
Estimating Freezing Injury on Olive Trees: A Comparative Study of Computing Models Based on Electrolyte Leakage and Tetrazolium Tests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Olive Cultivars and Orchard Management
2.2. Temperature Treatments
2.3. Electrolyte Leakage (EL)
2.4. Triphenyl Tetrazolium Chloride (TZ) Assay
2.5. Nonlinear Regression Models (NLRs)
3. Results and Discussion
3.1. Findings of the Electrolyte Leakage (EL) Assay
3.1.1. Comparing and Selecting the Best-Fitted NLR Model for EL
3.1.2. Assessment of NLR Model Coefficients for EL
3.1.3. Conducting a Sensitivity Analysis of the EL Model
3.1.4. Comparison of EL Modeling Outcomes for Different Olive Cultivars
3.1.5. Analysis of the Rate of Change in EL Results
3.2. Findings of the Triphenyl Tetrazolium Chloride (TZ) Assay
3.2.1. Comparing and Selecting the Best-Fitted NLR Model for TZ
3.2.2. Assessment of NLR Model Coefficients for TZ
3.2.3. Conducting a Sensitivity Analysis of the TZ Model
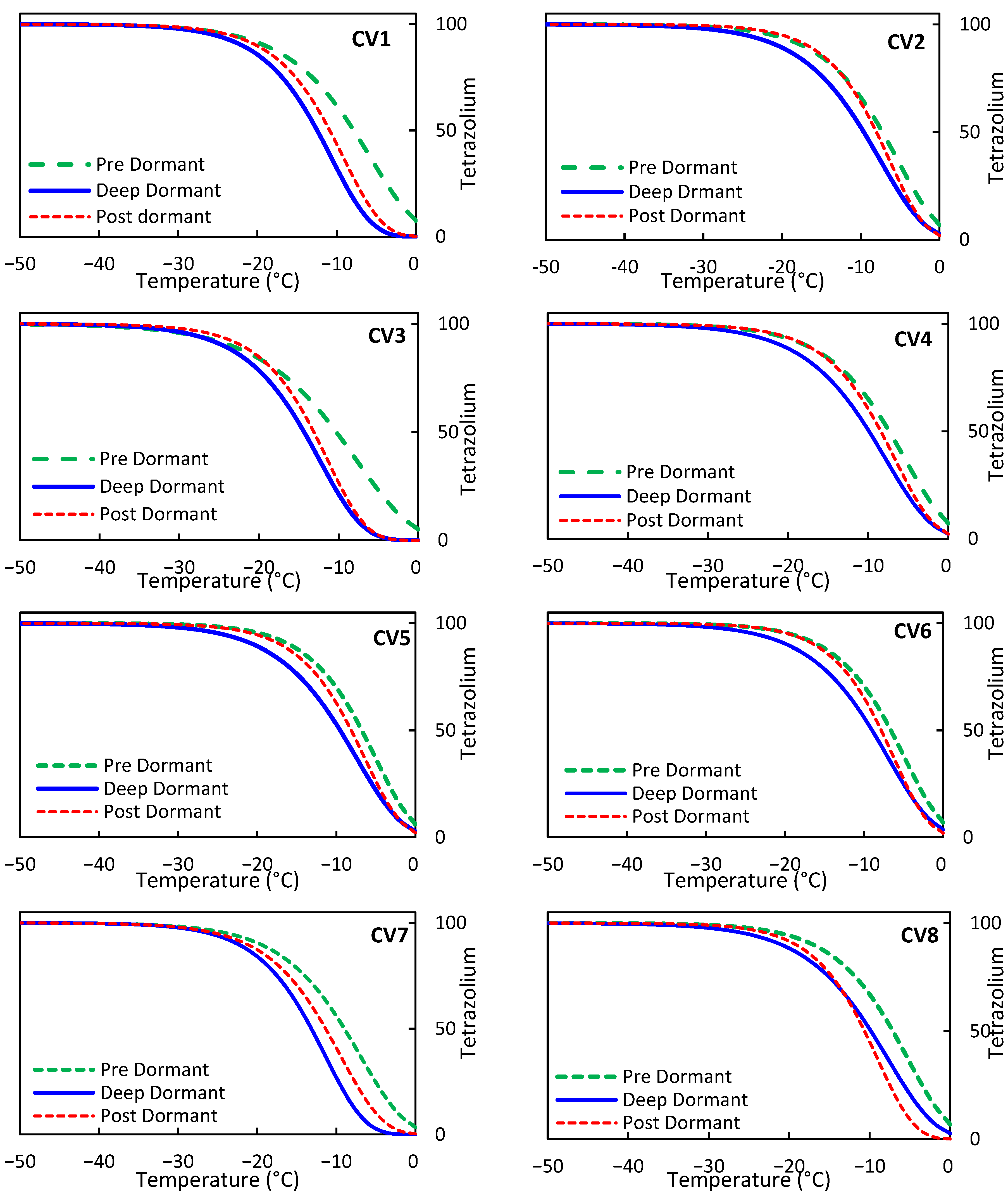
3.2.4. Comparison of TZ Modeling Outcomes across Various Olive Cultivars
3.2.5. Analysis of the Rate of Change in TZ Results
3.3. Assessment of T50 and T90 Values Based on the TZ and EL Models in Different Olive Varieties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dahdouh, A.; Khay, I.; Le Brech, Y.; El Maakoul, A.; Bakhouya, M. Olive oil industry: A review of waste stream composition, environmental impacts, and energy valorization paths. Environ. Sci. Pollut. Res. 2023, 30, 45473–45497. [Google Scholar] [CrossRef] [PubMed]
- Petruccelli, R.; Bartolini, G.; Ganino, T.; Zelasco, S.; Lombardo, L.; Perri, E.; Durante, M.; Bernardi, R. Cold Stress, Freezing Adaptation, Varietal Susceptibility of Olea europaea L.: A Review. Plants 2022, 11, 1367. [Google Scholar] [CrossRef]
- Fraga, H.; Moriondo, M.; Leolini, L.; Santos, J.A. Mediterranean olive orchards under climate change: A review of future impacts and adaptation strategies. Agronomy 2020, 11, 56. [Google Scholar] [CrossRef]
- Bartolozzi, F.; Fontanazza, G. Assessment of frost tolerance in olive (Olea europaea L.). Sci. Hortic. 1999, 81, 309–319. [Google Scholar] [CrossRef]
- Cansev, A.; Gulen, H.; Eris, A. Cold-hardiness of olive (Olea europaea L.) cultivars in cold-acclimated and non-acclimated stages: Seasonal alteration of antioxidative enzymes and dehydrin-like proteins. J. Agric. Sci. 2009, 147, 51–61. [Google Scholar] [CrossRef]
- Lodolini, E.M.; Alfei, B.; Santinelli, A.; Cioccolanti, T.; Polverigiani, S.; Neri, D. Frost tolerance of 24 olive cultivars and subsequent vegetative re-sprouting as indication of recovery ability. Sci. Hortic. 2016, 211, 152–157. [Google Scholar] [CrossRef]
- López-Bernal, Á.; García-Tejera, O.; Testi, L.; Orgaz, F.; Villalobos, F.J. Studying and modelling winter dormancy in olive trees. Agric. For. Meteorol. 2020, 280, 107776. [Google Scholar] [CrossRef]
- Yuldasheva, K.T.; Soliyeva, M.; Kimsanova, X.; Arabboev, A.; Kayumova, S. Evaluation of winter frost resistance of cultivated varieties of olives. Acad. Int. Multidiscip. Res. J. 2021, 11, 627–632. [Google Scholar] [CrossRef]
- Gómez-del-Campo, M.; Barranco, D. Field evaluation of frost tolerance in 10 olive cultivars. Plant Genet. Resour. 2005, 3, 385–390. [Google Scholar] [CrossRef]
- Lindén, L.; Palonen, P.; Lindén, M. Relating freeze-induced electrolyte leakage measurements to lethal temperature in red raspberry. J. Am. Soc. Hortic. Sci. 2000, 125, 429–435. [Google Scholar] [CrossRef]
- Murray, M.; Cape, J.; Fowler, D. Quantification of frost damage in plant tissues by rates of electrolyte leakage. New Phytol. 1989, 113, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.J.; Lee, H.J. Evaluation of freezing injury in temperate fruit trees. Hortic. Environ. Biotechnol. 2020, 61, 787–794. [Google Scholar] [CrossRef]
- Whitlow, T.H.; Bassuk, N.L.; Ranney, T.G.; Reichert, D.L. An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiol. 1992, 98, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Karami, H.; Rezaei, M.; Sarkhosh, A.; Rahemi, M.; Jafari, M. Cold hardiness assessment in seven commercial fig cultivars (Ficus carica L.). Gesunde Pflanz. 2018, 70, 195–203. [Google Scholar] [CrossRef]
- Saadati, S.; Baninasab, B.; Mobli, M.; Gholami, M. Measurements of freezing tolerance and their relationship with some biochemical and physiological parameters in seven olive cultivars. Acta Physiol. Plant. 2019, 41, 51. [Google Scholar] [CrossRef]
- Lim, C.C.; Arora, R.; Townsend, E.C. Comparing Gompertz and Richards functions to estimate freezing injury in Rhododendron using electrolyte leakage. J. Am. Soc. Hortic. Sci. 1998, 123, 246–252. [Google Scholar] [CrossRef]
- Tudela, V.; Santibáñez, F. Modelling impact of freezing temperatures on reproductive organs of deciduous fruit trees. Agric. For. Meteorol. 2016, 226–227, 28–36. [Google Scholar] [CrossRef]
- Villouta, C.; Workmaster, B.A.; Atucha, A. Freezing stress damage and growth viability in Vaccinium macrocarpon Ait. bud structures. Physiol. Plant. 2021, 172, 2238–2250. [Google Scholar] [CrossRef]
- Archontoulis, S.V.; Miguez, F.E. Nonlinear regression models and applications in agricultural research. Agron. J. 2015, 107, 786–798. [Google Scholar] [CrossRef]
- Li, Q.; Gao, H.; Zhang, X.; Ni, J.; Mao, H. Describing Lettuce Growth Using Morphological Features Combined with Nonlinear Models. Agronomy 2022, 12, 860. [Google Scholar] [CrossRef]
- Jannatizadeh, A.; Rezaei, M.; Rohani, A.; Lawson, S.; Fatahi, R. Towards modeling growth of apricot fruit: Finding a proper growth model. Hortic. Environ. Biotechnol. 2022, 64, 209–222. [Google Scholar] [CrossRef]
- Connor, D.J.; Fereres, E. The physiology of adaptation and yield expression in olive. Hortic. Rev. 2010, 31, 155–229. [Google Scholar]
- Sanzani, S.M.; Schena, L.; Nigro, F.; Sergeeva, V.; Ippolito, A.; Salerno, M.G. Abiotic diseases of olive. J. Plant Pathol. 2012, 94, 469–491. [Google Scholar]
- Larcher, W. Temperature stress and survival ability of Mediterranean sclerophyllous plants. Plant Biosyst. 2000, 134, 279–295. [Google Scholar] [CrossRef]
- Wisniewski, M.; Nassuth, A.; Arora, R. Cold Hardiness in Trees: A Mini-Review. Front. Plant Sci. 2018, 9, 1394. [Google Scholar] [CrossRef]
- Guerra, D.; Lamontanara, A.; Bagnaresi, P.; Orrù, L.; Rizza, F.; Zelasco, S.; Beghè, D.; Ganino, T.; Pagani, D.; Cattivelli, L. Transcriptome changes associated with cold acclimation in leaves of olive tree (Olea europaea L.). Tree Genet. Genomes 2015, 11, 113. [Google Scholar] [CrossRef]
- Sakai, A.; Larcher, W. Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Springer Science & Business Media: Berlin, Heidelberg, 2012; Volume 62. [Google Scholar]
- Gusta, L.V.; Wisniewski, M. Understanding plant cold hardiness: An opinion. Physiol. Plant. 2013, 147, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, E.; Wisniewski, M. Response of fruit tree tissues to freezing temperatures. HortScience 1991, 26, 501–504. [Google Scholar] [CrossRef]
- Rubio-Valdés, G.; Cabello, D.; Rapoport, H.F.; Rallo, L. Olive Bud Dormancy Release Dynamics and Validation of Using Cuttings to Determine Chilling Requirement. Plants 2022, 11, 3461. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Form | Model | Name |
|---|---|---|---|
| EXP0 | M1 | Exponential model without LAG | |
| EXPLAG | M2 | Exponential model with LAG | |
| L2p | M3 | 2p-logistic model | |
| GOM | M4 | Gompertz model | |
| LOG | M5 | Logistic model | |
| GOM2 | M6 | Gompertz model 2 | |
| LOG2 | M7 | Logistic model 2 | |
| TPLOG | M8 | Two-pool logistic | |
| OPLOG | M9 | One-pool logistic | |
| MGOM | M10 | Modified Gompertz model | |
| RCD | M11 | Richard model | |
| ELM | . | M12 | Exponential-linear model |
| OPG | . | M13 | One pool Gompertz |
| RCD2 | M14 | Richard model 2 | |
| RCD3 | M15 | Richard model 3 | |
| RCD4 | M16 | Richard model 4 | |
| GOM3 | M17 | Gompertz model 3 | |
| GOM4 | M18 | Gompertz model 4 |
| CV1 | CV2 | CV3 | CV4 | CV5 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | |
| M2 | 7.53 | 5.14 | 5.01 | 8.47 | 4.52 | 5.22 | 8.68 | 5.36 | 6.89 | 7.2 | 4.89 | 5.03 | 7.23 | 5.22 | 7.07 |
| M3 | 8.5 | 4.37 | 3.49 | 9.06 | 4.84 | 6.03 | 4.89 | 3.96 | 5.39 | 8.04 | 3.5 | 5.45 | 7.95 | 6.77 | 8.44 |
| M4 | 7.79 | 4.62 | 3.95 | 8.48 | 4.56 | 5.49 | 6.34 | 4.47 | 5.87 | 7.39 | 3.75 | 4.93 | 7.37 | 5.94 | 7.72 |
| M5 | 8.5 | 4.37 | 3.49 | 9.06 | 4.84 | 6.03 | 4.89 | 3.96 | 5.39 | 8.04 | 3.5 | 5.45 | 7.95 | 6.77 | 8.44 |
| M6 | 7.79 | 4.62 | 3.95 | 8.48 | 4.56 | 5.49 | 6.34 | 4.47 | 5.87 | 7.39 | 3.75 | 4.93 | 7.37 | 5.94 | 7.72 |
| M8 | 8.46 | 4.38 | 3.3 | 9.04 | 4.84 | 6.03 | 3.6 | 3.64 | 5.21 | 8.02 | 3.48 | 5.45 | 7.88 | 6.78 | 8.45 |
| M10 | 7.79 | 4.62 | 3.95 | 8.48 | 4.56 | 5.49 | 6.34 | 4.47 | 5.87 | 7.39 | 3.75 | 4.93 | 7.37 | 5.94 | 7.72 |
| M14 | 7.79 | 4.3 | 3.4 | 8.45 | 4.57 | 5.49 | 3.82 | 3.76 | 5.41 | 7.31 | 3.5 | 4.93 | 7.22 | 5.95 | 7.72 |
| CV6 | CV7 | CV8 | CV9 | CV10 | |||||||||||
| Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | |
| M2 | 7.83 | 3.29 | 6.18 | 6.91 | 7.24 | 6.46 | 6.98 | 6 | 5.69 | 7.51 | 5.52 | 6.4 | 8.61 | 4.22 | 7.48 |
| M3 | 8.51 | 4.66 | 7.51 | 5.07 | 5.56 | 4.74 | 8.06 | 7.54 | 6.92 | 8.42 | 6.93 | 7.68 | 5.59 | 4.11 | 6.11 |
| M4 | 7.91 | 3.87 | 6.74 | 5.5 | 6.17 | 5.32 | 7.33 | 6.8 | 6.28 | 7.77 | 6.18 | 6.95 | 6.73 | 4.06 | 6.57 |
| M5 | 8.51 | 4.66 | 7.51 | 5.07 | 5.56 | 4.74 | 8.06 | 7.54 | 6.92 | 8.42 | 6.93 | 7.68 | 5.59 | 4.11 | 6.11 |
| M6 | 7.91 | 3.87 | 6.74 | 5.5 | 6.17 | 5.32 | 7.33 | 6.8 | 6.28 | 7.77 | 6.18 | 6.95 | 6.73 | 4.06 | 6.57 |
| M8 | 8.47 | 4.66 | 7.51 | 4.81 | 5.21 | 4.24 | 8.06 | 7.57 | 6.95 | 8.38 | 6.94 | 7.69 | 4.29 | 4.11 | 5.65 |
| M10 | 7.91 | 3.87 | 6.74 | 5.5 | 6.17 | 5.32 | 7.33 | 6.8 | 6.28 | 7.77 | 6.18 | 6.95 | 6.73 | 4.06 | 6.57 |
| M14 | 7.86 | 3.86 | 6.74 | 5.08 | 5.27 | 4.49 | 7.32 | 6.81 | 6.28 | 7.69 | 6.18 | 6.95 | 5.05 | 4.05 | 5.82 |
| CV1 | CV2 | ||||||
|---|---|---|---|---|---|---|---|
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.47 ** ± 0.41 | 3.62 ** ± 0.31 | 3.98 ** ± 0.28 | 2.63 ** ± 0.41 | 3.07 ** ± 0.28 | 2.67 ** ± 0.29 | |
| 0.18 ** ± 0.02 | 0.09 ** ± 0.01 | 0.11 ** ± 0.01 | 0.18 ** ± 0.02 | 0.10 ** ± 0.01 | 0.11 ** ± 0.01 | ||
| 0.87, 0.86 | 0.92, 0.92 | 0.96, 0.96 | 0.90, 0.90 | 0.92, 0.91 | 0.89, 0.89 | ||
| CV3 | CV4 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 5.60 ** ± 0.63 | 4.48 ** ± 0.30 | 4.51 ** ± 0.47 | 2.43 ** ± 0.38 | 3.81 ** ± 0.27 | 3.14 ** ± 0.33 | |
| 0.16 ** ± 0.01 | 0.10 ** ± 0.00 | 0.10 ** ± 0.01 | 0.18 ** ± 0.02 | 0.12 ** ± 0.01 | 0.12 ** ± 0.01 | ||
| 0.96, 0.96 | 0.97, 0.97 | 0.92, 0.91 | 0.88, 0.88 | 0.97, 0.97 | 0.92, 0.91 | ||
| CV5 | CV6 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.36 ** ± 0.36 | 2.65 ** ± 0.33 | 2.26 ** ± 0.34 | 2.50 ** ± 0.36 | 2.54 ** ± 0.20 | 2.32 ** ± 0.32 | |
| 0.18 ** ± 0.02 | 0.12 ** ± 0.01 | 0.12 ** ± 0.01 | 0.18 ** ± 0.02 | 0.14 ** ± 0.01 | 0.14 ** ± 0.01 | ||
| 0.88, 0.88 | 0.88, 0.87 | 0.81, 0.80 | 0.91, 0.91 | 0.96, 0.96 | 0.87, 0.86 | ||
| CV7 | CV8 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 3.50 ** ± 0.30 | 4.65 ** ± 0.56 | 4.38 ** ± 0.44 | 2.42 ** ± 0.38 | 2.48 ** ± 0.33 | 2.37 ** ± 0.29 | |
| 0.14 ** ± 0.01 | 0.11 ** ± 0.01 | 0.11 ** ± 0.01 | 0.18 ** ± 0.02 | 0.10 ** ± 0.01 | 0.10 ** ± 0.01 | ||
| 0.96, 0.96 | 0.90, 0.90 | 0.93, 0.92 | 0.88, 0.87 | 0.83, 0.82 | 0.85, 0.84 | ||
| CV9 | CV10 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.57 ** ± 0.36 | 2.64 ** ± 0.33 | 2.32 ** ± 0.32 | 4.62 ** ± 0.56 | 3.67 ** ± 0.30 | 4.70 ** ± 0.55 | |
| 0.18 ** ± 0.02 | 0.11 ** ± 0.00 | 0.13 ** ± 0.01 | 0.15 ** ± 0.01 | 0.08 ** ± 0.01 | 0.10 ** ± 0.01 | ||
| 0.92, 0.92 | 0.87, 0.86 | 0.86, 0.85 | 0.94, 0.93 | 0.90, 0.90 | 0.87, 0.86 | ||
| CV1 | CV2 | CV3 | CV4 | CV5 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | |
| M3 | 9.24 | 5.78 | 4.97 | 8.63 | 6.2 | 7.32 | 8.12 | 5.26 | 7.15 | 8.09 | 6.22 | 6.98 | 8.21 | 6.68 | 6.34 |
| M4 | 8.8 | 6.21 | 5.65 | 7.84 | 5.16 | 6.27 | 8.34 | 6.22 | 7.86 | 7.22 | 5.2 | 5.93 | 7.33 | 5.72 | 5.6 |
| M5 | 9.24 | 5.78 | 4.97 | 8.63 | 6.2 | 7.32 | 8.12 | 5.26 | 7.15 | 8.09 | 6.22 | 6.98 | 8.21 | 6.68 | 6.34 |
| M6 | 8.8 | 6.21 | 5.65 | 7.84 | 5.16 | 6.27 | 8.34 | 6.22 | 7.86 | 7.22 | 5.2 | 5.93 | 7.33 | 5.72 | 5.6 |
| M7 | 9.24 | 5.78 | 4.97 | 8.63 | 6.2 | 7.32 | 8.12 | 5.26 | 7.15 | 8.09 | 6.22 | 6.98 | 8.21 | 6.68 | 6.34 |
| M10 | 8.8 | 6.21 | 5.65 | 7.84 | 5.16 | 6.27 | 8.34 | 6.22 | 7.86 | 7.22 | 5.2 | 5.93 | 7.33 | 5.72 | 5.6 |
| M13 | 8.8 | 6.21 | 5.65 | 7.84 | 5.16 | 6.27 | 8.34 | 6.22 | 7.86 | 7.22 | 5.2 | 5.93 | 7.33 | 5.72 | 5.6 |
| M14 | 8.8 | 5.75 | 4.95 | 7.83 | 5.16 | 6.21 | 8.09 | 5.15 | 7.13 | 7.22 | 5.2 | 5.93 | 7.27 | 5.73 | 5.57 |
| CV6 | CV7 | CV8 | CV9 | CV10 | |||||||||||
| Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | Pre | Deep | Post | |
| M3 | 8.52 | 6.47 | 4.6 | 8.22 | 5.4 | 4 | 8.18 | 6.68 | 13.2 | 8.12 | 6.59 | 7.49 | 8.73 | 5.84 | 10.8 |
| M4 | 7.69 | 5.35 | 4.53 | 8.52 | 6.67 | 4.83 | 7.23 | 5.69 | 15.33 | 7.11 | 5.53 | 7.45 | 8.93 | 7.03 | 10.76 |
| M5 | 8.52 | 6.47 | 4.6 | 8.22 | 5.4 | 4 | 8.18 | 6.68 | 13.2 | 8.12 | 6.59 | 7.49 | 8.73 | 5.84 | 10.8 |
| M6 | 7.69 | 5.35 | 4.53 | 8.52 | 6.55 | 4.83 | 7.23 | 5.69 | 15.33 | 7.11 | 5.53 | 7.45 | 8.93 | 7.03 | 10.76 |
| M7 | 8.52 | 6.47 | 4.6 | 8.22 | 5.4 | 4 | 8.18 | 6.68 | 13.2 | 8.12 | 6.59 | 7.49 | 8.73 | 5.84 | 10.8 |
| M10 | 7.69 | 5.35 | 4.53 | 8.52 | 6.55 | 4.83 | 7.23 | 5.69 | 15.33 | 7.11 | 5.53 | 7.45 | 8.93 | 7.03 | 10.76 |
| M13 | 7.69 | 5.35 | 4.53 | 8.52 | 6.55 | 4.83 | 7.23 | 5.69 | 15.33 | 7.11 | 5.53 | 7.45 | 8.93 | 7.03 | 10.76 |
| M14 | 7.55 | 5.35 | 4.38 | 8.2 | 5.4 | 3.99 | 7.23 | 5.69 | 11.33 | 7.11 | 5.53 | 7.11 | 8.7 | 5.44 | 10.8 |
| CV1 | CV2 | ||||||
|---|---|---|---|---|---|---|---|
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.58 ** ± 0.37 | 8.24 ** ± 1.59 | 6.39 ** ± 1.00 | 2.68 ** ± 0.36 | 3.79 ** ± 0.40 | 3.59 ** ± 0.55 | |
| 0.17 ** ± 0.02 | 0.20 ** ± 0.02 | 0.20 ** ± 0.01 | 0.19 ** ± 0.02 | 0.18 ** ± 0.01 | 0.22 ** ± 0.02 | ||
| 0.93, 0.92 | 0.97, 0.97 | 0.97, 0.97 | 0.94, 0.94 | 0.98, 0.98 | 0.97, 0.97 | ||
| CV3 | CV4 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.98 ** ± 0.42 | 9.81 ** ± 2.09 | 10.51 ** ± 2.89 | 2.63 ** ± 0.32 | 3.80 ** ± 0.40 | 3.81 ** ± 0.48 | |
| 0.14 ** ± 0.01 | 0.19 ** ± 0.02 | 0.21 ** ± 0.02 | 0.18 ** ± 0.02 | 0.17 ** ± 0.01 | 0.20 ** ± 0.01 | ||
| 0.93, 0.92 | 0.96, 0.96 | 0.95, 0.94 | 0.95, 0.95 | 0.98, 0.97 | 0.97, 0.97 | ||
| CV5 | CV6 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.80 ** ± 0.34 | 3.58 ** ± 0.40 | 3.84 ** ± 0.47 | 2.67 ** ± 0.36 | 3.34 ** ± 0.34 | 3.98 ** ± 0.40 | |
| 0.21 ** ± 0.02 | 0.17 ** ± 0.01 | 0.21 ** ± 0.01 | 0.20 ** ± 0.02 | 0.18 ** ± 0.01 | 0.22 ** ± 0.01 | ||
| 0.96, 0.96 | 0.97, 0.97 | 0.98, 0.97 | 0.95, 0.95 | 0.97, 0.97 | 0.98, 0.98 | ||
| CV7 | CV8 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 3.38 ** ± 0.55 | 9.94 ** ± 2.03 | 5.87 ** ± 0.47 | 2.69 ** ± 0.34 | 3.66 ** ± 0.41 | 7.24 * ± 2.81 | |
| 0.18 ** ± 0.02 | 0.20 ** ± 0.02 | 0.19 ** ± 0.01 | 0.19 ** ± 0.02 | 0.17 ** ± 0.01 | 0.22 ** ± 0.04 | ||
| 0.94, 0.93 | 0.97, 0.97 | 0.99, 0.99 | 0.95, 0.95 | 0.97, 0.97 | 0.88, 0.88 | ||
| CV9 | CV10 | ||||||
| Pre | Deep | Post | Pre | Deep | Post | ||
| Cofe. | 2.69 ** ± 0.34 | 3.44 ** ± 0.37 | 4.38 ** ± 0.62 | 2.92 ** ± 0.43 | 9.98 ** ± 2.17 | 16.07 * ± 6.00 | |
| 0.19 ** ± 0.02 | 0.17 ** ± 0.01 | 0.23 ** ± 0.02 | 0.14 ** ± 0.01 | 0.18 ** ± 0.02 | 0.25 ** ± 0.03 | ||
| 0.95, 0.95 | 0.97, 0.97 | 0.98, 0.98 | 0.91, 0.91 | 0.96, 0.96 | 0.97, 0.96 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rezaei, M.; Rohani, A. Estimating Freezing Injury on Olive Trees: A Comparative Study of Computing Models Based on Electrolyte Leakage and Tetrazolium Tests. Agriculture 2023, 13, 1137. https://doi.org/10.3390/agriculture13061137
Rezaei M, Rohani A. Estimating Freezing Injury on Olive Trees: A Comparative Study of Computing Models Based on Electrolyte Leakage and Tetrazolium Tests. Agriculture. 2023; 13(6):1137. https://doi.org/10.3390/agriculture13061137
Chicago/Turabian StyleRezaei, Mehdi, and Abbas Rohani. 2023. "Estimating Freezing Injury on Olive Trees: A Comparative Study of Computing Models Based on Electrolyte Leakage and Tetrazolium Tests" Agriculture 13, no. 6: 1137. https://doi.org/10.3390/agriculture13061137