wang, M. Arbuscular mycorrhizal fungi on plant resistant to the effects of water stress research progress. Rural Sci. Technol. 2021, 12, 80–83. [Google Scholar]
- Richardson, A.E. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Funct. Plant Biol. 2001, 28, 897–906. [Google Scholar] [CrossRef]
- Malhotra, H.; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. Plant Nutr. Abiotic Stress Toler. 2018, 171–190. [Google Scholar]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility: A review. Agron. Sustain. Dev. 2010, 30, 401–422. [Google Scholar] [CrossRef] [Green Version]
- Sarula; Yang, H.; Tai, J.; Gao, J.; Li, Y. Effect of straw maturing agents on fungal diversity in soil with different textures under returned straw conditions. Chin. J. Eco-Agric. 2020, 28, 1061–1071. [Google Scholar]
- Harrison, M.J. The arbuscular mycorrhizal symbiosis: An underground association. Trends Plant Sci. 1997, 2, 54–60. [Google Scholar] [CrossRef]
- Allen, E.B.; West, N.E. Nontarget effects of the herbicide tebuthiuron on mycorrhizal fungi in sagebrush semidesert. Mycorrhiza 1993, 3, 75–78. [Google Scholar] [CrossRef]
- Brito, I.; Goss, M.J.; Alhoa, L.; Brigido, C.; van Tuinen, D.; Felix, M.; Carvalho, M. Agronomic management of AMF functional diversity to overcome biotic and abiotic stresses-The role of plant sequence and intact extraradical mycelium. Fungal Ecol. 2019, 40, 72–81. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Hashem, A.; Abd-Allah, E.F.; Ahmad, P. Arbuscular mycorrhiza in crop improvement under environmental stress. In Emerging Technologies and Management of Crop Stress Tolerance; Academic Press: Cambridge, MA, USA, 2014; pp. 69–95. [Google Scholar]
- Wang, X.X.; Li, H.; Chu, Q.; Feng, G.; Kuyper, T.W.; Rengel, Z. Mycorrhizal impacts on root trait plasticity of six maize varieties along a phosphorus supply gradient. Plant Soil 2020, 448, 71–86. [Google Scholar] [CrossRef]
- Jones, D.L.; Nguyen, C.; Finlay, R.D. Carbon flow in the rhizosphere: Carbon trading at the soil–root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Higo, M.; Azuma, M.; Kamiyoshihara, Y.; Kanda, A.; Tatewaki, Y.; Isobe, K. Impact of phosphorus fertilization on tomato growth and arbuscular mycorrhizal fungal communities. Microorganisms 2020, 8, 178. [Google Scholar] [CrossRef] [Green Version]
- Boomsma, C.R.; Vyn, T.J. Maize drought tolerance: Potential improvements through arbuscular mycorrhizal symbiosis. Field Crops Res. 2008, 108, 14–31. [Google Scholar] [CrossRef]
- Davies, F.T., Jr.; Potter, J.R.; Linderman, R.G. Mycorrhiza and repeated drought exposure affect drought resistance and extraradical hyphae development of pepper plants independent of plant size and nutrient content. J. Plant Physiol. 1992, 139, 289–294. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic insights into arbuscular mycorrhizal fungi-mediated drought stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.; Zou, Y.N.; Kuča, K.; Hashem, A.; Abd-Allah, E.F.; Wu, Q.-S. Elucidating the mechanisms underlying enhanced drought tolerance in plants mediated by arbuscular mycorrhizal fungi. Front. Microbiol. 2021, 12, 4029. [Google Scholar] [CrossRef] [PubMed]
- Climate Change 2007: Impacts, Adaptation and Vulnerability; Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007.
- Augé, R.M. Arbuscular mycorrhizae and soil/plant water relations. Can. J. Soil Sci. 2004, 84, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Liu, J.X.; Wang, X.L.; Li, L. Analysis of Rice Planting Changes and Influencing Factors in the Rice Grown Area along the Yellow River:Taking Yuanyang County of Henan Province as an example. Sci. Technol. Ind. 2021, 21, 109–113. [Google Scholar]
- Lu, R.K. Soil Agricultural Chemical Analysis Method; China Agricultural Science and Technology Press: Beijing, China, 2000; pp. 1–315. [Google Scholar]
- Hao, X.; Jiang, R.; Chen, T. Clustering 16S rRNA for OTU prediction: A method of unsupervised Bayesian clustering. Bioinformatics 2011, 27, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [Green Version]
- Beckers, B.; Op De Beeck, M.; Weyens, N.; Boerjan, W.; Vangronsveld, J. Structural variability and niche differentiation in the rhizosphere and endosphere bacterial microbiome of field-grown poplar trees. Microbiome 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ning, D.; Yang, Y.; He, N.; Li, X.; Cornell, C.R.; Bates, C.T.; Filimonenko, E.; Kuzyakov, Y.; Zhou, J.; et al. Precipitation balances deterministic and stochastic processes of bacterial community assembly in grassland soils. Soil Biol. Biochem. 2022, 168, 108635. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Wen, T.; Zhang, H.; Zhao, M.; Penton, C.R.; Thomashow, L.S.; Shen, Q. Predicting disease occurrence with high accuracy based on soil macroecological patterns of Fusarium wilt. ISME J. 2020, 14, 2936–2950. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.; Verbruggen, E.; Hu, Y.; Veresoglou, S.D.; Rillig, M.C.; Zhou, W.; Xu, T.; Li, H.; Hao, Z.; Chen, Y.; et al. Land use influences arbuscular mycorrhizal fungal communities in the farming–pastoral ecotone of northern China. New Phytol. 2014, 204, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Beals, E.W. Bray-Curtis ordination: An effective strategy for analysis of multivariate ecological data. Adv. Ecol. Res. 1984, 14, 1–55. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package 2.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the Third International Conference on Weblogs and Social Media, ICWSM 2009, San Jose, CA, USA, 17–20 May 2009; Volume 3, pp. 361–362. [Google Scholar] [CrossRef]
- Wang, W.; Luo, Z.; Zhou, R.; Xu, D.; Ai, J.; Ding, B. Habitat associations of woody plant species in Baishanzu subtropical broad-leaved evergreen forest. Biodivers. Sci. 2011, 19, 134–142. [Google Scholar] [CrossRef]
- Lan, G.; Getzin, S.; Wiegand, T.; Hu, Y.; Xie, G.; Zhu, H.; Cao, M. Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China. PLoS ONE 2012, 7, e46074. [Google Scholar] [CrossRef]
- Ning, D.; Deng, Y.; Tiedje, J.M.; Zhou, J. A general framework for quantitatively assessing ecological stochasticity. Proc. Natl. Acad. Sci. USA 2019, 116, 16892–16898. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Zheng, R.; Bai, S.L.; Wang, J.G.; Li, L.; Duan, G.Z. Advances of species diversity of arbuscular mycorrhizal fungi. Microbiol. China 2016, 43, 1836–1843. [Google Scholar]
- Guo, Y.; Du, Q.; Li, G.; Ni, Y.; Zhang, Z.; Ren, W.; Hou, X. Soil phosphorus fractions and arbuscular mycorrhizal fungi diversity following long-term grazing exclusion on semi-arid steppes in Inner Mongolia. Geoderma 2016, 269, 79–90. [Google Scholar] [CrossRef]
- Dai, X.L.; Wang, P.; Liu, R.R.; Hao, Y.B.; Ji, B.M. Effects of extreme drought on community composition of arbuscular mycorrhizal fungi in the typical grasslands in Inner Mongolia during different growing seasons. Pratacultural Sci. 2020, 37, 1440–1447. [Google Scholar]
- Zhang, Z.; Zhang, J.; Xu, G.; Zhou, L.; Li, Y. Arbuscular mycorrhizal fungi improve the growth and drought tolerance of Zenia insignis seedlings under drought stress. New For. 2019, 50, 593–604. [Google Scholar] [CrossRef]
- Ma, S.; Chen, X.; Su, H.; Xing, A.; Chen, G.; Zhu, J.; Zhu, B.; Fang, J. Phosphorus addition decreases soil fungal richness and alters fungal guilds in two tropical forests. Soil Biol. Biochem. 2022, 175, 108836. [Google Scholar] [CrossRef]
- Li, J. The Regulation Mechanism of Arbuscular Mycorrhizal Fungi on Carbon and Nitrogen Turnover under Drought Stress in Songnen Grassland. Ph.D. Thesis, The Northeast Normal University, Changchun, China, 2022. [Google Scholar] [CrossRef]
- Zhao, Q.; Jian, S.; Nunan, N.; Maestre, F.T.; Tedersoo, L.; He, J.; Wei, H.; Tan, X.; Shen, W. Altered precipitation seasonality impacts the dominant fungal but rare bacterial taxa in subtropical forest soils. Biol. Fertil. Soils 2017, 53, 231–245. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miransari, M. Arbuscular Mycorrhizal Fungi and Uptake of Nutrients. In Symbiotic Endophytes; Soil Biology; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 37. [Google Scholar]
- Williams, A.; Manoharan, L.; Rosenstock, N.P.; Olsson, P.A.; Hedlund, K. Long-term agricultural fertilization alters arbuscular mycorrhizal fungal community composition and barley (Hordeum vulgare) mycorrhizal carbon and phosphorus exchange. New Phytol. 2017, 213, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Chen, B.; Roos, P.; Borggaard, O.K.; Zhu, Y.; Jakobsen, I. Mycorrhiza and root hairs in barley enhance acquisition of phosphorus and uranium from phosphate rock but mycorrhiza decreases root to shoot uranium transfer. New Phytol. 2005, 165, 591–598. [Google Scholar] [CrossRef]
- Feng, H.; Feng, G.; Wang, J.; Li, X. Regulation of P status in host plant on alkaline phosphatase (ALP) activity in intraradical hyphae and development of extraradical hyphae of AM fungi. Jun Wu Xi Tong Mycosystema 2003, 22, 589–598. [Google Scholar]
- Dini-Andreote, F.; Stegen, J.C.; Van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yin, W.; Wang, C.; Zhang, A.; Zhang, H.; Zhang, T.; Ju, F. Untangling microbiota diversity and assembly patterns in the world’s largest water diversion canal. Water Res. 2021, 204, 117617. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-Y.; Sun, L.-Y.; Song, F.-B.; Yang, X.-Q.; Zhang, M.-J.; Li, S.-X.; Zhu, X.-C. Research advance in species diversity of arbuscular mycorrhizal fungi in terrestrial agricultural ecosystems. J. Appl. Ecol. 2019, 30, 3971. [Google Scholar]
- Xu, J.; Liu, S.; Song, S.; Guo, H.; Tang, J.; Yong, J.W.H.; Ma, Y.; Chen, X. Arbuscular mycorrhizal fungi influence decomposition and the associated soil microbial community under different soil phosphorus availability. Soil Biol. Biochem. 2018, 120, 181–190. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Soil Nutrient Content | |||
|---|---|---|---|
| Group | The Effective Phosphorus (P) (mg/kg) | The Effective Potassium (K) (mg/kg) | The Nitrate Nitrogen (N) (mg/kg) |
| DLP | 1.7 ± 0.1 | 21.7 ± 4.3 | 15.1 ± 2.1 |
| DNP | 6.6 ± 0.7 | 33.0 ± 10.3 | 16.1 ± 1.4 |
| NNP | 12.3 ± 2.0 | 19.7 ± 5.9 | 18.8 ± 3.6 |
| NLP | 4.2 ± 1.8 | 15.4 ± 2.9 | 12.4 ± 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Wang, Z.; Liu, W.; Zhang, X.; Wang, J.; Guo, P. Effects of Variations in Soil Moisture and Phosphorus Concentrations on the Diversity of the Arbuscular Mycorrhizal Fungi Community in an Agricultural Ecosystem. Agriculture 2023, 13, 1272. https://doi.org/10.3390/agriculture13061272
Shao Y, Wang Z, Liu W, Zhang X, Wang J, Guo P. Effects of Variations in Soil Moisture and Phosphorus Concentrations on the Diversity of the Arbuscular Mycorrhizal Fungi Community in an Agricultural Ecosystem. Agriculture. 2023; 13(6):1272. https://doi.org/10.3390/agriculture13061272
Chicago/Turabian StyleShao, Yizhen, Zhao Wang, Wenjun Liu, Xintong Zhang, Jing Wang, and Peng Guo. 2023. "Effects of Variations in Soil Moisture and Phosphorus Concentrations on the Diversity of the Arbuscular Mycorrhizal Fungi Community in an Agricultural Ecosystem" Agriculture 13, no. 6: 1272. https://doi.org/10.3390/agriculture13061272