
Effect of Dietary Salicin Standardized Extract from Salix alba Bark on Oxidative Stress Biomarkers and Intestinal Microflora of Broiler Chickens Exposed to Heat Stress
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Birds, Diets, and Experimental Design
2.2. Sample Collection
2.3. Chemical Analysis
2.4. Oxidative Stress Biomarkers in Liver
2.5. Intestinal Microbial Population Measurements
2.6. Statistical Analysis
3. Results
3.1. Effect of Salicin Standardized Extract from SAB on In Vitro Induced Lipid Peroxidation
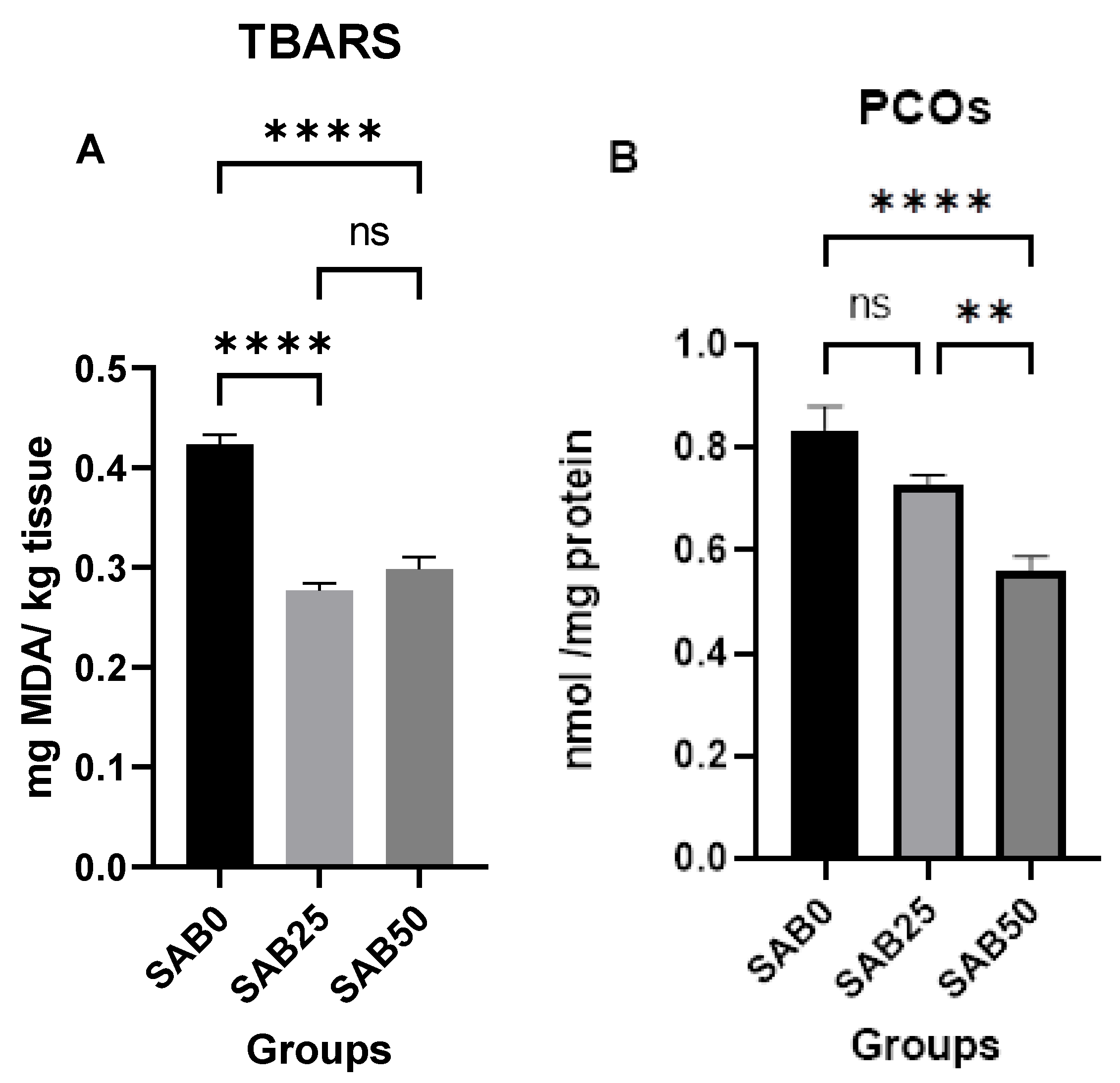
3.2. Oxidative Stress Biomarkers
3.3. Intestinal Microbial Population Measurements
3.4. Correlation between Antioxidant Capacity, Lipid and Protein Oxidation Biomarkers, and Intestinal Microflora
4. Discussion
4.1. Effect of Salicin Standardized Extract from Salix Alba Bark on In Vitro-Induced Lipid Peroxidation
4.2. Oxidative Stress Biomarkers
4.3. Intestinal Microbial Population Measurements
4.4. Correlation between Liver Oxidative Biomarkers and Intestinal Microflora
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Li, N.; Liu, J.; Zhang, Z.; Liu, Y.; Huang, C. Changes in global and regional characteristics of heat stress waves in the 21st century. Earth’s Future 2020, 8, e2020EF001636. [Google Scholar] [CrossRef]
- Perini, F.; Cendron, F.; Rovelli, G.; Castellini, C.; Cassandro, M.; Lasagna, E. Emerging genetic tools to investigate molecular pathways related to heat stress in chickens: A review. Animals 2020, 11, 46. [Google Scholar] [CrossRef]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.H.; Cheng, K.; Zheng, X.C.; Ahmad, H.; Zhang, L.L.; Wang, T. Effects of dietary supplementation with enzymatically treated Artemisia annua on growth performance, intestinal morphology, digestive enzyme activities, immunity, and antioxidant capacity of heat-stressed broilers. Poult. Sci. 2018, 97, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Emami, N.K.; Jung, U.; Voy, B.; Dridi, S. Radical response: Effects of heat stress-induced oxidative stress on lipid metabolism in the avian liver. Antioxidants 2020, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Calik, A.; Emami, N.K.; Schyns, G.; White, M.B.; Walsh, M.C.; Romero, L.F.; Dalloul, R.A. Influence of dietary vitamin E and selenium supplementation on broilers subjected to heat stress, Part II: Oxidative stress, immune response, gut integrity, and intestinal microbiota. Poult Sci. 2022, 101, 101858. [Google Scholar] [CrossRef]
- Valentin-Cortez, F.J.; Córdova-Gallardo, J.; Méndez-Sánchez, N. Narrative review of gut microbiota and liver diseases: Facts and fictions. Dig. Med. Res. 2022, 5, 16. [Google Scholar] [CrossRef]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Pečjak, M.; Leskovec, J.; Levart, A.; Salobir, J.; Rezar, V. Effects of dietary vitamin E, vitamin C, selenium and their combination on carcass characteristics, oxidative stability and breast meat quality of broiler chickens exposed to cyclic heat stress. Animals 2022, 12, 1789. [Google Scholar] [CrossRef]
- Untea, A.E.; Panaite, T.D.; Dragomir, C.; Ropota, M.; Olteanu, M.; Varzaru, I. Effect of dietary chromium supplementation on meat nutritional quality and antioxidant status from broilers fed with Camelina-meal-supplemented diets. Animal 2019, 13, 2939–2947. [Google Scholar] [CrossRef]
- Vlaicu, P.A.; Panaite, T.D.; Olteanu, M.; Turcu, R.P.; Saracila, M.; Criste, R.D. Effect of the dietary oregano (Origanum vulgare L.) powder and oil on the performance, carcass and organs development of broilers reared under heat stress (32 C). Lucr. Științifice-Univ. Științe Agric. Med. Vet. Ser. Zooteh. 2018, 69, 207–213. [Google Scholar]
- Saracila, M.; Panaite, T.D.; Tabuc, C.; Soica, C.; Untea, A.; Varzaru, I.; Criste, R.D. Maintaining intestinal microflora balance in heat-stressed broilers using dietary creeping wood sorrel (Oxalis corniculata) powder and chromium (chromium picolinate). Span. J. Agric. Res. 2020, 18, e0612. [Google Scholar] [CrossRef]
- Salah, A.S.; Ahmed-Farid, O.A.; Nassan, M.A.; El-Tarabany, M.S. Dietary curcumin improves energy metabolism, brain monoamines, carcass traits, muscle oxidative stability and fatty acid profile in heat-stressed broiler chickens. Antioxidants 2021, 10, 1265. [Google Scholar] [CrossRef]
- Adamiak, K.; Lewandowska, K.; Sionkowska, A. The influence of salicin on rheological and film-forming properties of collagen. Molecules 2021, 26, 1661. [Google Scholar] [CrossRef] [PubMed]
- Saracila, M.; Panaite, T.D.; Vlaicu, P.A.; Tabuc, C.; Palade, M.L.; Gavris, T.; Criste, R.D. Dietary Willow Bark Extract for Broilers Reared Under Heat Stress. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Anim. Sci. Biotechnol. 2018, 75, 92–98. [Google Scholar] [CrossRef]
- Al-Fataftah, A.R.; Abdelqader, A. Effect of Salix babylonica, Populus nigra and Eucalyptus camaldulensis ex-tracts in drinking water on performance and heat tolerance of broiler chickens during heat stress. Am. Eurasian J. Agric. Environ. Sci. 2013, 13, 1309–1313. [Google Scholar]
- Panaite, T.D.; Saracila, M.; Papuc, C.P.; Predescu, C.N.; Soica, C. Influence of dietary supplementation of Salix alba bark on performance, oxidative stress parameters in liver and gut microflora of broilers. Animals 2020, 10, 958. [Google Scholar] [CrossRef]
- Varzaru, I.; Untea, A.E.; Saracila, M. In vitro antioxidant properties of berry leaves and their inhibitory effect on lipid peroxidation of thigh meat from broiler chickens. Eur. J. Lipid Sci. Technol. 2020, 122, 1900384. [Google Scholar] [CrossRef]
- Untea, A.E.; Varzaru, I.; Vlaicu, P.A.; Turcu, R.P.; Panaite, T.D. Studies on antioxidant activities of grape pomace using in vitro, ex vivo, and in vivo models. J. Food Meas. Charact. 2022, 17, 121–128. [Google Scholar] [CrossRef]
- Janaszewska, A.; Bartosz, G. Assay of total antioxidant capacity: Comparison of four methods as applied to human blood plasma. Scand. J. Clin. Lab. Investig. 2002, 62, 231–236. [Google Scholar] [CrossRef]
- Reddy, Y.N.; Murthy, S.V.; Krishna, D.R.; Prabhakar, M.C. Role of free radicals and antioxidants in tuberculosis patients. Indian J. Tuberc. 2004, 51, 213–218. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid Peroxidation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; Volume 186, pp. 421–431. [Google Scholar]
- Patsoukis, N.; Zervoudakis, G.; Panagopoulos, N.T.; Georgiou, C.D.; Angelatou, F.; Matsokis, N.A. Thiol redox state (TRS) and oxidative stress in the mouse hippocampus after pentylenetetrazol-induced epileptic seizure. Neurosci. Lett. 2004, 357, 83–86. [Google Scholar] [CrossRef]
- Criste, R.D.; Panaite, T.D.; Tabuc, C.; Saracila, M.; Soica, C.; Olteanu, M. Effect of oregano and rosehip supplements on broiler (14–35 days) performance, carcass and internal organs development and gut health. AgroLife Sci. J. 2017, 6, 75–83. [Google Scholar]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft: New York, NY, USA, 2023. [Google Scholar]
- Bonaterra, G.A.; Heinrich, E.U.; Kelber, O.; Weiser, D.; Metz, J.; Kinscherf, R. Anti-inflammatory effects of the willow bark extract STW 33-I (Proaktiv®) in LPS-activated human monocytes and differentiated macrophages. Phytomedicine 2010, 17, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Bonaterra, G.A.; Kelber, O.; Weiser, D.; Metz, J.; Kinscherf, R. In vitro anti-proliferative effects of the willow bark extract STW 33-I. Arzneimittelforschung 2010, 60, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Durak, A.; Gawlik-Dziki, U. The study of interactions between active compounds of coffee and willow (Salix sp.) bark water extract. BioMed Res. Int. 2014, 2014, e386953. [Google Scholar] [CrossRef] [Green Version]
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837. [Google Scholar] [CrossRef] [Green Version]
- Basiouni, S.; Tellez-Isaias, G.; Latorre, J.D.; Graham, B.D.; Petrone-Garcia, V.M.; El-Seedi, H.R.; Yalçın, S.; El-Wahab, A.A.; Visscher, C.; May-Simera, H.L.; et al. Anti-inflammatory and antioxidative phytogenic substances against secret killers in poultry: Current status and prospects. Vet. Sci. 2023, 10, 55. [Google Scholar] [CrossRef]
- Khayyal, M.T.; El-Ghazaly, M.A.; Abdallah, D.M.; Okpanyi, S.N.; Kelber, O.; Weiser, D. Mechanisms involved in the anti-inflammatory effect of a standardized willow bark extract. Arzneimittelforschung 2005, 55, 677–687. [Google Scholar] [CrossRef]
- Zhai, K.; Hong, D.; Khan, J.G.; Hui, X.; Han, F.; Cao, W.; Gao, G.; Shan, L.; Wei, Z. Salicin from Alangium chinense ameliorates rheumatoid arthritis by modulating the Nrf2-HO-1-ROS pathways. J. Agric. Food Chem. 2018, 66, 6073–6082. [Google Scholar] [CrossRef]
- Moskaug, J.Ø.; Carlsen, H.; Myhrstad, M.C.W.; Blomhoff, R. Polyphenols and glutathione synthesis regulation. Am. J. Clin. Nutr. 2005, 81, 277S–283S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Bai, K.; He, J.; Niu, Y.; Lu, Y.; Zhang, L.; Wang, T. Curcumin attenuates hepatic mitochondrial dysfunction through the maintenance of thiol pool, inhibition of mtDNA damage, and stimulation of the mi-tochondrial thioredoxin system in heat-stressed broilers. J. Anim. Sci. 2018, 96, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; He, Y.; Arowolo, M.A.; Wu, S.; He, J. Polyphenols as Potential Attenuators of Heat Stress in Poultry Production. Antioxidants 2019, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Scaloni, A.; Giustarini, D.; Cavarra, E.; Tell, G.; Lungarella, G. Proteins as biological markers of oxidative/nitrosative stress in diseases. The contribution of redox-proteomics. Mass Spectrom. Rev. 2005, 24, 55–99. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Ishikado, A.; Sono, Y.; Matsumoto, M.; Robida-Stubbs, S.; Okunoa, A.; Goto, M.; King, G.L.; Blackwell, T.K.; Makino, T. Willow bark extract increases antioxidant enzymes and reduces oxidative stress through activation of Nrf2 in vascular endothelial cells and Caenorhabditis elegans. Free Radic. Biol. Med. 2013, 65, 1506–1515. [Google Scholar] [CrossRef] [Green Version]
- Saracila, M.; Panaite, T.D.; Papuc, C.P.; Criste, R.D. Heat stress in broiler chickens and the effect of dietary polyphenols, with special reference to willow (Salix spp.) bark supplements—A Review. Antioxidants 2021, 10, 686. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, D.I.; Tawfeek, N.; El-Shiekh, R.A.; Khalil, H.; Mahmoud, M.Y.; Bakr, A.F.; El-Shazly, A.M. Salix subserrata bark extract-loaded chitosan nanoparticles attenuate neurotoxicity induced by sodium arsenate in rats in relation with HPLC–PDA-ESI–MS/MS Profile. AAPS PharmSciTech 2023, 24, 15. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, S. Salicin inhibits AGE-induced degradation of type II collagen and aggrecan in hu-man SW1353 chondrocytes: Therapeutic potential in osteoarthritis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1043–1049. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Li, Y.; Yu, Z.; Yang, L.; Shang, R.; Yan, Z. Protective effect of vitamin C on triptolide-induced acute hepatotoxicity in mice through mitigation of oxidative stress. An. Acad. Bras. Ciências 2019, 91, e20181257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Majdeddin, M.; Degroote, J.; Van Liefferinge, E.; Van Noten, N.; Van Kerschaver, C.; Michiels, J. Effect of supplemental methyl sulfonyl methane on performance, carcass and meat quality and oxidative status in chronic cyclic heat-stressed finishing broilers. Poult. Sci. 2023, 102, 102321. [Google Scholar] [CrossRef] [PubMed]
- Saracila, M.; Panaite, T.; Tabuc, C.; Soica, C.; Untea, A.; Ayasan, T.; Criste, R.D. Dietary ascorbic acid and chromium supplementation for broilers reared under thermoneutral conditions vs. high heat stress. Sci. Pap. Anim. Sci. Ser. Lucr. Ştiinţifice-Ser. Zooteh. 2020, 73, 41–47. [Google Scholar]
- Patra, A.K.; Kar, I. Heat stress on microbiota composition, barrier integrity, and nutrient transport in gut, production performance, and its amelioration in farm animals. J. Anim. Sci. Technol. 2021, 63, 211–247. [Google Scholar] [CrossRef]
- Ringseis, R.; Eder, K. Heat stress in pigs and broilers: Role of gut dysbiosis in the impairment of the gut-liver axis and restoration of these effects by probiotics, prebiotics and synbiotics. J. Anim. Sci. Biotechnol. 2022, 13, 126. [Google Scholar] [CrossRef]
- Swelum, A.A.; Elbestawy, A.R.; El-Saadony, M.T.; Hussein, E.O.S.; Alhotan, R.; Suliman, G.M.; Taha, A.E.; Ba-Awadh, H.; El-Tarabily, K.A.; Abd El-Hack, M.E. Ways to minimize bacterial infections, with special reference to Escherichia coli, to cope with the first-week mortality in chicks: An updated overview. Poult. Sci. 2021, 100, 101039. [Google Scholar] [CrossRef]
- Szafraniec, G.M.; Szeleszczuk, P.; Dolka, B. Review on skeletal disorders caused by Staphylococcus spp. in poultry. Vet. Q. 2022, 42, 21–40. [Google Scholar] [CrossRef]
- Schmid, B.; Kötter, I.; Heide, L. Pharmacokinetics of salicin after oral administration of a standardised willow bark ex-tract. Eur. J. Clin. Pharm. 2001, 57, 387–391. [Google Scholar] [CrossRef]
- Dou, J.; Ilina, P.; Hemming, J.; Malinen, K.; Mäkkylä, H.; Oliveira de Farias, N.; Tammela, P.; de Aragão Umbuzeiro, G.; Räisänen, R.; Vuorinen, T. Effect of hybrid type and harvesting season on phytochemistry and antibacterial activity of extracted metabolites from Salix bark. J. Agric. Food Chem. 2022, 70, 2948–2956. [Google Scholar] [CrossRef]
- Carpa, R.; Remizovschi, A.; Burtescu, R.F.; Culda, C.A.; Kryvtsova, M.; Hasynets, Y.; Butiuc-Keul, A.; Dobrotă, C.; Farkas, A.; Olah, N.K. salicin content from Salix alba L. and Salix purpurea L. extracts and its antibacterial effects. Contrib. Bot. 2022, 57, 133–142. [Google Scholar] [CrossRef]
- Verma, N.; Verma, R.; Kumari, R.; Ranjha, R.; Paul, J. Effect of salicin on gut inflammation and on selected groups of gut microbiota in dextran sodium sulfate induced mouse model of colitis. Inflamm. Res. 2014, 63, 161–169. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Bao, Q.; Di, S.; Zhao, Y.; Zhao, S.; Zhang, H.; Tong, X. The interaction between the gut microbiota and herbal medicines. Biomed. Pharmacother. 2019, 118, 109252. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, B.; Tang, L.; Zhou, Y.; Wang, Q.; Gong, L.; Ni, J.; Li, W. Probiotic Bacillus alleviates oxidative stress-induced liver injury by modulating gut-liver axis in a rat model. Antioxidants 2022, 11, 291. [Google Scholar] [CrossRef] [PubMed]
- Conde de la Rosa, L.; Goicoechea, L.; Torres, S.; Garcia-Ruiz, C.; Fernandez-Checa, J.C. Role of Oxidative Stress in Liver Disorders. Livers 2022, 2, 283–314. [Google Scholar] [CrossRef]
- Kong, Y.; Olejar, K.J.; On, S.L.W.; Chelikani, V. The potential of Lactobacillus spp. for modulating oxidative stress in the gastrointestinal tract. Antioxidants 2020, 9, 610. [Google Scholar] [CrossRef]
- Durdun, C.; Papuc, C.; Crivineanu, M.; Nicorescu, V. Antioxidant potential of Lycopodium clavatum and Cnicus benedictus hydroethanolic extracts on stressed mice. Sci. Work.-Univ. Agron. Sci. Vet. Med. Buchar. Ser. C Vet. Med. 2011, 57, 61–68. [Google Scholar]
- Mavrommatis, A.; Giamouri, E.; Myrtsi, E.D.; Evergetis, E.; Filippi, K.; Papapostolou, H.; Koulocheri, S.D.; Zoidis, E.; Pappas, A.C.; Koutinas, A.; et al. Antioxidant status of broiler chickens fed diets supplemented with vinification by-products: A valorization approach. Antioxidants 2021, 10, 1250. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Grower Phase (14–35 Days) | Finisher Phase (36–42 Days) | ||||
|---|---|---|---|---|---|---|
| SAB0 | SAB25 | SAB50 | SAB0 | SAB25 | SAB50 | |
| Corn | 62.00 | 61.97 | 61.95 | 60.50 | 60.47 | 60.45 |
| Soybean meal | 26.58 | 26.58 | 26.58 | 25.46 | 25.46 | 25.46 |
| Gluten | 4.00 | 4.00 | 4.00 | 6.00 | 6.00 | 6.00 |
| Oil | 2.50 | 2.50 | 2.50 | 3.75 | 3.75 | 3.75 |
| Salix alba bark extract (25% salicin) | - | 0.025 | 0.050 | - | 0.025 | 0.050 |
| Calcium carbonate | 1.40 | 1.40 | 1.40 | 1.33 | 1.33 | 1.33 |
| Monocalcium phosphate | 1.36 | 1.36 | 1.36 | 1.13 | 1.13 | 1.13 |
| Salt | 0.37 | 0.37 | 0.37 | 0.33 | 0.33 | 0.33 |
| Methionine | 0.26 | 0.26 | 0.26 | 0.25 | 0.25 | 0.25 |
| Lysine | 0.48 | 0.48 | 0.48 | 0.20 | 0.20 | 0.20 |
| Choline | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Premix | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Total ingredients | 100 | 100 | 100 | 100 | 100 | 100 |
| Chemical analysis | ||||||
| Total polyphenols, mg/g GAE | 1.70 | 1.92 | 2.27 | 1.69 | 1.81 | 2.24 |
| Item | TBARS (mg/kg) | % Inhibition of LPO |
|---|---|---|
| Meat with induced LPO | 1.58 a | - |
| Meat with induced LPO and SAB (1000 mg/kg) | 1.39 b | 11.60 b |
| Meat with induced LPO and vitamin E (500 μM) | 1.19 c | 24.21 a |
| SEM | 0.547 | 0.813 |
| p-value | <0.0001 | <0.0001 |
| Item | Groups | SEM | p-Value | ||
|---|---|---|---|---|---|
| SAB0 | SAB25 | SAB50 | |||
| TAC (mmol Trolox/L) | 0.86 a | 0.98 a | 1.12 b | 0.027 | 0.001 |
| CAT (U/mL) | 1848.60 | 1788.38 | 1596.81 | 0.438 | 0.4346 |
| GSH (µmol/g tissue) | 3.138 a | 3.66 ab | 4.05 b | 0.179 | 0.012 |
| SOD (U/g tissue) | 2085.8 | 2187.3 | 2208.7 | 0.137 | 0.8161 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saracila, M.; Panaite, T.D.; Predescu, N.C.; Untea, A.E.; Vlaicu, P.A. Effect of Dietary Salicin Standardized Extract from Salix alba Bark on Oxidative Stress Biomarkers and Intestinal Microflora of Broiler Chickens Exposed to Heat Stress. Agriculture 2023, 13, 698. https://doi.org/10.3390/agriculture13030698
Saracila M, Panaite TD, Predescu NC, Untea AE, Vlaicu PA. Effect of Dietary Salicin Standardized Extract from Salix alba Bark on Oxidative Stress Biomarkers and Intestinal Microflora of Broiler Chickens Exposed to Heat Stress. Agriculture. 2023; 13(3):698. https://doi.org/10.3390/agriculture13030698
Chicago/Turabian StyleSaracila, Mihaela, Tatiana Dumitra Panaite, Nicoleta Corina Predescu, Arabela Elena Untea, and Petru Alexandru Vlaicu. 2023. "Effect of Dietary Salicin Standardized Extract from Salix alba Bark on Oxidative Stress Biomarkers and Intestinal Microflora of Broiler Chickens Exposed to Heat Stress" Agriculture 13, no. 3: 698. https://doi.org/10.3390/agriculture13030698