
Co-Contamination of Food and Feed with Mycotoxin and Bacteria and Possible Implications for Health

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
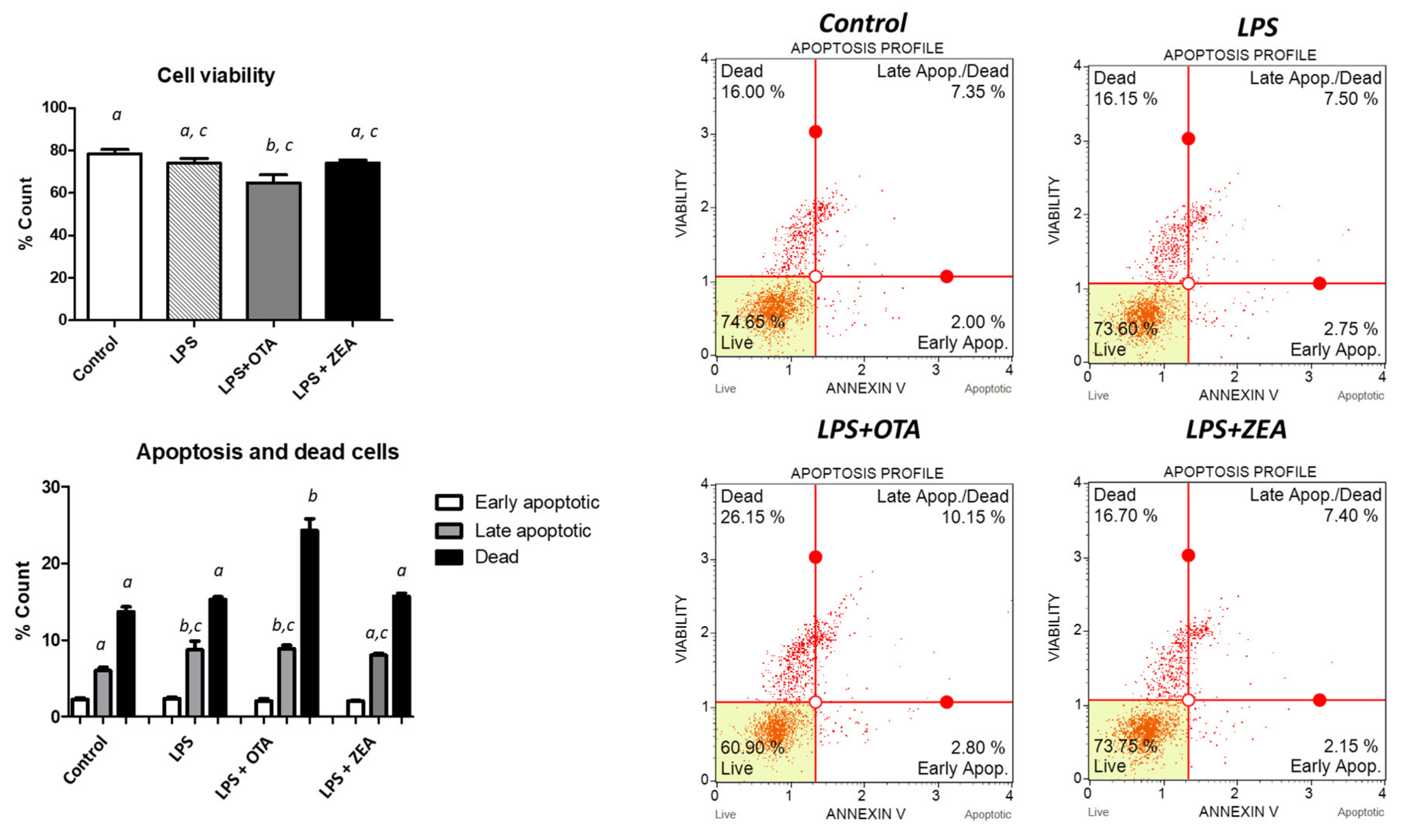
3.1. The Effect of Co-Exposure to LPS and Mycotoxins on Cell Proliferation, Apoptosis, and Necrosis
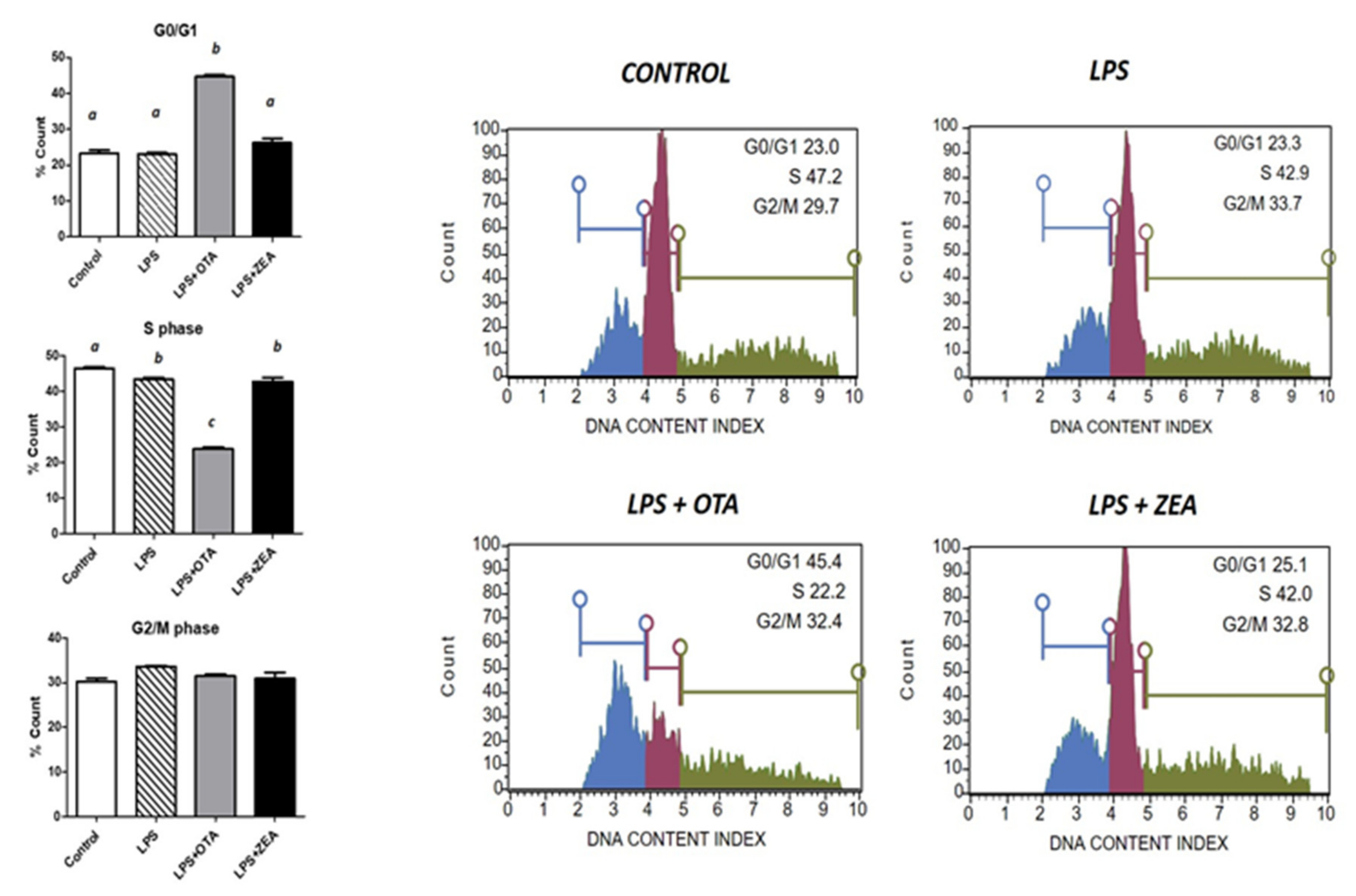
3.2. The Effect of Co-Exposure to LPS and Mycotoxins on Cell Cycle
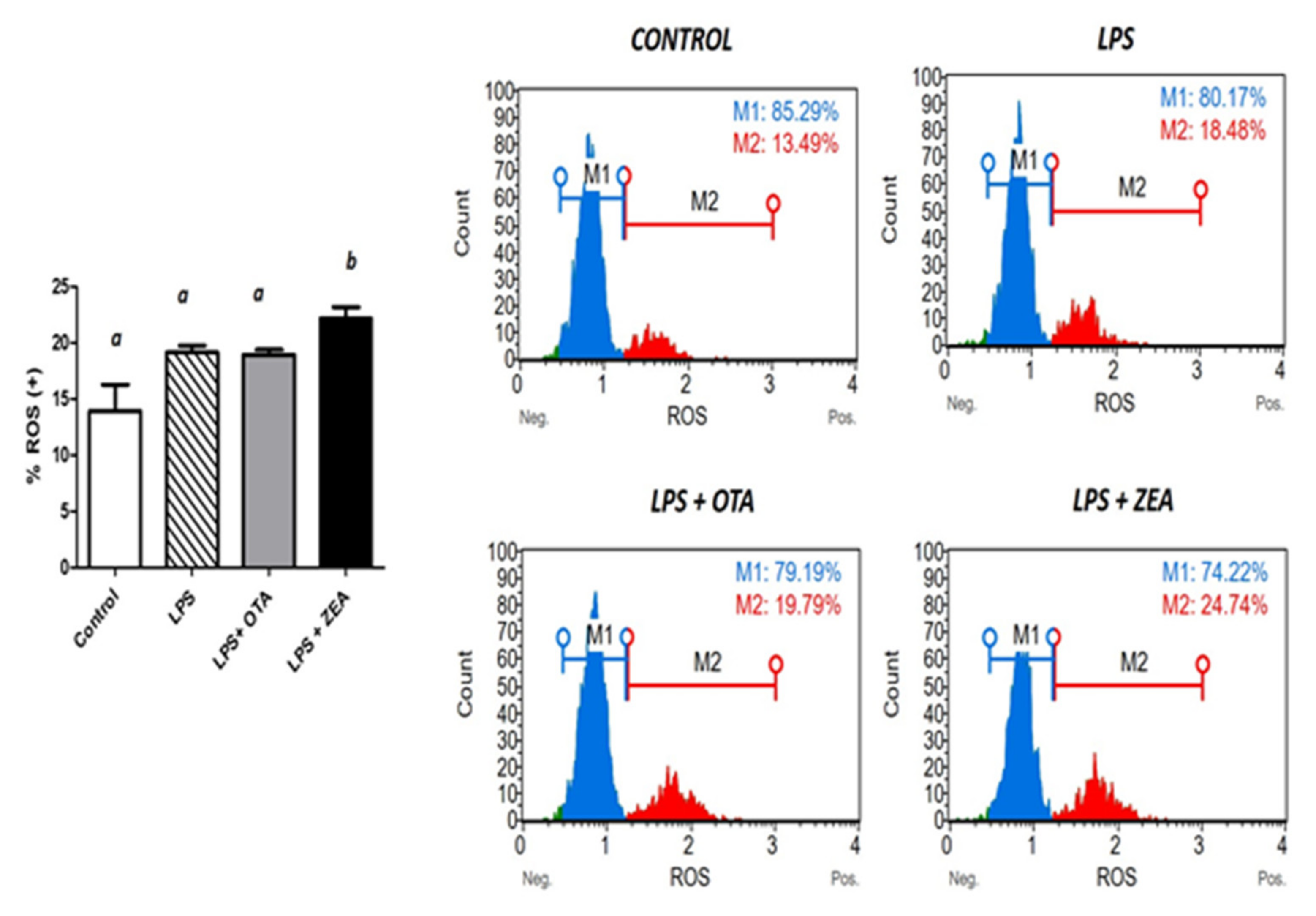
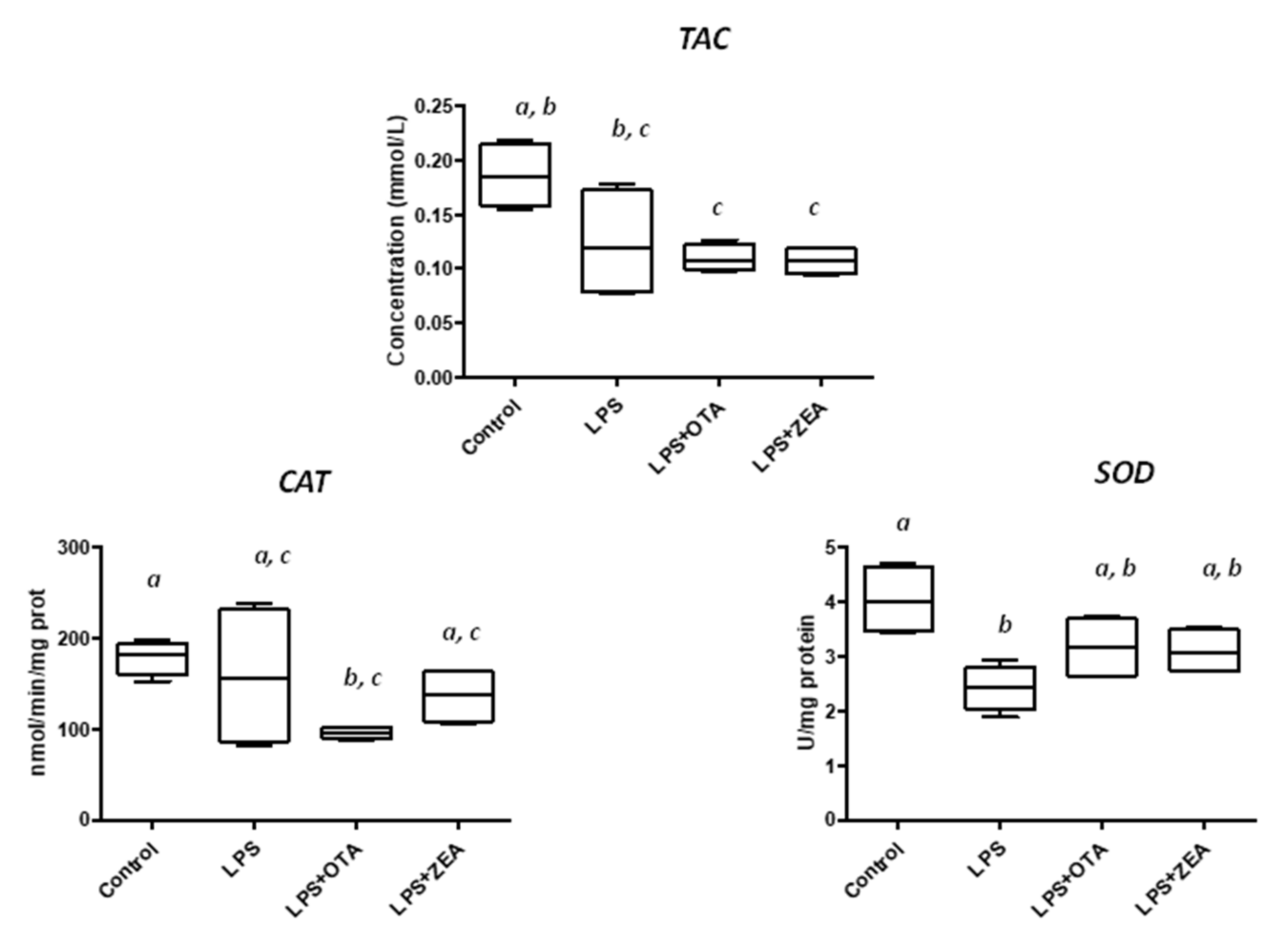
3.3. The Effect of Co-Exposure to LPS and Mycotoxins on ROS Production and on the Activity of the Enzymes Involved in Oxidative Defence
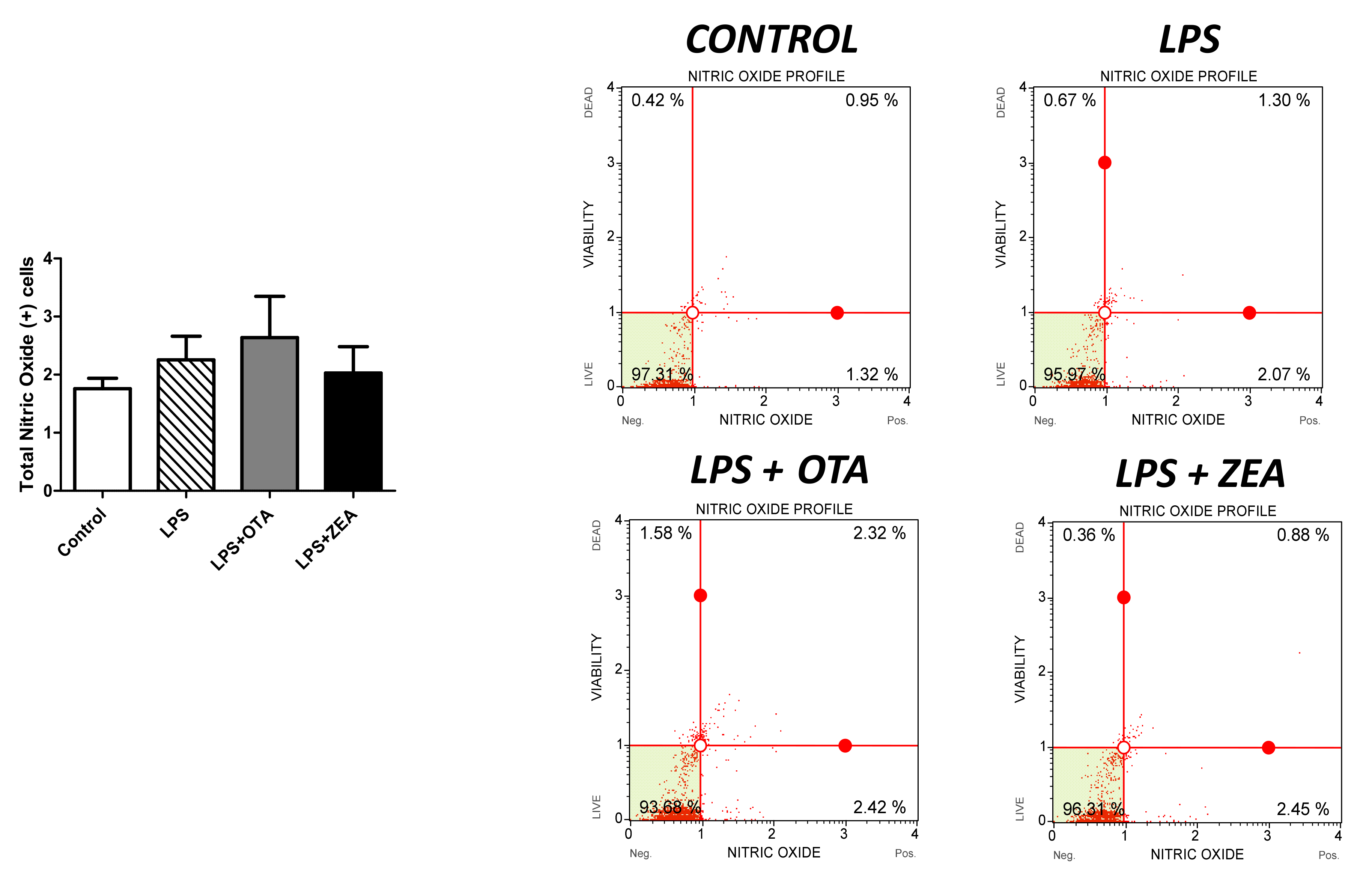
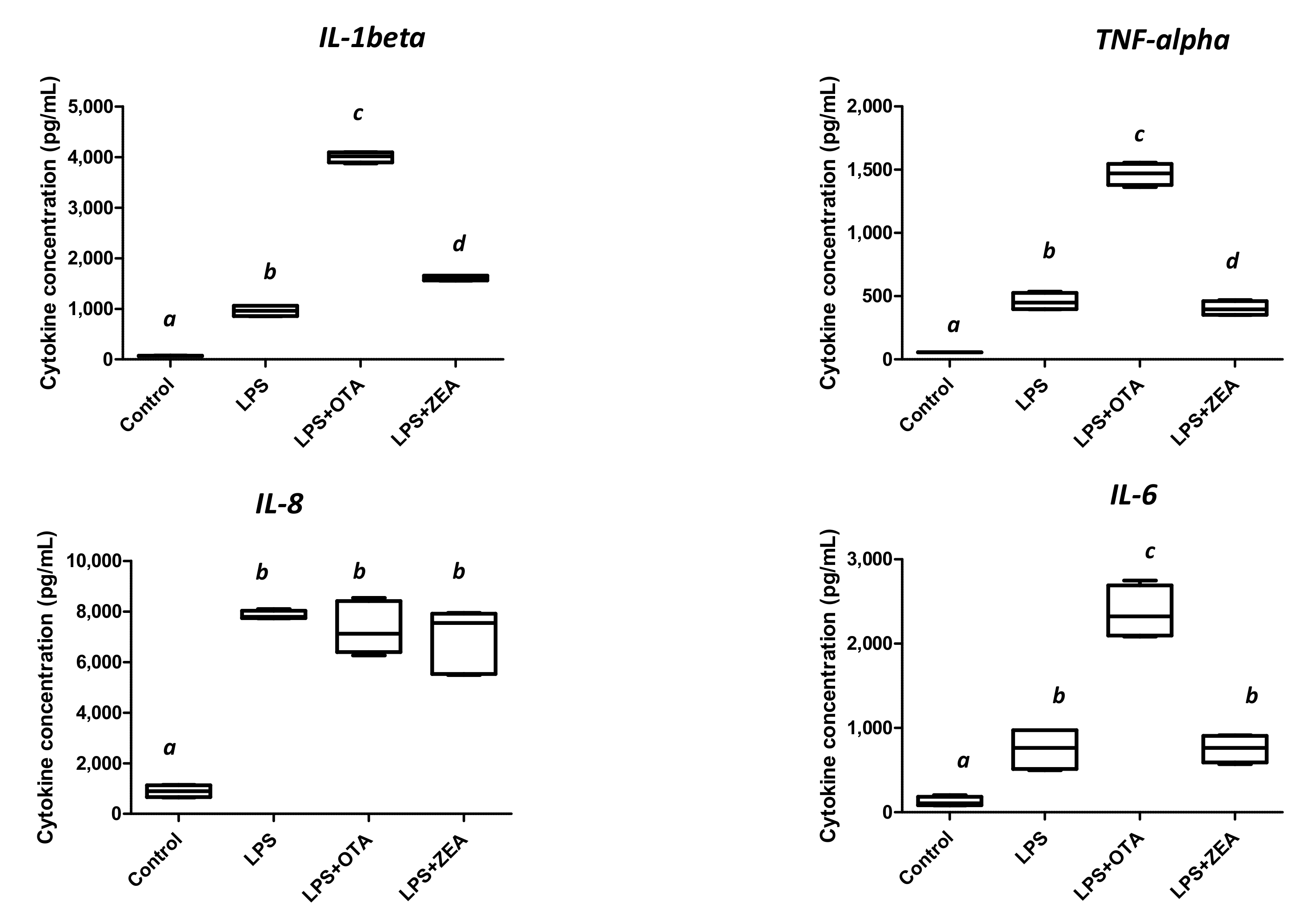
3.4. The Effect of Co-Exposure to LPS and Mycotoxins on Nitric Oxide and Inflammatory Cytokine Synthesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lorenzo, J.M.; Munekata, P.; Domínguez, R.; Pateiro, M.; Saraiva, J.; Franco Ruiz, D. Chapter 3—Main Groups of Microorganisms of Relevance for Food Safety and Stability: General Aspects and Overall Description. In Innovative Technologies for Food Preservation: Inactivation of Spoilage and Pathogenic Microorganisms; Academic Press: Cambridge, MA, USA, 2018; pp. 53–107. [Google Scholar]
- Alegbeleye, O.; Odeyemi, O.A.; Strateva, M.; Stratev, D. Microbial spoilage of vegetables, fruits and cereals. Appl. Food Res. 2022, 2, 100122. [Google Scholar] [CrossRef]
- Marroquín-Cardona, A.G.; Johnson, N.M.; Phillips, T.D.; Hayes, A.W. Mycotoxins in a changing global environment—A review. Food Chem. Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins affecting animals, foods, humans, and plants: Types, occurrence, toxicities, action mechanisms, prevention, and detoxification strategies—A revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [Green Version]
- Khoi, C.S.; Chen, J.H.; Lin, T.Y.; Chiang, C.K.; Hung, K.Y. Ochratoxin a-induced nephrotoxicity: Up-to-date evidence. Int. J. Mol. Sci. 2021, 22, 11237. [Google Scholar] [CrossRef]
- Gaffoor, I.; Trail, F. Characterization of two polyketide synthase genes involved in zearalenone biosynthesis in Gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1793–1799. [Google Scholar] [CrossRef] [Green Version]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and reproductive function in farm animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.E.; Taranu, I. Ochratoxin a and its effects on immunity. Toxin Rev. 2015, 34, 11–20. [Google Scholar] [CrossRef]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the immune response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- Qin, X.; Cao, M.; Lai, F.; Yang, F.; Ge, W.; Zhang, X.; Cheng, S.; Sun, X.; Qin, G.; Shen, W.; et al. Oxidative stress induced by zearalenone in porcine granulosa cells and its rescue by curcumin in vitro. PLoS ONE 2015, 10, e0127551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, J.A.; Griffin, P.M.; Angulo, F.J. Bacterial contamination of animal feed and its relationship to human foodborne illness. Clin. Infect. Dis. 2002, 35, 859–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeleliere, E.; Cobbina, S.J.; Abubakari, Z.I. Review of microbial food contamination and food hygiene in selected capital cities of Ghana. Cogent Food Agric. 2017, 3, 1395102. [Google Scholar] [CrossRef]
- Maciorowski, K.G.; Herrera, P.; Jones, F.T.; Pillai, S.; Ricke, S. Effects on poultry and livestock of feed contamination with bacteria and fungi. Anim. Feed Sci. Technol. 2007, 133, 109–136. [Google Scholar] [CrossRef]
- Zhang, G.; Meredith, T.C.; Kahne, D. On the essentiality of lipopolysaccharide to gram-negative bacteria. Curr. Opin. Microbiol. 2013, 16, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossol, M.; Heine, H.; Meusch, U.; Quandt, D.; Klein, C.; Sweet, M.J.; Hauschildt, S. Lps-induced cytokine production in human monocytes and macrophages. Crit. Rev. Immunol. 2011, 31, 379–446. [Google Scholar] [CrossRef] [PubMed]
- Abou Elazab, M.F.; Nasr, N.E.; Ahmed, M.S.; Alrashdi, B.M.; Dahran, N.; Alblihed, M.A.; Elmahallawy, E.K. The effects of bacterial lipopolysaccharide (lps) on turkey poults: Assessment of biochemical parameters and histopathological changes. Vet. Sci. 2022, 9, 240. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.J.; Gropp, J.; Dierick, N.; Costa, L.G.; Martelli, G.; Brantom, P.G.; Bampidis, V.; Renshaw, D.W.; Leng, L. Risks associated with endotoxins in feed additives produced by fermentation. Environ. Health 2016, 15, 5. [Google Scholar] [CrossRef] [Green Version]
- Luchi, M.; Morrison, D.C. Comparable endotoxic properties of lipopolysaccharides are manifest in diverse clinical isolates of gram-negative bacteria. Infect. Immun. 2000, 68, 1899–1904. [Google Scholar] [CrossRef] [Green Version]
- Svensson, M.; Han, L.; Silfversparre, G.; Häggström, L.; Enfors, S.O. Control of endotoxin release in Escherichia coli fed-batch cultures. Bioprocess Biosyst. Eng. 2005, 27, 91–97. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Schatzmayr, G.; Taranu, I.; Marin, D.; Puel, O.; Oswald, I.P. Mycotoxins co-contamination: Methodological aspects and biological relevance of combined toxicity studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3489–3507. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and inflammatory effects of individual and combined exposure of hepg2 cells to zearalenone and its metabolites. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.D.; Jaykus, L.A. Virus–bacteria interactions: Implications and potential for the applied and agricultural sciences. Viruses 2018, 10, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kew, M.C. Synergistic interaction between aflatoxin b1 and hepatitis b virus in hepatocarcinogenesis. Liver Int. 2003, 23, 405–409. [Google Scholar] [CrossRef]
- Granados-Chinchilla, F.; Valenzuela-Martínez, C.; García-Murillo, B.; Aguilar-Madrigal, D.; Redondo-Solano, M.; Molina, A. Microbiological safety and presence of major mycotoxins in animal feed for laboratory animals in a developing country: The case of Costa Rica. Animals 2021, 11, 2389. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Sulyok, M.; Böhm, J. Mycotoxin cocktail in the samples of oilseed cake from early maturing cotton varieties associated with cattle feeding problems. Toxins 2015, 7, 2188–2197. [Google Scholar] [CrossRef] [Green Version]
- Cegielska-Radziejewska, R.; Stuper-Szablewska, K.; Szablewski, T. Microflora and mycotoxin contamination in poultry feed mixtures from western Poland. Ann. Agric. Environ. Med. AAEM 2013, 20, 30–35. [Google Scholar]
- Gagiu, V.; Avram, M.; Iorga, E.; Belc, N.; Diaconu, M.; Diaconu, A.; Pricop, M. Microbial and mycotoxin contamination of maize during storage. Rom. Biotechnol. Lett. 2007, 12, 3389–3394. [Google Scholar]
- Taranu, I.; Marin, D.; Pistol, G.C.; Untea, A.; Vlassa, M.; Filip, M.; Gras, M.; Rotar, C.; Anghel, A.C. Assessment of the ability of dietary yeast-fermented rapeseed meal to modulate inflammatory and oxidative stress in piglets after weaning. J. Anim. Feed Sci. 2022, 31, 109–122. [Google Scholar] [CrossRef]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of zearalenone and its derivatives on porcine immune response. Toxicol. Vitr. 2011, 25, 1981–1988. [Google Scholar] [CrossRef]
- Marin, D.E.; Pistol, G.C. Effect of sea buckthorn meal extract in alleviating the toxic effect of ochratoxin a and zearalenone in porcine peripheral blood mononuclear cells. Arch. Zootech. 2021, 24, 84–92. [Google Scholar] [CrossRef]
- Marin, D.E.; Bulgaru, C.V.; Anghel, C.A.; Pistol, G.C.; Dore, M.I.; Palade, M.L.; Taranu, I. Grape seed waste counteracts aflatoxin b1 toxicity in piglet mesenteric lymph nodes. Toxins 2020, 12, 800. [Google Scholar] [CrossRef] [PubMed]
- Abdolshahi, A.; Behdad Shokrollahi, Y. Chapter 2—Food contamination. In Mycotoxins and Food Safety; Suna, S., Ed.; IntechOpen: Rijeka, Croatia, 2020. [Google Scholar]
- Todd, E.C.D. Foodborne Diseases: Overview of Biological Hazards and Foodborne Diseases. In Encyclopedia of Food Safety; Academic Press: Cambridge, MA, USA, 2014; pp. 221–242. [Google Scholar] [CrossRef]
- Erceg, S.; Mateo, E.M.; Zipancic, I.; Rodríguez Jiménez, F.J.; Pérez Aragó, M.A.; Jiménez, M.; Soria, J.M.; Garcia-Esparza, M. Assessment of toxic effects of ochratoxin a in human embryonic stem cells. Toxins 2019, 11, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Boesch-Saadatmandi, C.; Lou, Y.; Wolffram, S.; Huebbe, P.; Rimbach, G. Ochratoxin a induces apoptosis in neuronal cells. Genes Nutr. 2009, 4, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Qian, G.; Liu, D.; Hu, J.; Gan, F.; Hou, L.; Chen, X.; Huang, K. Ochratoxin a-induced autophagy in vitro and in vivo promotes porcine circovirus type 2 replication. Cell Death Dis. 2017, 8, e2909. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Wang, B.; Li, X.; Wang, T.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. Zearalenone promotes cell proliferation or causes cell death? Toxins 2018, 10, 184. [Google Scholar] [CrossRef] [Green Version]
- Munshi, N.; Fernandis, A.Z.; Cherla, R.P.; Park, I.-W.; Ganju, R.K. Lipopolysaccharide-induced apoptosis of endothelial cells and its inhibition by vascular endothelial growth factor. J. Immunol. 2002, 168, 5860–5866. [Google Scholar] [CrossRef] [Green Version]
- Assaf, H.; Azouri, H.; Pallardy, M. Ochratoxin a induces apoptosis in human lymphocytes through down regulation of bcl-xl. Toxicol. Sci. 2004, 79, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Oh, S.Y.; Jo, I. Zearalenone induces endothelial cell apoptosis through activation of a cytosolic ca(2+)/erk1/2/p53/caspase 3 signaling pathway. Toxins 2021, 13, 187. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, H.; Wang, J.; Han, S.; Zhang, Y.; Ma, M.; Zhu, Q.; Zhang, K.; Yin, H. Zearalenone induces apoptosis and cytoprotective autophagy in chicken granulosa cells by pi3k-akt-mtor and mapk signaling pathways. Toxins 2021, 13, 199. [Google Scholar] [CrossRef]
- Cimbalo, A.; Alonso Garrido, M.; Font, G.; Manyes, L. Toxicity of mycotoxins in vivo on vertebrate organisms: A review. Food Chem. Toxicol. 2020, 137, 111161. [Google Scholar] [CrossRef] [PubMed]
- Amadori, M.; Filipe, J.; Riva, F.; Vitali, A.; Ruggeri, J.; Lacetera, N. Characterization of the blastogenic response to lps of bovine peripheral blood mononuclear cells. PLoS ONE 2018, 13, e0204827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, Y.; Cui, J.; Xing, L.; Shen, H.; Wu, S.; Lian, H.; Wang, J.; Yan, X.; Zhang, X. Ochratoxin a induces oxidative DNA damage and g1 phase arrest in human peripheral blood mononuclear cells in vitro. Toxicol. Lett. 2012, 211, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.D. Reactive oxygen species and programmed cell death. Trends Biochem. Sci. 1996, 21, 83–86. [Google Scholar] [CrossRef]
- Feng, Y.-Q.; Zhao, A.-H.; Wang, J.-J.; Tian, Y.; Yan, Z.-H.; Dri, M.; Shen, W.; de Felici, M.; Li, L. Oxidative stress as a plausible mechanism for zearalenone to induce genome toxicity. Gene 2022, 829, 146511. [Google Scholar] [CrossRef]
- Bromfield, J.J.; Iacovides, S.M. Evaluating lipopolysaccharide-induced oxidative stress in bovine granulosa cells. J. Assist. Reprod. Genet. 2017, 34, 1619–1626. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Ngkelo, A.; Meja, K.; Yeadon, M.; Adcock, I.; Kirkham, P.A. Lps induced inflammatory responses in human peripheral blood mononuclear cells is mediated through nox4 and giα dependent pi-3kinase signalling. J. Inflamm. 2012, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Kalliolias, G.D.; Ivashkiv, L.B. Tnf biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Xu, Q.; Choksi, S.; Liu, Z. Switching from tnf-induced inflammation to death signaling. Mol. Cell. Oncol. 2018, 5, e1392402. [Google Scholar] [CrossRef]
- Xaus, J.; Comalada, M.; Valledor, A.F.; Lloberas, J.; López-Soriano, F.; Argilés, J.M.; Bogdan, C.; Celada, A. Lps induces apoptosis in macrophages mostly through the autocrine production of tnf-alpha. Blood 2000, 95, 3823–3831. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marin, D.E.; Pistol, G.C.; Procudin, C.V.; Taranu, I. Co-Contamination of Food and Feed with Mycotoxin and Bacteria and Possible Implications for Health. Agriculture 2022, 12, 1970. https://doi.org/10.3390/agriculture12111970
Marin DE, Pistol GC, Procudin CV, Taranu I. Co-Contamination of Food and Feed with Mycotoxin and Bacteria and Possible Implications for Health. Agriculture. 2022; 12(11):1970. https://doi.org/10.3390/agriculture12111970
Chicago/Turabian StyleMarin, Daniela Eliza, Gina Cecilia Pistol, Cristina Valeria Procudin, and Ionelia Taranu. 2022. "Co-Contamination of Food and Feed with Mycotoxin and Bacteria and Possible Implications for Health" Agriculture 12, no. 11: 1970. https://doi.org/10.3390/agriculture12111970