
Study on Phyllosphere Microbial Community of Nettle Leaf during Different Seasons
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Materials Preparation
2.3. Amino Acids Content Analysis
2.4. Bacterial and Fungal Community Sequencing Analysis
3. Statistical Analysis
4. Results
4.1. Amino Acid Contents of Nettle Leaves under Seasonal Variation
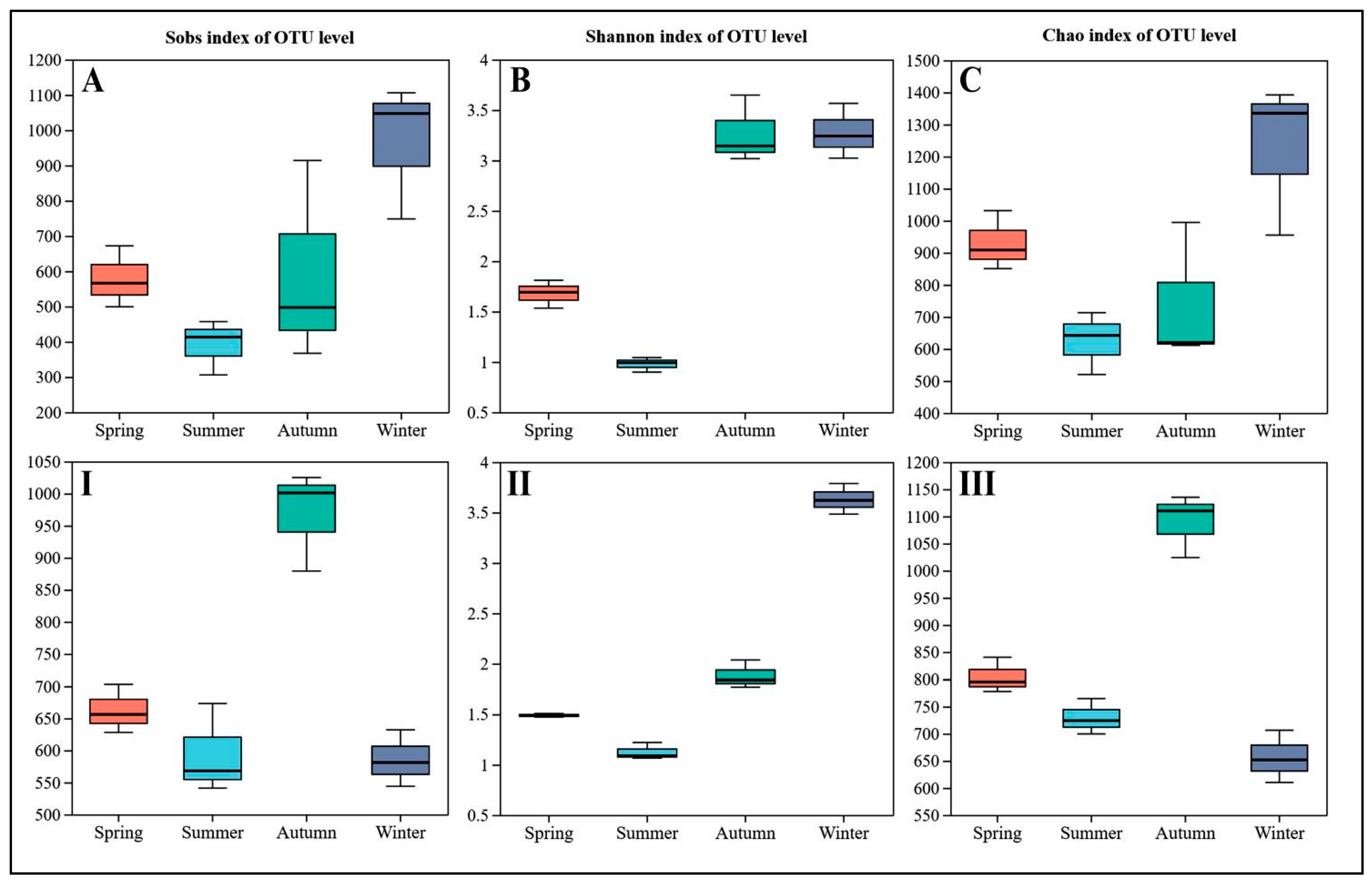
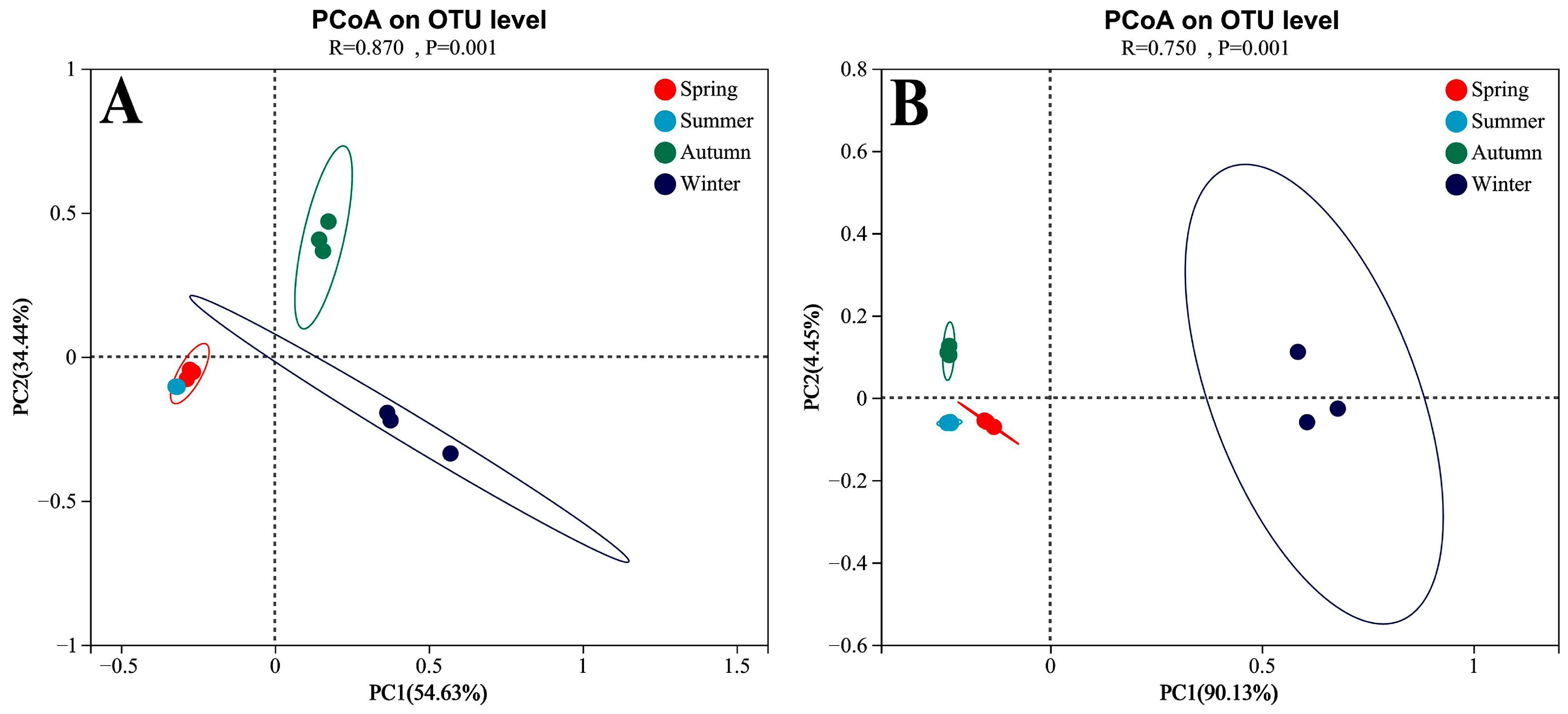
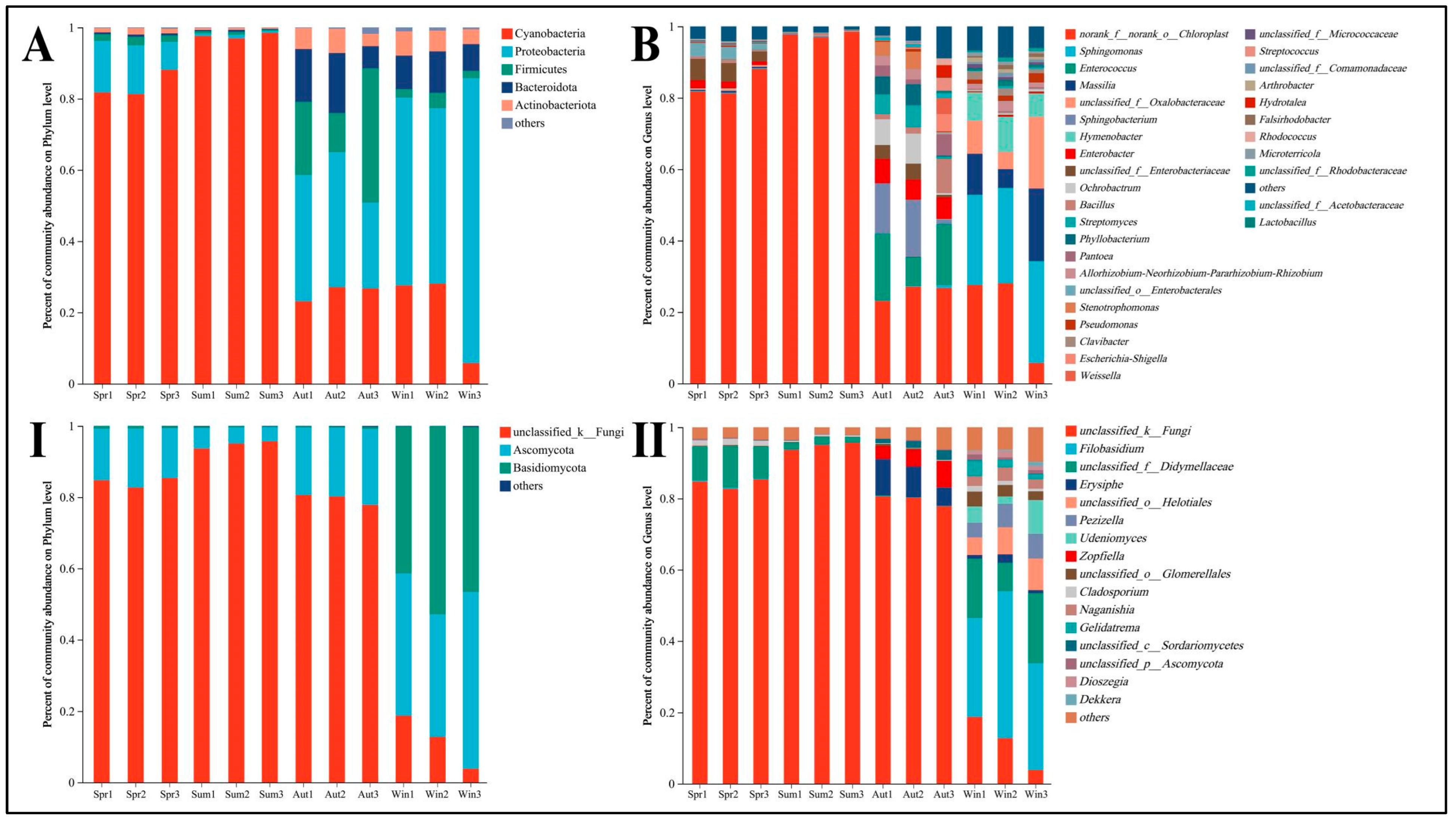
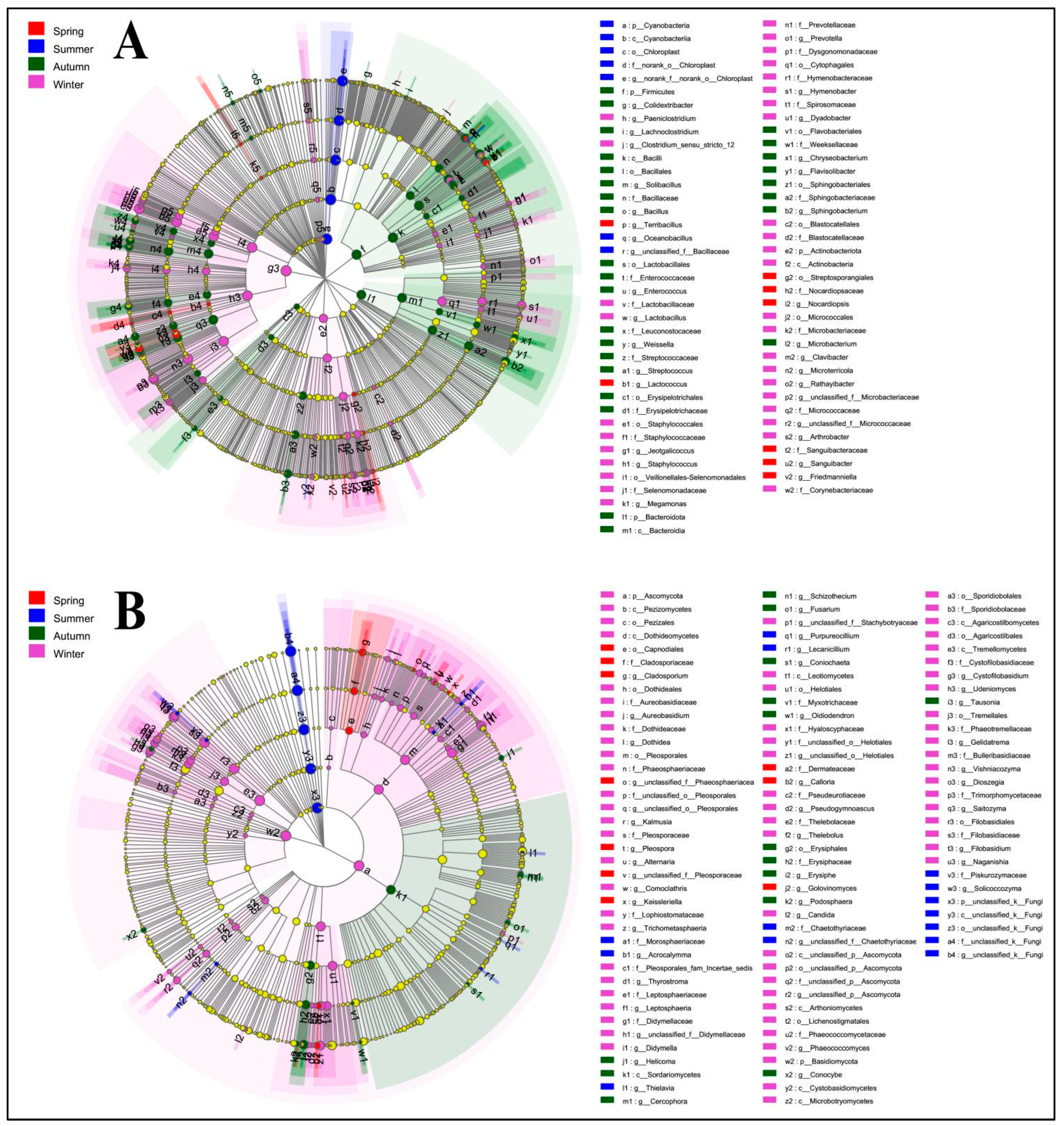
4.2. Phyllosphere Microbe Composition across Seasons
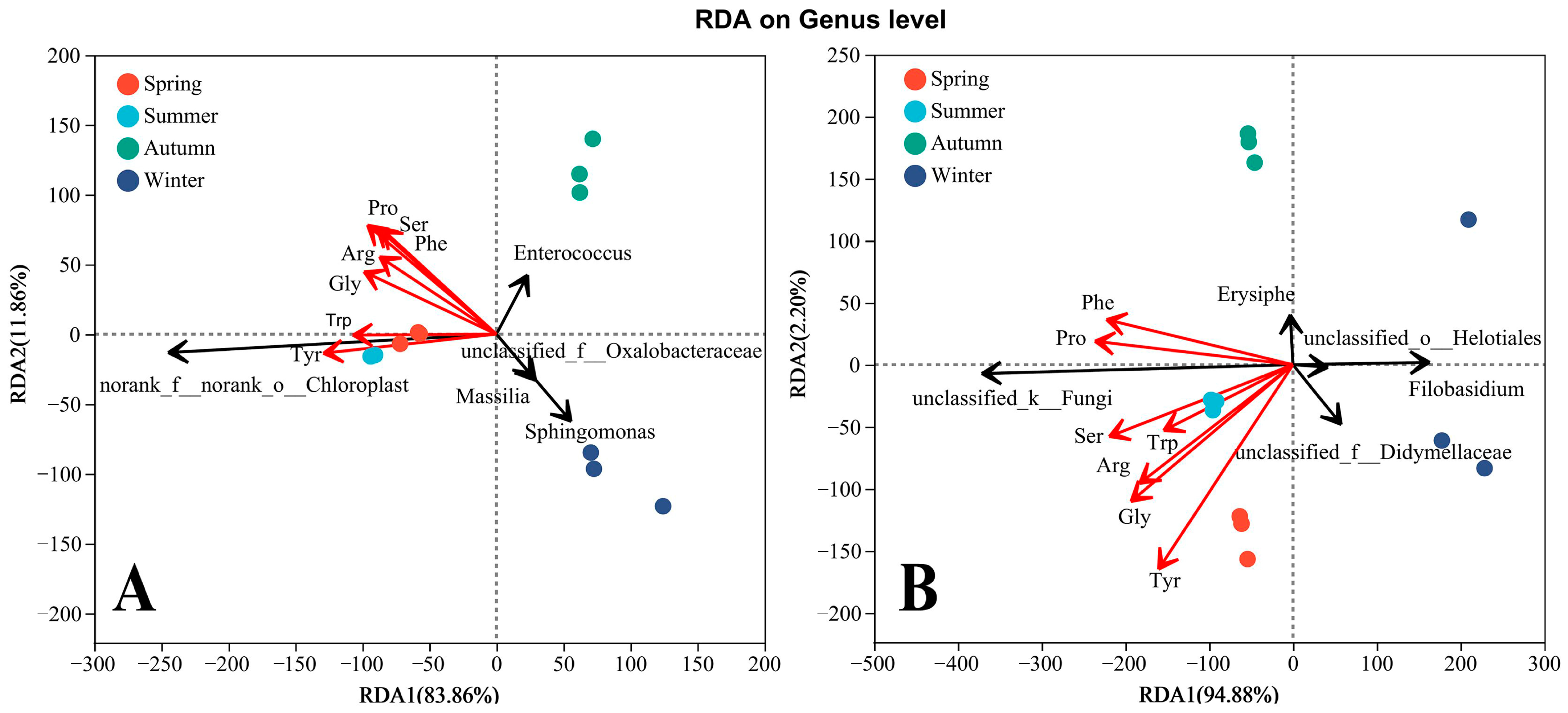
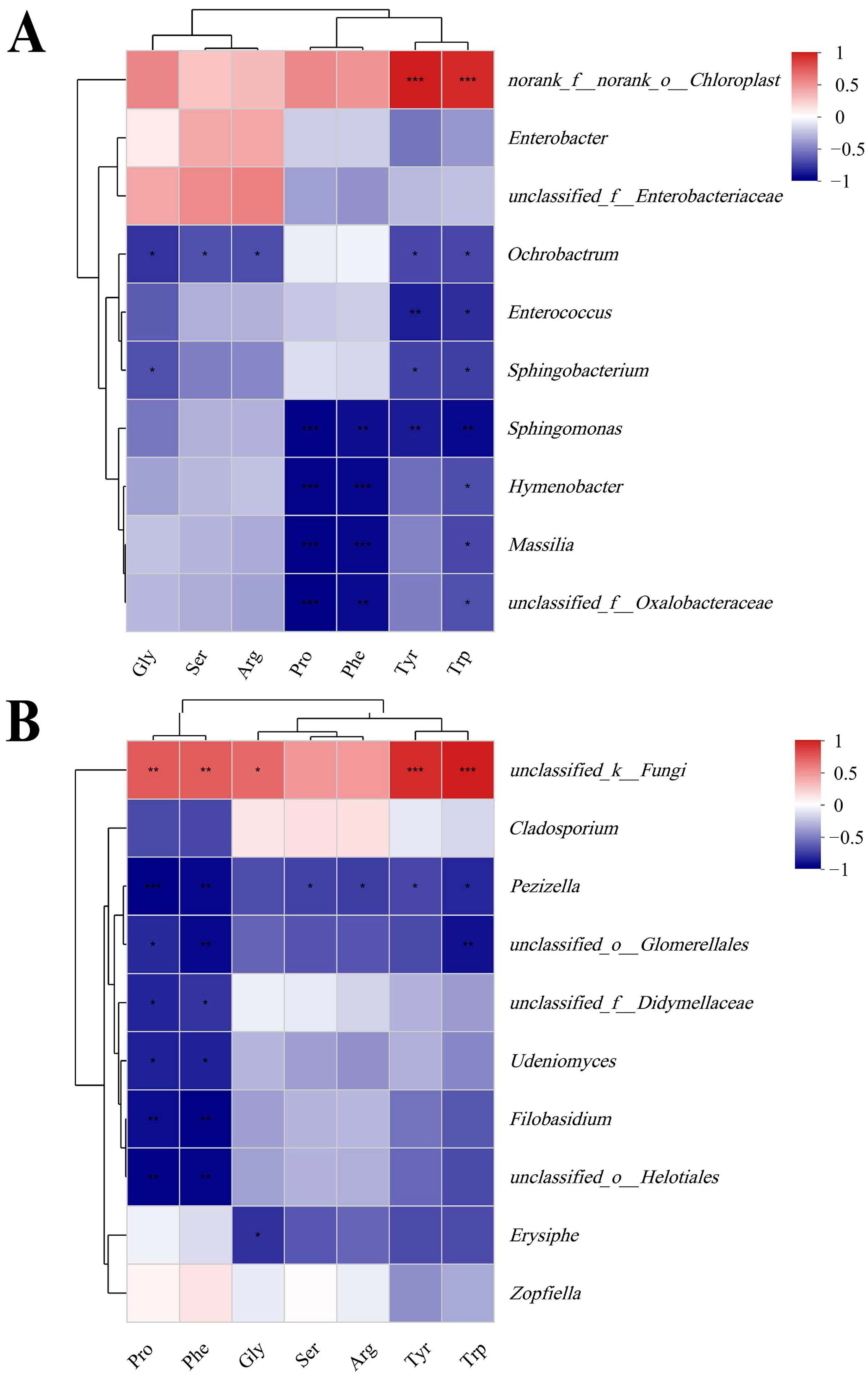
4.3. Relationship between Phyllosphere Microbial Composition and Amino Acids
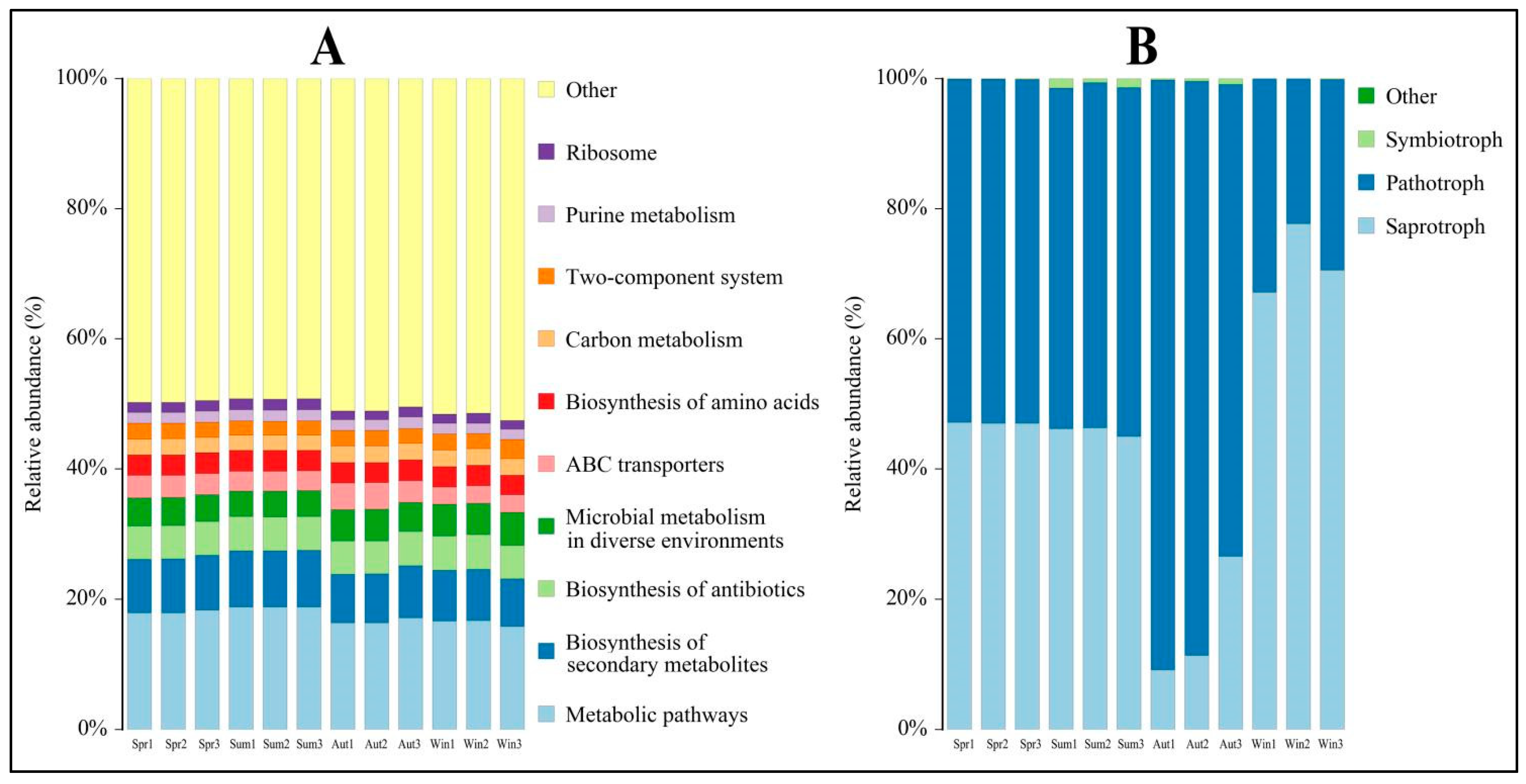
4.4. Functional Prediction of Phyllosphere Microbes
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Becker, K.; Grosse-Veldmann, B.; Weigend, M. Weeding the nettles V: Taxonomic and phylogenetic studies of the eastern Asian species Urtica thunbergiana Sieb. & Zucc. (Urticaceae). Phytotaxa 2017, 323, 201–216. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Jin, Y.M. Nutritive Value of Nettle and its Processing and Storage Technology; China Agricultural Science and Technology Press: Beijing, China, 2015. [Google Scholar]
- Li, R.; Qin, M.J.; Yu, G.D. The research outline and the developing value of Urtica. China Wild Plant Resour. 2002, 21, 24–26. (In Chinese) [Google Scholar]
- Wu, X.Z.; Liu, B.R.; Yan, X.; Liu, R.T.; An, H. Response of soil microbial biomass and microbial entropy to desertification in desert grassland. Chin. J. Appl. Ecol. 2019, 30, 2691–2698. [Google Scholar] [CrossRef]
- Karunarathna, A.; Tibpromma, S.; Jayawardena, R.S.; Nanayakkara, C.; Asad, S.; Xu, J.; Hyde, K.D.; Karunarathna, S.C.; Stephenson, S.L.; Lumyong, S.; et al. Fungal Pathogens in Grasslands. Front. Cell. Infect. Microbiol. 2021, 11, 695087. [Google Scholar] [CrossRef] [PubMed]
- Dondoni, A.; Massi, A. Design and Synthesis of New Classes of Heterocyclic C-Glycoconjugates and Carbon-Linked Sugar and Heterocyclic Amino Acids by Asymmetric Multicomponent Reactions (AMCRs). Cheminform 2006, 39, 451–463. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, D.; Liu, Q. Connections Between Amino Acid Metabolisms in Plants: Lysine as an Example. Front. Plant Sci. 2020, 11, 928. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Amir, R.; Galili, G.; Cohen, H. The metabolic roles of free amino acids during seed development. Plant Sci. 2018, 275, 11–18. [Google Scholar] [CrossRef]
- Yang, Q.-Q.; Zhao, D.-S.; Zhang, C.-Q.; Wu, H.-Y.; Li, Q.-F.; Gu, M.-H.; Sun, S.S.-M.; Liu, Q.-Q. A Connection between Lysine and Serotonin Metabolism in Rice Endosperm. Plant Physiol. 2018, 176, 1965–1980. [Google Scholar] [CrossRef] [Green Version]
- Galili, G.; Amir, R.; Fernie, A.R. The Regulation of Essential Amino Acid Synthesis and Accumulation in Plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [Green Version]
- Dlamini, S.P.; Akanmu, A.O.; Babalola, O.O. Rhizospheric microorganisms: The gateway to a sustainable plant health. Front. Sustain. Food Syst. 2022, 6, 925802. [Google Scholar] [CrossRef]
- Xu, N.; Zhao, Q.; Zhang, Z.; Zhang, Q.; Wang, Y.; Qin, G.; Ke, M.; Qiu, D.; Peijnenburg, W.J.G.M.; Lu, T.; et al. Phyllosphere Microorganisms: Sources, Drivers, and Their Interactions with Plant Hosts. J. Agric. Food Chem. 2022, 70, 4860–4870. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Xu, L.; Montoya, L.; Madera, M.; Hollingsworth, J.; Chen, L.; Purdom, E.; Singan, V.; Vogel, J.; Hutmacher, R.B.; et al. Co-occurrence networks reveal more complexity than community composition in resistance and resilience of microbial communities. Nature Communications 2022, 13, 3867. [Google Scholar] [CrossRef] [PubMed]
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.M.; Leveau, J.H.J. Microbiology of the phyllosphere: A playground for testing ecological concepts. Oecologia 2011, 168, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, A.; Halverson, L.J.; Beattie, G.A. Bacterial degradation of airborne phenol in the phyllosphere. Environ. Microbiol. 2010, 9, 383–392. [Google Scholar] [CrossRef]
- Innerebner, G.; Knief, C.; Vorholt, J.A. Protection of Arabidopsis thaliana against Leaf-Pathogenic Pseudomonas syringae by Sphingomonas Strains in a Controlled Model System. Appl. Environ. Microbiol. 2011, 77, 3202–3210. [Google Scholar] [CrossRef] [Green Version]
- Opacic, N.; Radman, S.; Uher, S.F.; Benko, B.; Voca, S.; Zlabur, J.S. Nettle cultivation practices-from open field to modern hydroponics: A case study of specialized metabolites. Plants 2022, 11, 483. [Google Scholar] [CrossRef]
- Simancas, B.; Juvany, M.; Cotado, A.; Munne-Bosch, S. Sex-related differences in photoinhibition, photo-oxidative stress and photoprotection in stinging nettle (Urtica dioica L.) exposed to drought and nutrient deficiency. J. Photochem. Photobiol. B-Biol. 2016, 156, 22–28. [Google Scholar] [CrossRef]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyilosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Agler, M.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.-T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishore, G.K.; Pande, S.; Podile, A.R. Biological Control of Late Leaf Spot of Peanut (Arachis hypogaea) with Chitinolytic Bacteria. Phytopathology 2005, 95, 1157–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivakumar, N.; Sathishkumar, R.; Selvakumar, G.; Shyamkumar, R.; Arjunekumar, K. Phyllospheric microbiomes: Diversityecological significance, and biotechmological applications. Plant Microbiomes Sustain. Agric. 2020, 25, 113–172. [Google Scholar]
- Moormann, J.; Heinemann, B.; Hildebrandt, T.M. News about amino acid metabolism in plant–microbe interactions. Trends Biochem. Sci. 2022, 47, 839–850. [Google Scholar] [CrossRef]
- Moore, S.; Stein, W.H. Chromatographic determination of amino acids by the use of automatic recording equipment. Method Enzymol. 1963, 6, 819–831. [Google Scholar]
- Schloss, P.D. A High-Throughput DNA Sequence Aligner for Microbial Ecology Studies. PLoS ONE 2009, 4, e8230. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, I.; Shahid, M.A.; Babar, A. Role of sugars, amino acids and organic acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2019, 52, 355–363. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Rathinasabapathi, B.; Babar, A. UPLC-HRMS-based untargeted metabolic profiling reveals changes in chickpea (Cicer arietinum) metabolome following long-term drought stress. Plant Cell Environ. 2018, 42, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Obata, T.; Witt, S.; Lisec, J.; Palacios-Rojas, N.; Florez-Sarasa, I.; Yousfi, S.; Araus, J.L.; Cairns, J.E.; Fernie, A.R. Metabolite profiles of maize leaves in drought, heat and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physiol. 2015, 169, 2665–2683. [Google Scholar] [CrossRef] [Green Version]
- Suguiyama, V.F.; da Silva, E.A.; Meirelles, S.T.; Centeno, D.D.C.; Braga, M.R. Leaf metabolite profile of the Brazilian resurrection plant Barbacenia purpurea Hook. (Velloziaceae) shows two time-dependent responses during desiccation and recovering. Front. Plant Sci. 2014, 5, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fait, A.; Yellin, A.; Fromm, H. GABA shunt deficiencies and accumulation of reactive oxygen intermediates: Insight from Arabidopsis mutants. FEBS Lett. 2005, 579, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, N. Constraints placed by community diversity on the enzymatic response of microbial decomposer communities to climate change in southern california. In Proceedings of the Agu Fall Meeting AGU Fall Meeting Abstracts, Union City, CA, USA, 14–18 December 2015. [Google Scholar]
- Zhang, Y.; Xie, Y.-Z.; Ma, H.-B.; Zhang, J.; Jing, L.; Wang, Y.-T.; Li, J.-P. Responses of soil microorganisms to simulated climate change in desert grassland in northern China. J. Water Clim. Chang. 2022, 13, 1842–1854. [Google Scholar] [CrossRef]
- Chaudhry, V.; Runge, P.; Sengupta, P.; Doehlemann, G.; Parker, J.E.; Kemen, E. Shaping the leaf microbiota: Plant–microbe–microbe interactions. J. Exp. Bot. 2020, 72, 36–56. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Haney, C.H. Drought dampens microbiome development. Nat. Plants 2021, 7, 994–995. [Google Scholar] [CrossRef]
- Lombardino, J.; Bijlani, S.; Singh, N.K.; Wood, J.M.; Barker, R.; Gilroy, S.; Wang, C.C.C.; Venkateswaran, K. Genomic Characterization of Potential Plant Growth-Promoting Features of Sphingomonas Strains Isolated from the International Space Station. Microbiol. Spectr. 2022, 10, e01994-21. [Google Scholar] [CrossRef]
- Adhikari, T.B.; Joseph, C.M.; Yang, G.; Phillips, D.A.; Nelson, L.M. Evaluation of bacteria isolated from rice for plant growth promotion and biological control of seedling disease of rice. Can. J. Microbiol. 2001, 47, 916–924. [Google Scholar] [CrossRef]
- Orthová, I.; Kampfer, P.; Glaeser, S.P.; Kaden, R.; Busse, H.-J. Massilia norwichensis sp. nov., isolated from an air sample. Int. J. Syst. Evol. Microbiol. 2015, 65, 56–64. [Google Scholar] [CrossRef]
- Zheng, B.-X.; Bi, Q.-F.; Hao, X.-L.; Zhou, G.; Yang, X.-R. Massilia phosphatilytica sp. nov., a phosphate solubilizing bacteria isolated from a long-term fertilized soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2514–2519. [Google Scholar] [CrossRef]
- Terashima, M.; Ohashi, K.; Takasuka, T.E.; Kojima, H.; Fukui, M. Antarctic heterotrophic bacterium Hymenobacter nivis P3T displays light-enhanced growth and expresses putative photoactive proteins. Environ. Microbiol. Rep. 2019, 11, 227–235. [Google Scholar] [CrossRef]
- Glushakova, A.M.; Kachalkin, A.V. Endophytic yeasts in Malus domestica and Pyrus communis fruits under anthropogenic impact. Microbiology 2017, 3, 128–135. [Google Scholar] [CrossRef]
- Singh, H. A Novel Radiation-Resistant Strain of Filobasidium sp. Isolated from the West Sea of Korea. J. Microbiol. Biotechnol. 2013, 23, 1493–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, D. Pezizella ericae sp.nov., the perfect state of a typical mycorrhizal endophyte of ericaceae. Trans. Br. Mycol. Soc. 1974, 63, 381–383. [Google Scholar] [CrossRef]
- Li, G.; Kusari, S.; Lamshöft, M.; Schüffler, A.; Laatsch, H.; Spiteller, M. Antibacterial Secondary Metabolites from an Endophytic Fungus, Eupenicillium sp. LG41. J. Nat. Prod. 2014, 77, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Ferus, P.; Barta, M.; Konôpková, J. Endophytic fungus beauveria bassiana can enhance drought tolerance in red oak Seedlings. Trees 2019, 33, 1179–1186. [Google Scholar] [CrossRef]
- Bradshaw, M.; Braun, U.; Götz, M.; Jurick, W., ΙΙ. Phylogeny and taxonomy of powdery mildew caused by Erysiphe species on Lupinus hosts. Mycologia 2021, 114, 76–88. [Google Scholar] [CrossRef]
- Takeuchi, M.; Sawada, H.; Oyaizu, H.; Yokota, A.; Jiang, C.; Xu, L.; Yang, Y.-R.; Guo, G.-Y.; Ma, J.; Liu, Y. Phylogenetic Evidence for Sphingomonas and Rhizomonas as Nonphotosynthetic Members of the Alpha-4 Subclass of the Proteobacteria. Int. J. Syst. Evol. Microbiol. 1994, 44, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ge, C.; Xu, S.; Wu, Y.; Sahito, Z.A.; Ma, L.; Pan, F.; Zhou, Q.; Huang, L.; Feng, Y.; et al. The endophytic bacterium Sphingomonas SaMR12 alleviates Cd stress in oilseed rape through regulation of the GSH-AsA cycle and antioxidative enzymes. BMC Plant Biol. 2020, 20, 63. [Google Scholar] [CrossRef] [Green Version]
- Markovska, Y.K.; Gorinova, N.I.; Nedkovska, M.P.; Miteva, K.M. Cadmium-induced oxidative damage and antioxidant responses in Brassica juncea plants. Biol. Plant. 2009, 53, 151–154. [Google Scholar] [CrossRef]
- Liang, Q.; Lloyd-Jones, G. Sphingobium scionense sp. nov., an aromatic hydrocarbon-degrading bacterium isolated from contaminated sawmill soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 413–416. [Google Scholar] [CrossRef]
- Chiacchio, M.; Wild, M. Influence of NAO and clouds on long-term seasonal variations of surface solar radiation in Europe. J. Geophys. Res. Atmos. 2010, 115, D00D22. [Google Scholar] [CrossRef]
- Yan, T.J.; Zheng, L.L.; Li, X.N.; Liu, L.P.; Pan, Y.; Wang, J.; Long, F.; Zhou, P.; Shi, X.D. Bacterial diversity and function prediction of cigar tobacco leaves based on high-throughput sequencing. Acta Tab. Sin. 2022, 28, 125–132. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids (AA) % | Spring | Summer | Autumn | Winter | SEM | p Value | |
|---|---|---|---|---|---|---|---|
| Non-essential AA | Alanine | 1.83 a | 1.26 b | 0.92 c | 0.51 d | 0.15 | *** |
| Arginine | 2.18 a | 1.56 b | 1.57 b | 1.04 c | 0.12 | ** | |
| Aspartate | 2.11 a | 1.82 b | 1.55 c | 1.24 d | 0.10 | *** | |
| Cysteine | 1.29 a | 1.10 b | 0.94 c | 0.62 d | 0.07 | *** | |
| Glutamate | 1.86 a | 1.61 b | 1.32 c | 0.91 d | 0.11 | *** | |
| Glycine | 0.82 a | 0.71 b | 0.64 c | 0.49 d | 0.04 | *** | |
| Histidine | 1.20 a | 0.88 b | 0.64 b | 0.51 c | 0.08 | ** | |
| Proline | 1.68 c | 2.12 a | 1.79 b | 1.00 d | 0.12 | *** | |
| Serine | 2.13 a | 1.75 b | 1.77 b | 1.04 c | 0.12 | ** | |
| Tyrosine | 1.13 b | 1.22 a | 0.03 c | 0.02 c | 0.17 | ** | |
| Essential AA | Isoleucine | 0.51 b | 0.62 a | 0.42 b | 0.21 c | 0.05 | ** |
| Leucine | 2.12 b | 2.62 a | 2.00 b | 1.34 c | 0.14 | ** | |
| Lysine | 1.94 a | 1.59 b | 1.62 b | 1.15 c | 0.09 | ** | |
| Methionine | 1.38 a | 1.21 b | 1.04 c | 0.82 d | 0.06 | *** | |
| Phenylalanine | 1.70 c | 2.48 a | 2.04 b | 1.01 d | 0.16 | *** | |
| Threonine | 0.97 b | 1.18 a | 0.83 c | 0.42 d | 0.08 | *** | |
| Tryptophan | 0.42 b | 0.79 a | 0.35 b c | 0.29 c | 0.06 | *** | |
| Valine | 0.11 b | 0.30 a | 0.12 b | 0.06 c | 0.03 | *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, S.; Chen, Y.; Huang, R.; Chai, Y.; Ma, C.; Zhang, F. Study on Phyllosphere Microbial Community of Nettle Leaf during Different Seasons. Agriculture 2023, 13, 1271. https://doi.org/10.3390/agriculture13061271
Jia S, Chen Y, Huang R, Chai Y, Ma C, Zhang F. Study on Phyllosphere Microbial Community of Nettle Leaf during Different Seasons. Agriculture. 2023; 13(6):1271. https://doi.org/10.3390/agriculture13061271
Chicago/Turabian StyleJia, Shuan, Yongcheng Chen, Rongzheng Huang, Yuxin Chai, Chunhui Ma, and Fanfan Zhang. 2023. "Study on Phyllosphere Microbial Community of Nettle Leaf during Different Seasons" Agriculture 13, no. 6: 1271. https://doi.org/10.3390/agriculture13061271