

A Review of Key Techniques for in Ovo Sexing of Chicken Eggs

Abstract
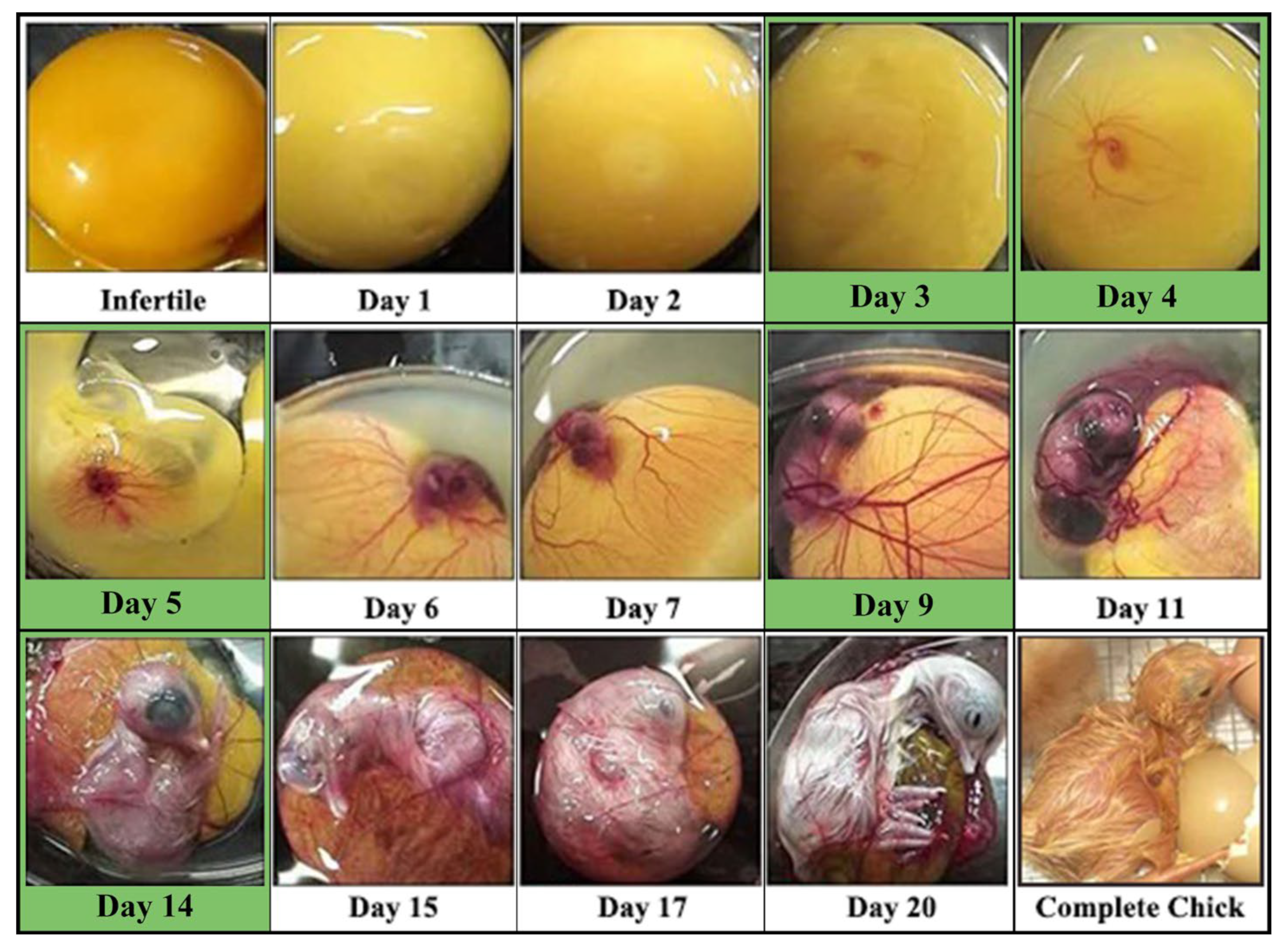
:1. Introduction
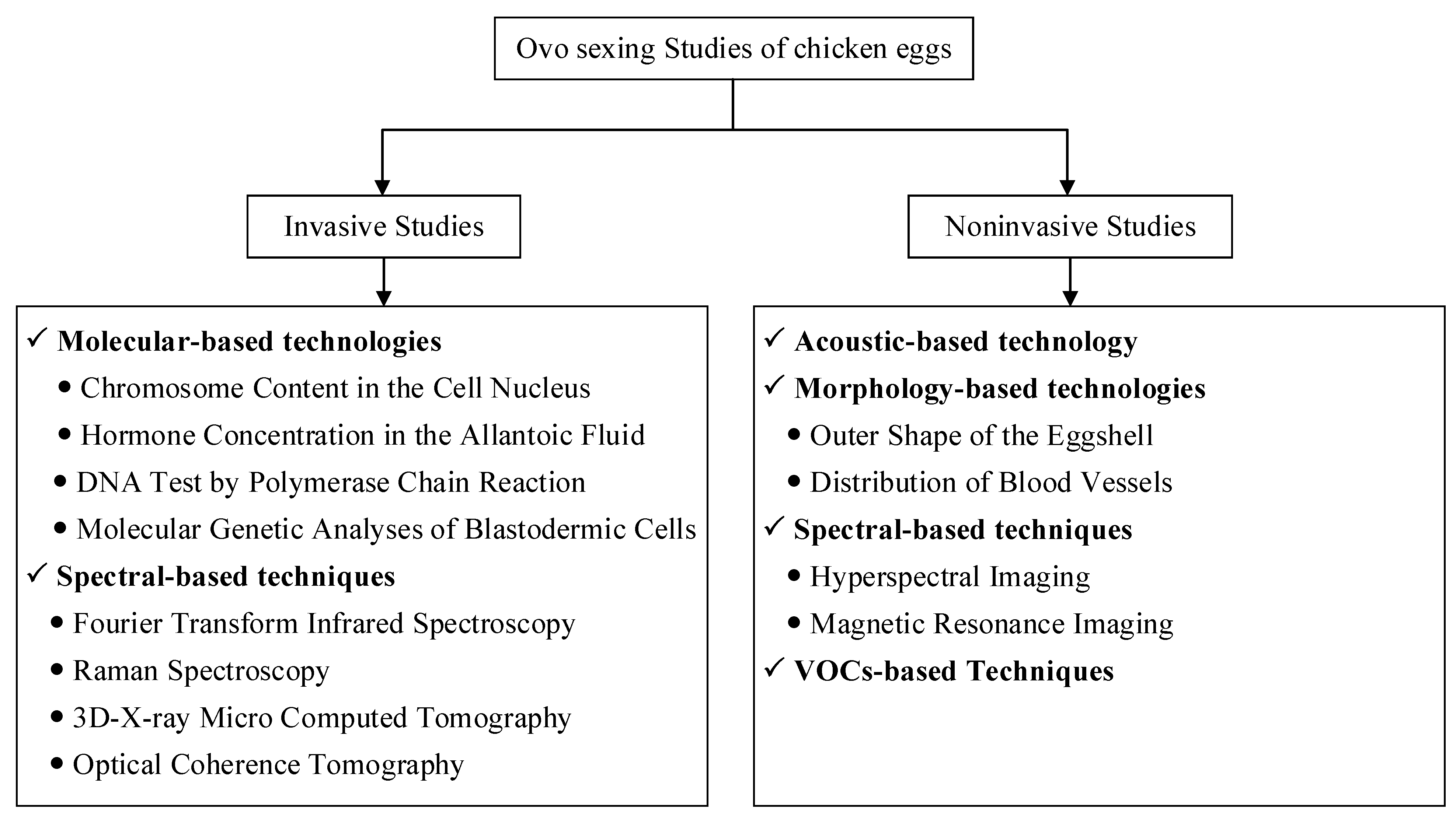
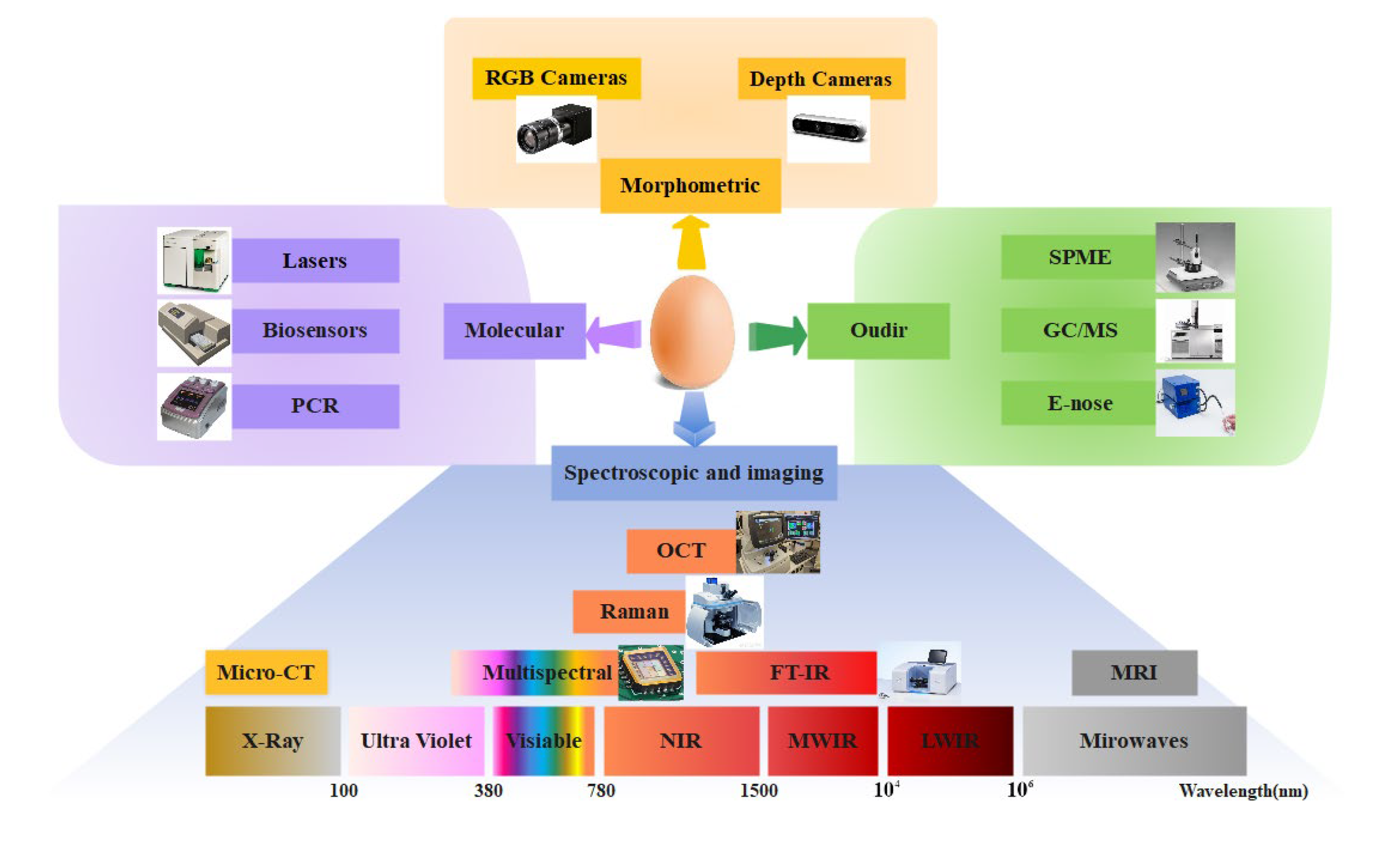
2. Research Progress of Chicken in Ovo Sexing Methods
2.1. Molecular-Based Techniques
2.1.1. Chromosome Content in the Cell Nucleus
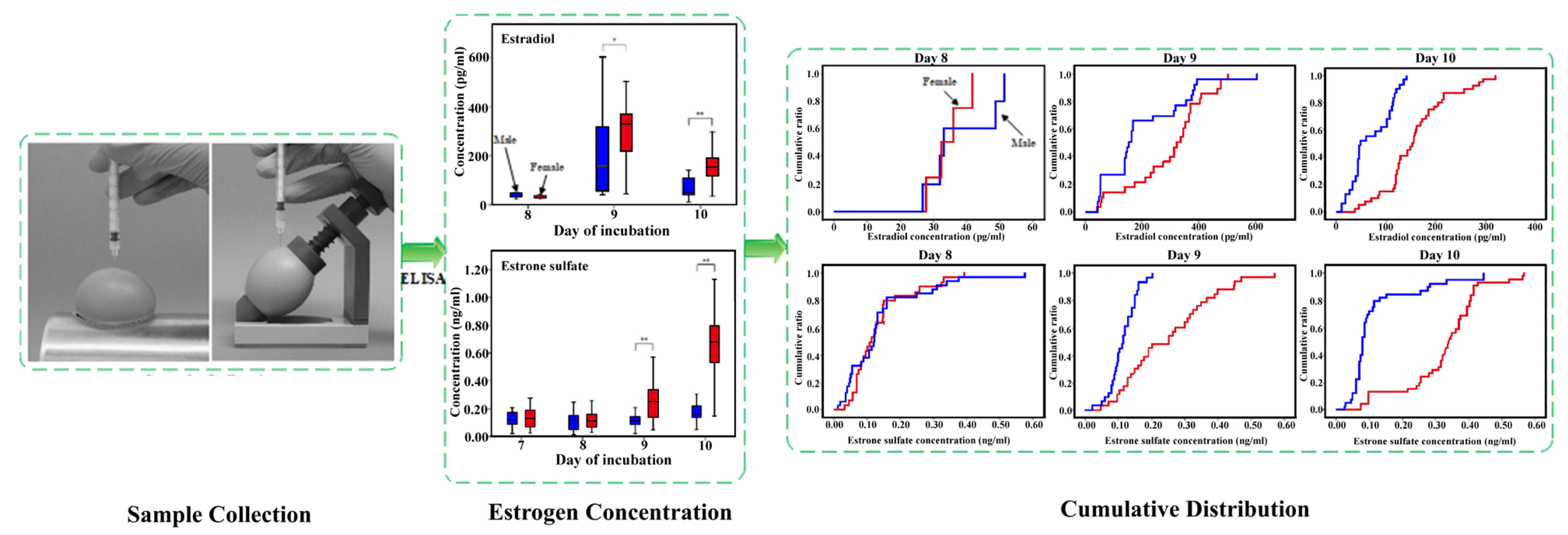
2.1.2. Hormone Concentration in the Allantoic Fluid
2.1.3. DNA Test by Polymerase Chain Reaction

| Categories | Samples | Methods | Results | Reference |
|---|---|---|---|---|
| Nuclear DNA content in the cell | Blood samples from birds | The flow cytometric technique was used to estimate the nuclear DNA content of erythrocytes in blood samples. | The samples were stored at 4–20 °C and successfully analyzed after at least five days. The flow cytometric technique can be completed in a few minutes. | [3] |
| Hormone concentration in the allantoic fluid | Allantoic fluid from eggs | Allantoic fluid was collected and estrone sulfate concentration was measured on day 9 of incubation, showing that male embryos displayed significantly lower hormone levels than female embryos. | This method can detect sex differences by day 9 with an accuracy greater than 98%. The fluid analysis required 4 h and reduced the hatching rate by 3%. | [4] |
| DNA test by PCR | Blood from ISA brown chicken eggs | The Hologic Invader® sexing assay, which includes a W-rpt/CR2 probe mixture and core reagent kit, was developed to monitor fluorescence. | This assay can distinguish the sex of individual animals with as little as 1 ng of DNA or 125 nl of whole blood or as few as 250 cells. | [5] |
| ISA brown, Dekalb white, Bovan brown, and Shaver black eggs | The Q-PCR technique was used to incubate beads chelated with potential PCR inhibitors followed by centrifugation to identify genes on chromosomes W (SWIM and Xho-I) and Z (DMRT). | The Q-PCR method exhibits 100% concordance and specificity for the in ovo sexing of 176 embryos. | [28] | |
| Fertilized white Leghorn chicken eggs | The qRT–PCR technique was performed on mRNA from chick gonads and other tissues. | The qRT–PCR results of each sample agreed with those obtained by morphological examinations and PCR analyses. | [29] |
2.1.4. Molecular Genetic Analyses of Blastodermic Cells
2.2. Spectral-Based Techniques
2.2.1. Fourier Transform Infrared Spectroscopy
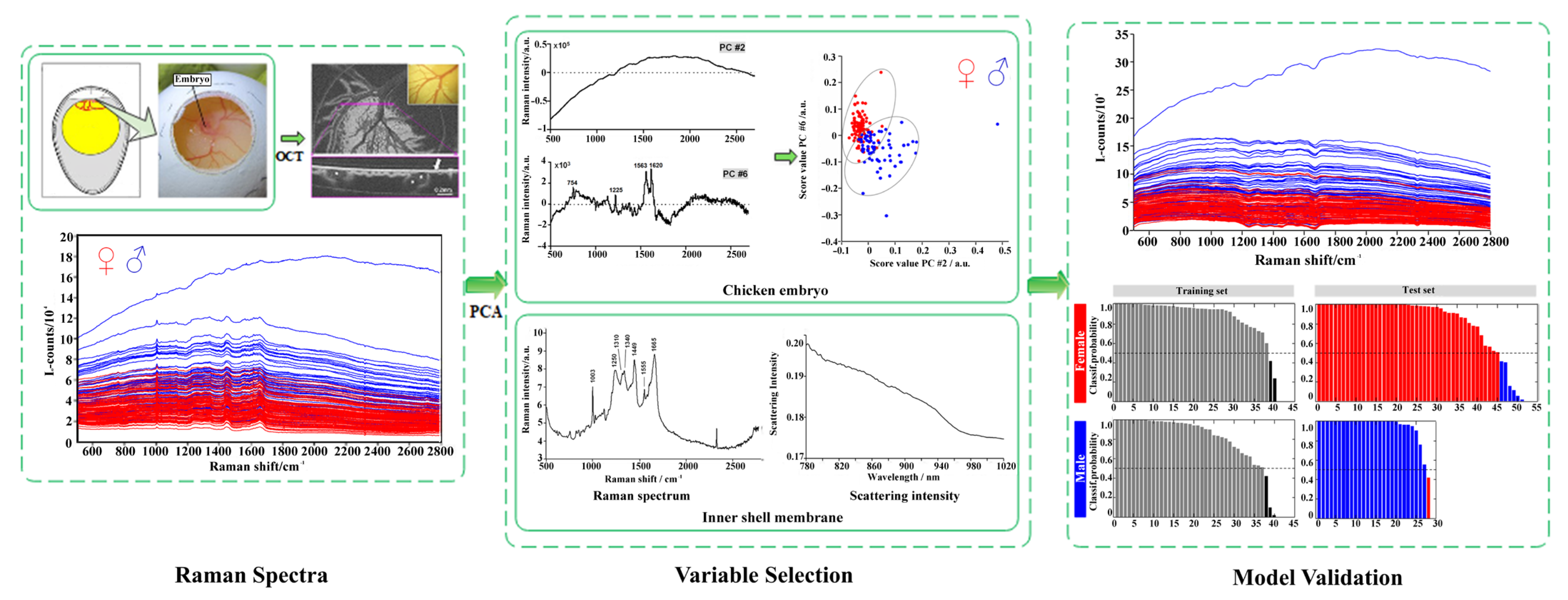
2.2.2. Raman Spectroscopy
2.2.3. Hyperspectral Imaging
2.2.4. 3D X-Ray Microcomputed Tomography
2.2.5. Optical Coherence Tomography
2.2.6. Magnetic Resonance Imaging
2.3. Acoustic-Based Techniques
2.4. Morphology-Based Techniques
2.4.1. Outer Shape of the Eggshell
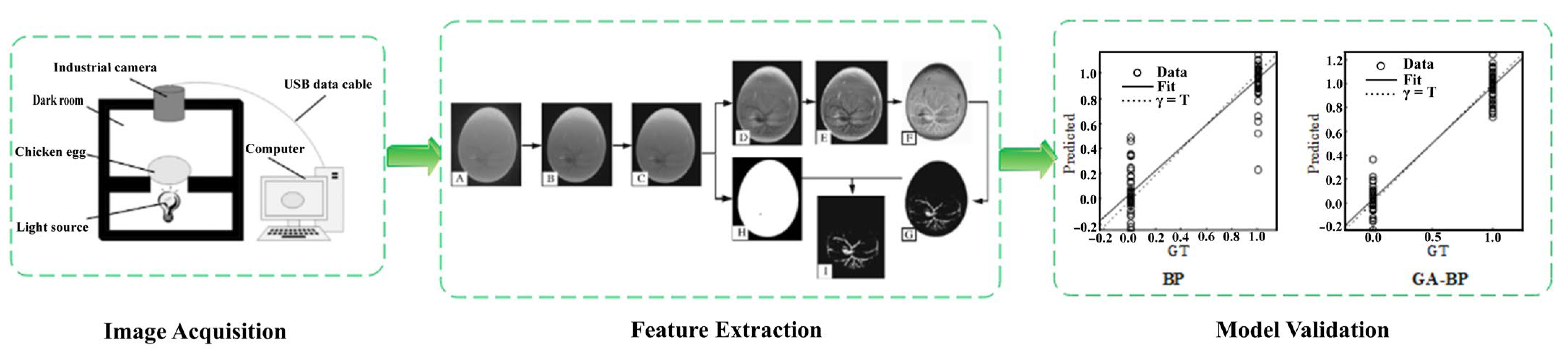
2.4.2. Distribution of Blood Vessels
2.5. Volatile-Organic-Compound-Based Techniques
3. Breakthroughs and Perspectives
3.1. Mechanisms: Analysis of Physiological Changes in Chicken Eggs during Incubation
3.2. Big Data Support: From Sensing to Modeling
3.2.1. Multisensing Techniques
3.2.2. Data Processing
3.2.3. Modeling and Decision-Making
3.3. Commercialization and Practical Applications
3.4. Prospective
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Krautwald-Junghanns, M.-E.; Cramer, K.; Fischer, B.; Förster, A.; Galli, R.; Kremer, F.; Mapesa, E.U.; Meissner, S.; Preisinger, R.; Preusse, G.; et al. Current approaches to avoid the culling of day-old male chicks in the layer industry, with special reference to spectroscopic methods. Poult Sci. 2018, 97, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kaleta, E.F.; Redmann, T. Approaches to determine the sex prior to and after incubation of chicken eggs and of day-old chicks. Worlds Poult. Sci. J. 2008, 64, 391–399. [Google Scholar] [CrossRef]
- Redelman, D.; Fleury, S.A.; Garner, D.L. Flow cytometry for sexing birds. Trends Ecol. Evol. 1997, 12, 489. [Google Scholar] [CrossRef]
- Weissmann, A.; Reitemeier, S.; Hahn, A.; Gottschalk, J.; Einspanier, A. Sexing domestic chicken before hatch: A new method for in ovo gender identification. Theriogenology 2013, 80, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Clinton, M.; Nandi, S.; Zhao, D.; Olson, S.; Peterson, P.; Burdon, T.; McBride, D. Real-Time Sexing of Chicken Embryos and Compatibility with in ovo Protocols. Sex Dev. 2016, 10, 210–216. [Google Scholar] [CrossRef]
- Burkhardt, A.; Geissler, S.; Koch, E. Optical coherence tomography as approach for the minimal invasive localization of the germinal disc in ovo before chicken sexing. Biophotonics Photonic Solut. Better Health Care II SPIE 2010, 7715, 77151W. [Google Scholar] [CrossRef]
- Steiner, G.; Bartels, T.; Stelling, A.; Krautwald-Junghanns, M.E.; Fuhrmann, H.; Sablinskas, V.; Koch, E. Gender determination of fertilized unincubated chicken eggs by infrared spectroscopic imaging. Anal. Bioanal. Chem. 2011, 400, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Krautwald-Junghanns, M.E.; Cramer, K.; Fischer, B.; Förster, A.; Galli, R.; Kremer, F.; Bartels, T. Spektroskopische Methoden zur Geschlechtsfrühdiagnose in der Legehennenvermehrung. Rundsch Fleischhyg Leb. 2014, 66, 440–442. [Google Scholar]
- Galli, R.; Preusse, G.; Uckermann, O.; Bartels, T.; Krautwald-Junghanns, M.-E.; Koch, E.; Steiner, G. In Ovo Sexing of Domestic Chicken Eggs by Raman Spectroscopy. Anal. Chem. 2016, 88, 8657–8663. [Google Scholar] [CrossRef] [PubMed]
- Galli, R.; Koch, E.; Preusse, G.; Schnabel, C.; Bartels, T.; Krautwald-Junghanns, M.-E.; Steiner, G. Contactless in ovo sex determination of chicken eggs. Curr. Dir. BioMed Eng. 2017, 3, 131–134. [Google Scholar] [CrossRef]
- Galli, R.; Preusse, G.; Schnabel, C.; Bartels, T.; Cramer, K.; Krautwald-Junghanns, M.-E.; Koch, E.; Steiner, G. Sexing of chicken eggs by fluorescence and Raman spectroscopy through the shell membrane. PLoS ONE 2018, 13, e0192554. [Google Scholar] [CrossRef]
- Bartels, T.; Fischer, B.; Krüger, P.; Koch, E.; Ryll, M.; Krautwald-Junghanns, M. 3D-X-ray microcomputer tomography and optical coherence tomography as methods for the localization of the blastoderm in the newly laid unincubated chicken egg. Dtsch Tierarztl Wochenschr. 2008, 5, 182–188. [Google Scholar]
- Glahn, R.P.; Mitsos, W.J.; Wideman, J.R.R.F. Evaluation of sex differences in embryonic heart rates. Poult Sci. 1987, 66, 1398–1401. [Google Scholar] [CrossRef] [PubMed]
- Imholt, D. Morphometrische Studien an Eiern von Hybrid-und Rassehühnern mit Versuchen zur Detektion einer Beziehung zwischen der Form von Eiern und dem Geschlecht der darin befindlichen Küken: Eine oologische und mathematische Studie. VVB Laufersweiler. Published online 2010. Available online: http://geb.uni-giessen.de/geb/volltexte/2010/7760/ (accessed on 8 February 2023).
- Yilmaz-Dikmen, B.; Dikmen, S. A morphometric method of sexing white layer Eggs. Rev. Bras. Cienc. Avic. 2013, 15, 203–210. [Google Scholar] [CrossRef]
- Tang, Y.; Hong, Q.; Wang, Q.; Zhu, Z. Sex identification of chicken eggs based on blood line texture features and GA-BP neural network. J. Huazhong Agric. Univ. 2018, 37, 130–135. [Google Scholar]
- Göhler, D.; Fischer, B.; Meissner, S. In-ovo sexing of 14-day-old chicken embryos by pattern analysis in hyperspectral images (VIS/NIR spectra): A non-destructive method for layer lines with gender-specific down feather color. Poult. Sci. 2017, 96, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Davenel, A.; Eliat, P.A.; Quellec, S.; Nys, Y. Attempts for early gender determination of chick embryos in ovo using Magnetic Resonance Imaging. In Proceedings of the XXII European Symposium on the Quality of Poultry Meat & XVI European Symposium on the Quality of Eggs and Egg Products, Nantes, France, 10–13 May 2015. [Google Scholar]
- Xiang, X.; Wang, Y.; Yu, Z.; Ma, M.; Zhu, Z.; Jin, Y. Non-destructive characterization of egg odor and fertilization status by SPME/GC-MS coupled with electronic nose. J. Sci. Food Agric. 2019, 99, 3264–3275. [Google Scholar] [CrossRef]
- Nandi, S.; McBride, D.; Blanco, R.; Clinton, M. Sex diagnosis and sex determination. Worlds Poult. Sci. J. 2003, 59, 7–13. [Google Scholar]
- Tiersch, T.R. Identification of sex in chickens by flow cytometry. Worlds Poult. Sci. J. 2003, 59, 25–32. [Google Scholar]
- Phelps, P.; Bhutada, A.; Bryan, S.; Chalker, A.; Ferrell, B.; Neuman, S.; Ricks, C.; Tran, H.; Butt, T. Automated identification of male layer chicks prior to hatch. Worlds Poult. Sci. J. 2003, 59, 33–38. [Google Scholar]
- Weissmann, A.; Förster, A.; Gottschalk, J.; Reitemeier, S.; Krautwald-Junghanns, M.-E.; Preisinger, R.; Einspanier, A. In ovo-gender identification in laying hen hybrids: Effects on hatching and production performance. Eur. Poult. 2014, 78, 25. [Google Scholar] [CrossRef]
- Rivers, J.W.; Houtz, J.L.; Betts, M.G.; Horton, B.M. No evidence for a link between forest herbicides and offspring sex ratio in a migratory songbird using high-throughput molecular sexing. Conserv. Physiol. 2017, 5, cox054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, R. The isolation of conserved DNA sequences related to the human sex-determining region Y gene from the lesser black-backed gull (Larus fuscus). Proc. R Soc. Lond. Ser. Biol. Sci. 1991, 244, 123–128. [Google Scholar] [CrossRef]
- Clinton, M.; Haines, L.; Belloir, B.; McBride, D. Sexing chick embryos: A rapid and simple protocol. Br. Poult. Sci. 2001, 42, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Turkyilmaz, M.K.; Karagenc, L.; Fidan, E. Sexing of newly-hatched chicks using DNA isolated from chorio-allantoic membrane samples by polymerase chain reaction in Denizli chicken. Br. Poult. Sci. 2010, 51, 525–529. [Google Scholar] [CrossRef]
- He, L.; Martins, P.; Huguenin, J.; Van, T.-N.; Manso, T.; Galindo, T.; Gregoire, F.; Catherinot, L.; Molina, F.; Julien, E. Simple, sensitive and robust chicken specific sexing assays, compliant with large scale analysis. PLoS ONE 2019, 14, e0213033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Z.; Lu, Y.; Rui, L.; Yu, X.; Li, Z. Sexing chick mRNA: A protocol based on quantitative real-time polymerase chain reaction. Poult. Sci. 2017, 96, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Harrisson, F.; Vakaet, L. The chicken blastoderm: In vitro methods of investigation of developmental processes. Biol. Struct. Morphog. 1989, 1, 1–6. [Google Scholar]
- Bartels, T.; Fischer, B.; Krüger, P.; Koch, E.; Ryll, M.; Krautwald-Junghanns, M.E. 3D-Röntgen-Mikrocomputertomographie und Optische Kohä renztomographie als Methoden zur Lagebestimmung des Blastoderms im unbebrüteten Hühnerei. Dtsch. Tierärztliche Wochenschr. 2008, 115, 182–188. [Google Scholar] [CrossRef]
- Harz, M.; Krause, M.; Bartels, T.; Cramer, K.; Rösch, P.; Popp, J. Minimal Invasive Gender Determination of Birds by Means of UV-Resonance Raman Spectroscopy. Anal. Chem. 2008, 4, 1080–1086. [Google Scholar] [CrossRef]
- Steiner, G.; Bartels, T.; Krautwald-Junghanns, M.-E.; Boos, A.; Koch, E. Sexing of turkey poults by Fourier transform infrared spectroscopy. Anal. Bioanal. Chem. 2010, 396, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Morita, V.S.; Boleli, I.C.; Cargnelutti Filho, A. Hematological values and body, heart and liver weights of male and female broiler embryos of young and old breeder eggs. Rev. Bras. Cienc. Avic. 2009, 11, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, Q.; Ma, M.; Xiao, S.; Shi, H. Non-Destructive Detection of Male and Female Information of Early Duck Embryos Based on Visible/Near Infrared Spectroscopy and Deep Learning. Spectrosc. Spectr. Anal. 2021, 41, 1800–1805. [Google Scholar]
- Alin, K.; Fujitani, S.; Kashimori, A.; Suzuki, T.; Ogawa, Y.; Kondo, N. Non-invasive broiler chick embryo sexing based on opacity value of incubated eggs. Comput. Electron. Agric. 2019, 158, 30–35. [Google Scholar] [CrossRef]
- Rahman, A.; Syduzzaman, M.; Khaliduzzaman, A.; Fujitani, S.; Kashimori, A.; Suzuki, T.; Ogawa, Y.; Kondo, N. Nondestructive sex-specific monitoring of early embryonic development rate in white layer chicken eggs using visible light transmission. Br. Poult. Sci. 2020, 61, 209–216. [Google Scholar] [CrossRef]
- Corion, M.; Keresztes, J.; De Ketelaere, B.; Saeys, W. In ovo sexing of eggs from brown breeds with a gender-specific color using visible-near-infrared spectroscopy: Effect of incubation day and measurement configuration. Poult. Sci. 2022, 101, 101782. [Google Scholar] [CrossRef]
- Preuße, G.; Porstmann, V.; Bartels, T.; Schnabel, C.; Galli, R.; Koch, E.; Oelschlägel, M.; Uckermann, O.; Steiner, G. Highly sensitive and quick in ovo sexing of domestic chicken eggs by two-wavelength fluorescence spectroscopy. Anal. Bioanal. Chem. 2022, 415, 603–613. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, W.; Yu, M.; Sun, Y.; Gu, X.; Ma, L.; Li, Z.; Hu, P.; Tu, K. Gender determination of early chicken hatching eggs embryos by hyperspectral imaging. Nongye Gongcheng Xuebao/Trans. Chin. Soc. Agric. Eng. 2016, 32, 181–186. [Google Scholar] [CrossRef]
- Zhu, Z.; Hong, Q.; Wu, L.; Wang, Q.; Ma, M. Early Identification of Male and Female Embryos Based on UV/Vis Transmission Spectroscopy and Extreme Learning Machine. Spectrosc. Spectr. Anal. 2019, 39, 2780–2787. [Google Scholar]
- Geng, L.; Hu, Y.; Xi, J.; Liu, Y. Detection on the Fertility of Hatching Eggs Based on Heart Rate Threshold. Am. Sci. Res. J. Eng. Technol. Sci. 2019, 53, 49–63. [Google Scholar]
- Mappatao, G. Duck Egg Sexing by Eccentricity Determination Using Image Processing. J. Telecommun. Electron. Comput. Eng. 2018, 10, 1–9. [Google Scholar]
- Dioses, J.L.; Medina, R.P.; Fajardo, A.C.; Hernandez, A.A. Performance of Classification Models in Japanese Quail Egg Sexing. In Proceedings of the 2021 IEEE 17th International Colloquium on Signal Processing & Its Applications (CSPA), Langkawi, Malaysia, 5–6 March 2021; pp. 29–34. [Google Scholar] [CrossRef]
- Pardo, J.I.S.; González, F.J.N.; Ariza, A.G.; Arbulu, A.A.; Jurado, J.M.L.; Bermejo, J.V.D.; Vallejo, M.E.C. Traditional sexing methods and external egg characteristics combination allow highly accurate early sex determination in an endangered native turkey breed. Front. Vet. Sci. 2022, 9, 948502. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Tang, Y.; Hong, Q.; Huang, P.; Wang, Q.; Ma, M. Female and male identification of early chicken embryo based on blood line features of hatching egg image and deep belief networks. Trans. Chinese Soc. Agric. Eng. 2018, 34, 197–203. [Google Scholar]
- Yimenu, S.M.; Kim, J.Y.; Kim, B.S. Prediction of egg freshness during storage using electronic nose. Poult. Sci. 2017, 96, 3733–3746. [Google Scholar] [CrossRef]
- Cavanna, D.; Zanardi, S.; Dall’Asta, C.; Suman, M. Ion mobility spectrometry coupled to gas chromatography: A rapid tool to assess eggs freshness. Food Chem. 2019, 271, 691–696. [Google Scholar] [CrossRef]
- Whittaker, D.J.; Soini, H.A.; Gerlach, N.M.; Posto, A.L.; Novotny, M.V.; Ketterson, E.D. Role of Testosterone in Stimulating Seasonal Changes in a Potential Avian Chemosignal. J. Chem. Ecol. 2011, 37, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Webster, B.; Hayes, W.; Pike, T.W. Avian egg odour encodes information on embryo sex, fertility and development. PLoS ONE 2015, 10, e0116345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouda, M.; Ma, M.; Sheng, L.; Xiang, X. SPME-GC-MS & metal oxide E-Nose 18 sensors to validate the possible interactions between bio-active terpenes and egg yolk volatiles. Food Res. Int. 2019, 125, 108611. [Google Scholar] [CrossRef]
- Corion, M.; De Ketelaere, B.; Hertog, M.; Lammertyn, J. Profiling the emission of volatile organic compounds from chicken hatching eggs in the first half of incubation. IFRG eMeeting 2021, 5, 114–122. [Google Scholar]
- Xiang, X.; Hu, G.; Jin, Y.; Jin, G.; Ma, M. Nondestructive characterization gender of chicken eggs by odor using SPME/GC-MS coupled with chemometrics. Poult. Sci. 2022, 101, 101619. [Google Scholar] [CrossRef]
- Hirst, C.E.; Major, A.T.; Smith, C.A. Sex determination and gonadal sex differentiation in the chicken model. Int. J. Dev. Biol. 2018, 62, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Galli, R.; Preusse, G.; Uckermann, O.; Bartels, T.; Krautwald-Junghanns, M.-E.; Koch, E.; Steiner, G. In ovo sexing of chicken eggs by fluorescence spectroscopy. Anal. Bioanal. Chem. 2017, 409, 1185–1194. [Google Scholar] [CrossRef]
- Ayers, K.L.; Sinclair, A.H.; Smith, C.A. The molecular genetics of ovarian differentiation in the avian model. Sex Dev. 2013, 7, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-H.; Kai, Z.; Wu, L.; Wang, C. Egg Freshness Detection Based on Hyper-Spectra. Spectrosc. Spectr. Anal. 2015, 36, 2596–2600. [Google Scholar]
- Barthelmebs, L.; Calas-Blanchard, C.; Istamboulie, G.; Marty, J.L.; Noguer, T. Biosensors as analytical tools in food fermentation industry. Adv. Exp. Med. Biol. 2010, 698, 293–307. [Google Scholar] [CrossRef]
- Kamilaris, A.; Prenafeta-Boldú, F.X. Deep learning in agriculture: A survey. Comput. Electron. Agric. 2018, 147, 70–90. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X. Nondestructive Sexing of Chicken Eggs Based on Odor and Matrix Differences Analyzed by Multi-Omicmulti-Omics. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2019. [Google Scholar]
- Ren, G.; Lin, T.; Ying, Y.; Chowdhary, G.; Ting, K.C. Agricultural robotics research applicable to poultry production: A review. Comput. Electron. Agric. 2020, 169, 105216. [Google Scholar] [CrossRef]
- Ren, S.; He, K.; Girshick, R.; Sun, J. Faster R-CNN: Towards real-time object detection with region proposal networks. Adv. Neural. Inf. Process. Syst. 2015, 2015, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Redmon, J.; Divvala, S.; Girshick, R.; Farhadi, A. You Only Look Once: Unified, Real-Time Object Detection. In Proceedings of the 2016 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), Las Vegas, NV, USA, 27–30 June 2016; pp. 779–788. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Anguelov, D.; Erhan, D.; Szegedy, C.; Reed, S.; Fu, C.Y.; Berg, A.C. SSD: Single shot multibox detector. In Proceedings of the Computer Vision–ECCV 2016: 14th European Conference, Amsterdam, The Netherlands, 11–14 October 2016; pp. 21–37. [Google Scholar] [CrossRef] [Green Version]
- Afsah-Hejri, L.; Hajeb, P.; Ara, P.; Ehsani, R.J. A Comprehensive Review on Food Applications of Terahertz Spectroscopy and Imaging. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1563–1621. [Google Scholar] [CrossRef]
- Afsah-Hejri, L.; Akbari, E.; Toudeshki, A.; Homayouni, T.; Alizadeh, A.; Ehsani, R. Terahertz spectroscopy and imaging: A review on agricultural applications. Comput. Electron. Agric. 2020, 177, 105628. [Google Scholar] [CrossRef]
- Shen, S.; Liu, X.; Shen, Y.; Qu, J.; Pickwell-MacPherson, E.; Wei, X.; Sun, Y. Recent Advances in the Development of Materials for Terahertz Metamaterial Sensing. Adv. Opt. Mater. 2022, 10, 2101008. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, D.; Choi, M.H.; Son, J.H.; Seo, M. Highly Sensitive and Selective Detection of Steroid Hormones Using Terahertz Molecule-Specific Sensors. Anal. Chem. 2019, 91, 6844–6849. [Google Scholar] [CrossRef] [PubMed]
- The SELEGGT Process. Available online: https://www.seleggt.com/seleggt-process/ (accessed on 18 December 2022).
- In Ovo. Available online: https://inovo.nl/ (accessed on 18 December 2022).
- CHEGGY is Ready for Practical Use. Available online: https://www.agri-at.com/en/products/in-ovo-sex-determination/cheggy/70-cheggy-is-ready-for-practical-use?rCH=2 (accessed on 8 February 2023).
- EggXYt. Available online: https://www.eggxyt.com/ (accessed on 8 February 2023).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





| Types | Samples | Incubation Age | Band Range | Data Processing and Modeling | Accuracy (%) | Reference |
|---|---|---|---|---|---|---|
| FT-IR | Blastoderm cells | >3 d | 1790 &1000 cm−1 | Removal of outliers and linear two-point baseline corrections | Not specified | [7] |
| Raman | White eggs | 3.5 d | 785 nm | PCA, genetic optimal region selection, and nonlinear discriminant analysis | 91 | [11] |
| Transmission | Brown eggs | 14 d | 600–950 nm | PCA and a linear discriminant analysis model | 97.47 | [17] |
| Hyperspectral | White eggs from brown chickens | 10 d | 600–900 nm | SVM, PLSDA, and ANN | SVM: 75 PLSDA: 75 ANN: 82.86 | [40] |
| Jing No. 1 chickens | 7 d | 360–780 nm | MSC, CARS, SPA, and GA-ELM | 87.14 | [41] | |
| Guo Shao No. 1 chickens | 7 d | 500–900 nm | SPA+CNN | 93.83 | [35] |
| Categories | Samples | Methods | Results | Reference |
|---|---|---|---|---|
| Outer shape of the eggshell | White eggs | The egg width (W), length (L), weight, and volume were measured to determine the sex of the fertilized white eggs before incubation. The egg volume and shape index were estimated according to these measurements as follows: | A multiple logistic regression model was built to determine the sex of the hatching chick. The female chick sex probability (IP1) was calculated as follows: IP1 = −0.39531+(0.01214 × SI) (R2 = 0.25). Eggs with greater SI values (p = 0.001) and lower volumes (p = 0.004) were more likely to produce female chicks. | [15] |
| Duck eggs | The eccentricity (Ecc) of each sample was used to determine a specific threshold value to separate male and female eggs. | The results show that with an eccentricity threshold value of 0.6441, the duck egg sex prediction accuracy reached up to 86%. | [43] | |
| Japanese quail eggs | The W, L, SI, Ecc, area, geometric mean diameter (Gd), and sphericity were measured and calculated to establish an in ovo sexing model for the eggs. | The Gaussian naïve Bayes model is the best classifier for data using two features (Ecc and SI), achieving an average accuracy of 82.88% (males: 85.14% and females: 80.16%). | [44] | |
| Turkey eggs | The combination of plumage color, physical external egg characteristics, the color of down feathers, and behavioral approaches to determine sex of turkey eggs. | The specificity values were found to be 49.12, 93.33, and 100%, while the sensitivity values were observed to be 74.64, 91.03, and 100%, which translated into accuracy of 63.10, 92.26, and 100% in black, black-roan, and bronze-roan poults, respectively. | [45] | |
| Distribution of blood vessels | Jingfen No. 1 eggs | The 11-dimensional feature parameters of the image were extracted with difference box, gray level co-occurrence matrix, gray histogram, and geometric methods. A GA-BP-based neural network was established for in ovo sexing of chicken eggs on day 4 of incubation. | The classification accuracies of the training and prediction set were 99.73% and 82.80%, respectively. | [16] |
| Jingfen No. 1 eggs | The 2916-dimensional fully informative image features were extracted through a gray horizontal co-occurrence matrix with 5-dimensional features and histogram of gradients (HOG) orientation. In addition, the 96-dimensional features were simplified using sampling and PCA dimensionality reduction-gray co-occurrence matrix methods. Support vector machine (SVM), backpropagation (BP) neural network, and deep belief network (DBN) models were constructed. | The DBN model had the highest accuracies of 76.67% (male) and 90% (female), with an average accuracy of 83.33%. The DBN model had the longest discriminant time of 7.8 s. | [46] |
| Methods | Principles | Merits | Reference |
|---|---|---|---|
| Moving average | is the width of the window | It can directly process all spectra without splitting the test and correction sets. | [57] |
| SG | is the weighting factor in moving window smoothing (window length 2𝑙 + 1) | It is suitable for stable denoising of spectral signals and obtains better effects in terms of eliminating high-frequency noise. | [57] |
| MSC | where denote the relative offset coefficients and translations of after linear regression | This method eliminates the scattering effects caused by uneven distributions. | [41] |
| Differential algorithm | where G is the differential width | This method eliminates the effects of background interference and highlights changes in spectral patterns. | [57] |
| Methods | Characteristic | Application Scope | References |
|---|---|---|---|
| PCA | Transforms large and highly correlated spectral data into feature information with fewer dimensions. | Suitable for multifactor spectral feature extraction, as this approach ensures that the main feature information is retained. | [11] |
| ICA | Decomposes the observed mixed signal into statistically independent components, which can reduce the dimension by eliminating redundant information in the original data. | Utilizes higher-order statistical information, which is more conducive to decomposing the observed signal. | [57] |
| SPA | Extracts several characteristic wavelengths in the full wavelength band to eliminate redundant information in the original spectral matrix. | Eliminates redundant information in the original spectral matrix and can be used to screen spectral feature wavelengths. | [35,41] |
| CARS | The wavelength points with large absolute values of regression coefficients in the PLS model are selected by the ARS technique, which can effectively identify the optimal variable combinations. | Addresses the problem of variable combination explosion in the variable selection process. | [35,41] |
| Type | Model | Methods | Accuracy (%) | Reference |
|---|---|---|---|---|
| Linear | SVM | Hyperspectral imaging | Modeling: 80.65 Validation: 75 | [40] |
| Blood vessel distribution | Modeling: 100 Validation: 63.33 | [46] | ||
| Outer shape of the eggshell | 81.51 | [44] | ||
| Kernel Naïve Bayes | Outer shape of the eggshell | 82.88 | [44] | |
| PLS-DA | Hyperspectral imaging | Modeling: 72.58 Validation: 75 | [40] | |
| Vis-NIR | 99.05 | [38] | ||
| Nonlinear | GA-BP | Blood vessel distribution | Modeling: 99.73 Validation: 82.80 | [46] |
| GA-ELM | Blood vessel distribution | Modeling: 100 Validation: 87.14 | [41] | |
| ANN | Hyperspectral imaging | Modeling: 88.14 Validation: 82.86 | [40] | |
| DBN | Blood vessel distribution | Modeling: 100 Validation: 83.33 | [46] | |
| CNN | VIS/NIR spectra | Modeling: 93.36 Validation: 93.83 | [35] |
| Principle | Incubation Day | Sample | Technique | Invasiveness | Precision | Capacity | Marketing |
|---|---|---|---|---|---|---|---|
| Chromoso-mic | 9 d | Allantoic liquid | PCR on cells suspended in allantoic fluid | Yes | 97-99% | 3000/h | PLANTegg |
| Molecular | 9 d | Allantoic liquid | Determination of estrone sulphate | Yes | 98% | SELEGGT: 3600/h; In Ovo: 1500/h | SELEGGT, In ovo |
| Physiological | 13 d | Whole egg | Hyper-spectral imaging via feather color | No | 95% | 20,000/h | Agri Advanced Technologies |
| Genome editing | 0 d | Whole egg | Imaging by fluorescence of a molecule produced by males after editing | No | 100% | / | EggXYT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, N.; Li, B.; Zhu, J.; Wang, H.; Zhao, Y.; Zhao, W. A Review of Key Techniques for in Ovo Sexing of Chicken Eggs. Agriculture 2023, 13, 677. https://doi.org/10.3390/agriculture13030677
Jia N, Li B, Zhu J, Wang H, Zhao Y, Zhao W. A Review of Key Techniques for in Ovo Sexing of Chicken Eggs. Agriculture. 2023; 13(3):677. https://doi.org/10.3390/agriculture13030677
Chicago/Turabian StyleJia, Nan, Bin Li, Jun Zhu, Haifeng Wang, Yuliang Zhao, and Wenwen Zhao. 2023. "A Review of Key Techniques for in Ovo Sexing of Chicken Eggs" Agriculture 13, no. 3: 677. https://doi.org/10.3390/agriculture13030677