
The Promotive Effect of Putrescine on Growth, Biochemical Constituents, and Yield of Wheat (Triticum aestivum L.) Plants under Water Stress
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Morphological Growth and Yield Traits
2.3. Leaf Pigments
2.4. Total Soluble Sugars (TSS)
2.5. Total Free Amino Acids
2.6. Free Proline
2.7. Total Phenolic Content
2.8. Total Flavonoids
2.9. Protein Profile
2.10. Statistical Analysis
3. Results
3.1. Morphological Characteristics
3.1.1. Plant Height, Root Length, Leaves Number, Flag Leaf Area, and Tillers Number
3.1.2. Weights of Fresh and Dry of Both Shoot and Root
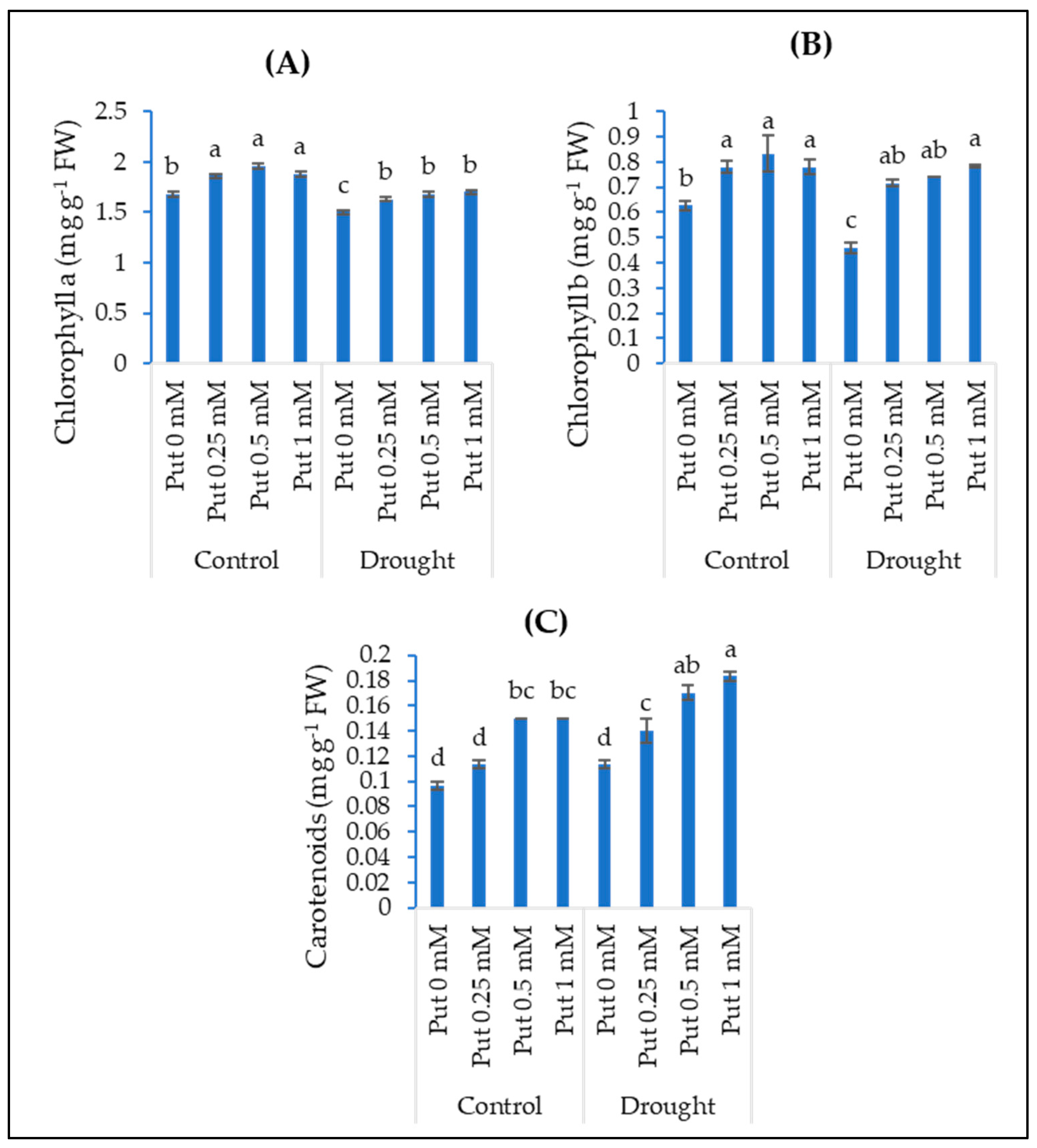
3.2. Photosynthetic Pigments
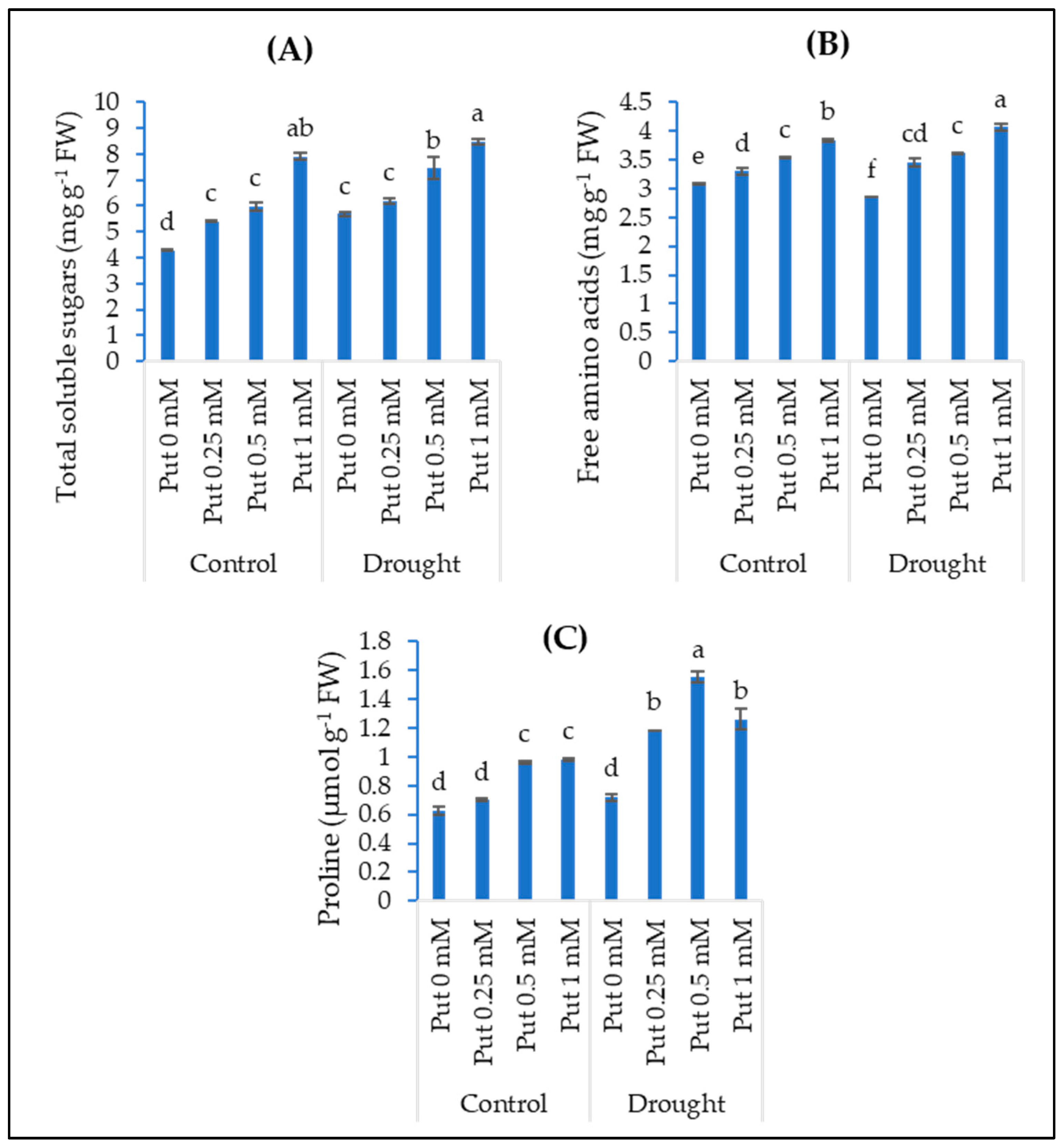
3.3. Osmolytes (Soluble Sugars, Free Amino Acids, Free Proline)
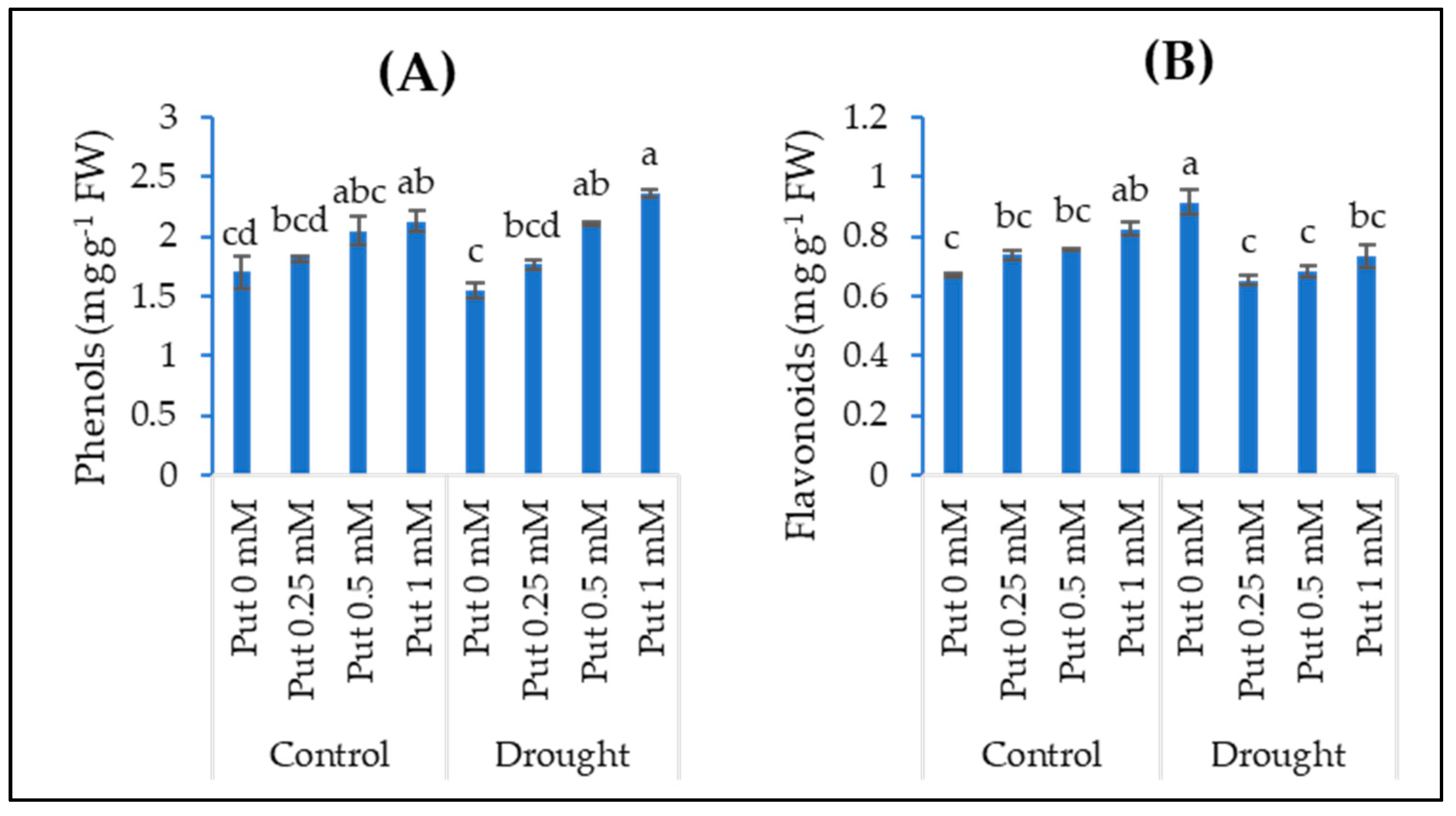
3.4. Phenols and Flavonoids
3.5. Profile of Protein
3.6. Yield Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahari, A.; Pirdashti, H.; Yaghubi, M. The effects of amino acid fertilizers spraying on photosynthetic pigments and antioxidant enzymes of wheat (Triticum aestivum L.) under salinity stress. Int. J. Plant Prod. 2013, 4, 787–793. [Google Scholar]
- Abdel-Fattah, G.M.; Asrar, A.W.A. Arbuscular mycorrhizal fungal application to improve growth and tolerance of wheat (Triticum aestivum L.) plants grown in saline soil. Acta Physiol. Plant. 2012, 34, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Hussein, H.-A.A.; Alshammari, S.O.; Kenawy, S.K.M.; Elkady, F.M.; Badawy, A.A. Grain-priming with L-arginine improves the growth performance of wheat (Triticum aestivum L.) plants under drought stress. Plants 2022, 11, 1219. [Google Scholar] [CrossRef]
- FAO. FAO Publications Catalogue 2022; FAO: Rome, Italy, 2022. [Google Scholar]
- Ewais, E.A.; Ismail, M.A.; Amin, M.A.; Badawy, A.A. Efficiency of salicylic acid and glycine on sugar beet plants grown under heavy metals pollution. Egypt J. Biotechnol. 2015, 48, 112–126. [Google Scholar]
- Abdel Latef, A.A.H.; Omer, A.M.; Badawy, A.A.; Osman, M.S.; Ragaey, M.M. Strategy of salt tolerance and interactive impact of Azotobacter chroococcum and/or Alcaligenes faecalis inoculation on canola (Brassica napus L.) plants grown in saline soil. Plants 2021, 10, 110. [Google Scholar] [CrossRef]
- Attia, M.S.; Hashem, A.H.; Badawy, A.A.; Abdelaziz, A.M. Biocontrol of early blight disease of eggplant using endophytic Aspergillus terreus: Improving plant immunological, physiological and antifungal activities. Bot. Stud. 2022, 63, 26. [Google Scholar] [CrossRef]
- Timmusk, S.; Abd El-Daim, I.A.; Copolovici, L.; Tanilas, T.; Kännaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenström, E.; Niinemets, Ü. Drought-tolerance of wheat improved by rhizosphere bacteria from harsh environments: Enhanced biomass production and reduced emissions of stress volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [Green Version]
- Hassanein, R.A.; Amin, A.B.A.E.S.; Rashad, E.S.M.; Ali, H. Effect of thiourea and salicylic acid on antioxidant defense of wheat plants under drought stress. Int. J. ChemTech Res. 2015, 7, 346–354. [Google Scholar]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought stress tolerance in wheat and barley: Advances in physiology, breeding and genetics research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowron, E.; Trojak, M. Effect of exogenously-applied abscisic acid, putrescine and hydrogen peroxide on drought tolerance of barley. Biologia 2021, 76, 453–468. [Google Scholar] [CrossRef]
- Ayad, H.S.; Reda, F.; Abdalla, M.S.A. Effect of putrescine and zinc on vegetative growth, photosynthetic pigments, lipid peroxidation and essential oil content of gerenium. World J. Agric. Sci. 2010, 6, 601–608. [Google Scholar]
- Ebeed, H.T.; Hassan, N.M.; Aljarani, A.M. Exogenous applications of polyamines modulate drought responses in wheat through osmolytes accumulation, increasing free polyamine levels and regulation of polyamine biosynthetic genes. Plant Physiol. Biochem. 2017, 118, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, H.; Ghorbanpour, M.; Brestic, M. Exogenous putrescine changes redox regulations and essential oil constituents in field-grown Thymus vulgaris L. under well-watered and drought stress conditions. Ind. Crops Prod. 2018, 122, 119–132. [Google Scholar] [CrossRef]
- Li, L.; Gu, W.; Li, C.; Li, W.; Li, C.; Li, J.; Wei, S. Exogenous spermidine improves drought tolerance in maize by enhancing the antioxidant defence system and regulating endogenous polyamine metabolism. Crop Pasture Sci. 2018, 69, 1076–1091. [Google Scholar] [CrossRef]
- Ahmed, A.H.H.; Darwish, E.; Hamoda, S.A.F.; Alobaidy, M.G. Effect of putrescine and humic acid on growth, yield and chemical composition of cotton plants grown under saline soil conditions. Am. J. Agric. Environ. Sci. 2013, 13, 479–497. [Google Scholar]
- Seleem, E.A.; Ibrahim, H.M.S.; Taha, Z.K. Exogenous application of ascorbic acid and putrescine: A natural eco-friendly potential for alleviating NaCl stress in barley (Hordeum vulgare). Emir. J. Food Agric. 2021, 33, 657–670. [Google Scholar] [CrossRef]
- Metzner, H.; Rau, H.; Senger, H. Untersuchungen zur Synchronisierbarkeit einzelner Pigmentmangel-Mutanten von Chlorella. Planta 1965, 65, 186–194. [Google Scholar] [CrossRef]
- Cerning, B.J. A note on sugar determination by the anthrone method. Cereal Chem. 1975, 52, 857–860. [Google Scholar]
- Rosen, H. A modified ninhydrin colorimetric analysis for amino acids. Arch. Biochem. Biophys. 1957, 67, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Savitree, M.; Isara, P.; Nittaya, S.L.; Worapan, S. Radical scavenging activity and total phenolic content of medicinal plants used in primary health care. J. Pharm. Sci. 2004, 9, 32–35. [Google Scholar]
- Pourmorad, F.; Hosseinimehr, S.J.; Shahabimajd, N. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afr. J. Biotechnol. 2006, 5, 1142–1145. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods; The Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Da Costa, M.; Huang, B. Changes in antioxidant enzyme activities and lipid peroxidation for bentgrass species in response to drought stress. J. Am. Soc. Hortic. Sci. 2007, 132, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Doneva, D.; Pál, M.; Brankova, L.; Szalai, G.; Tajti, J.; Khalil, R.; Ivanovska, B.; Velikova, V.; Misheva, S.; Janda, T.; et al. The effects of putrescine pre-treatment on osmotic stress responses in drought-tolerant and drought-sensitive wheat seedlings. Physiol. Plant. 2021, 171, 200–216. [Google Scholar] [CrossRef]
- Kusvuran, S. Microalgae (Chlorella vulgaris Beijerinck) alleviates drought stress of broccoli plants by improving nutrient uptake, secondary metabolites, and antioxidative defense system. Hortic. Plant J. 2021, 7, 221–231. [Google Scholar] [CrossRef]
- Ahmed, A.H.H.; Darwish, E.; Alobaidy, M.G. Impact of putrescine and 24-epibrassinolide on growth, yield and chemical constituents of cotton (Gossypium barbadense L.) plant grown under drought stress conditions. Asian J. Plant Sci. 2017, 16, 9–23. [Google Scholar] [CrossRef] [Green Version]
- Abd Elbar, O.H.; Farag, R.E.; Shehata, S.A. Effect of putrescine application on some growth, biochemical and anatomical characteristics of Thymus vulgaris L. under drought stress. Ann. Agric. Sci. 2019, 64, 129–137. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, V.P.; Gupta, N.K. Efficacy of putrescine and benzyladenine on photosynthesis and productivity in relation to drought tolerance in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2012, 18, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Hong, X.; Xiao-xia, W.; Yun-cheng, L. Effect of polyamine on seed germination of wheat under drought stress is related to changes in hormones and carbohydrates. J. Integr. Agric. 2016, 15, 2759–2774. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.K.; Rahman, M.H.; Rana, M.S.; Lim, J.D.; Lim, Y.S. Exogenous putrescine enhances salt tolerance and ginsenosides content in korean ginseng (Panax ginseng Meyer) sprouts. Plants 2021, 10, 1313. [Google Scholar] [CrossRef]
- Al-Kandari, M.; Redha, A.; Suleman, P. Polyamine accumulation and osmotic adjustment as adaptive responses to water and salinity stress in Conocarpus lancifolius. Funct. Plant Sci. Biotechnol. 2009, 3, 42–48. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Zeid, I.M.; Shedeed, Z.A. Response of alfalfa to putrescine treatment under drought stress. Biol. Plant. 2006, 50, 635–640. [Google Scholar] [CrossRef]
- El-Bassiouny, H.M.; Mostafa, H.A.; El-Khawas, S.A.; Hassanein, R.A.; Khalil, S.I.; Abd El-Monem, A.A. Physiological responses of wheat plant to foliar treatments with arginine or putrescine. Aust. J. Basic Appl. Sci. 2008, 2, 1390–1403. [Google Scholar]
- Talaat, I.M.; Bekheta, M.A.; Mahgoub, M.H. Physiological response of periwinkle plants (Catharanthus roseus L.) to tryptophan and putrescine. Int. J. Agric. Biol 2005, 7, 210–213. [Google Scholar]
- Shallan, M.A.; Hassan, H.M.M.; Namich, A.A.M.; Ibrahim, A.A. Effect of sodium niroprusside, putrescine and glycine betaine on alleviation of drought stress in cotton plant. Am. J. Agric. Environ. Sci. 2012, 12, 1252–1265. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Varshney, P.; Yusuf, M.; Ahmad, A. Polyamines: Potent modulators of plant responses to stress. J. Plant Interact. 2013, 8, 1–16. [Google Scholar] [CrossRef]
- Mustafavi, S.H.; Shekari, F.; Maleki, H.H. Influence of exogenous polyamines on antioxidant defence and essential oil production in valerian (Valeriana officinalis L.) plants under drought stress. Acta Agric. Slov. 2016, 107, 81–91. [Google Scholar] [CrossRef]
- Sequera-mutiozabal, M.; Antoniou, C.; Tiburcio, A.F.; Alcázar, R.; Fotopoulos, V. Polyamines: Emerging hubs promoting drought and salt stress tolerance in plants. Curr. Mol. Biol. Rep. 2017, 3, 28–36. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, J.; Wang, H.; Su, L.; Zhao, C. Biochar addition alleviate the negative effects of drought and salinity stress on soybean productivity and water use efficiency. BMC Plant Biol. 2020, 20, 288. [Google Scholar] [CrossRef]
- Hammad, S.A.R.; Ali, O.A.M. Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agric. Sci. 2014, 59, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Zeid, F.A.; Omer, E.A.; Amin, A.Y.; Hanafy, S.A.H. Effect of putrescine and salicylic acid on Ajwain plant (Trachyspermum ammi) at vegetative stage grown under drought stress. Int. J. Agric. Sci. Res. 2014, 4, 61–80. [Google Scholar]
- Abdel-azem, H.S.; Shehata, S.M.; El-yazied, A.A.; Safia, M. Snap bean response to salicylic acid and putrescine used separately and jointly under two sowing dates. Middle East J. Appl. Sci. 2015, 05, 1211–1221. [Google Scholar]
- Elewa, T.A.; Sadak, M.S.; Saad, A.M. Proline treatment improves physiological responses in quinoa plants under drought stress. Biosci. Res. 2017, 14, 21–33. [Google Scholar]
- Türkan, I.; Bor, M.; Özdemir, F.; Koca, H. Differential responses of lipid peroxidation and antioxidants in the leaves of drought-tolerant P. acutifolius Gray and drought-sensitive P. vulgaris L. subjected to polyethylene glycol mediated water stress. Plant Sci. 2005, 168, 223–231. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Scalschi, L.; Vicedo, B.; Marcos-Barbero, E.L.; Morcuende, R.; Camañes, G. Putrescine: A key metabolite involved in plant development, tolerance and resistance responses to stress. Int. J. Mol. Sci. 2022, 23, 2971. [Google Scholar] [CrossRef]
- Killiny, N.; Nehela, Y. Citrus polyamines: Structure, biosynthesis, and physiological functions. Plants 2020, 9, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Bano, A.; Babar, M.A. Metabolic and physiological changes induced by plant growth regulators and plant growth promoting rhizobacteria and their impact on drought tolerance in Cicer arietinum L. PLoS ONE 2019, 14, e0213040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omer, A.M.; Osman, M.S.; Badawy, A.A. Inoculation with Azospirillum brasilense and/or Pseudomonas geniculata reinforces flax (Linum usitatissimum) growth by improving physiological activities under saline soil conditions. Bot. Stud. 2022, 63, 15. [Google Scholar] [CrossRef]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 2021, 40, 1245–1256. [Google Scholar] [CrossRef]
- Badawy, A.A.; Abdelfattah, N.A.H.; Salem, S.S.; Awad, M.F.; Fouda, A. Efficacy assessment of biosynthesized copper oxide nanoparticles (CuO-NPs) on stored grain insects and their impacts on morphological and physiological traits of wheat (Triticum aestivum L.) plant. Biology 2021, 10, 233. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. Dual application of 24-epibrassinolide and spermine confers drought stress tolerance in maize (Zea mays L.) by modulating polyamine and protein metabolism. J. Plant Growth Regul. 2016, 35, 518–533. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Feiz, F.S.; Hakimi, L.; Mousavi, A.; Jahromi, M.G. The effects of glycine betaine and L-arginine on biochemical properties of pot marigold (Calendula officinalis L.) under water stress. Iran. J. Plant Physiol. 2019, 9, 2795–2805. [Google Scholar]
- Hammad, S.A.R.; El-Gamal, S.M. Response of pepper plants grown under water stress condition to biofertilizers (Halex 2) and mineral nitrogen. Minufiya J. Agric. Res. 2004, 29, 1–27. [Google Scholar]
- Farooq, M.; Wahid, A.; Lee, D.-J. Exogenously applied polyamines increase drought tolerance of rice by improving leaf water status, photosynthesis and membrane properties. Acta Physiol. Plant. 2009, 31, 937–945. [Google Scholar] [CrossRef]
- Amin, M.A.; Ismail, M.A.; Badawy, A.A.; Awad, M.A.; Hamza, M.F.; Awad, M.F.; Fouda, A. The Potency of fungal-fabricated selenium nanoparticles to improve the growth performance of Helianthus annuus L. and control of cutworm Agrotis ipsilon. Catalysts 2021, 11, 1551. [Google Scholar] [CrossRef]
- Badawy, A.A.; Alotaibi, M.O.; Abdelaziz, A.M.; Osman, M.S.; Khalil, A.M.A.; Saleh, A.M.; Mohammed, A.E.; Hashem, A.H. Enhancement of seawater stress tolerance in barley by the endophytic fungus Aspergillus ochraceus. Metabolites 2021, 11, 428. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhong, M.; Shu, S.; Du, N.; Sun, J.; Guo, S. Proteomic and physiological analyses reveal putrescine responses in roots of cucumber stressed by NaCL. Front. Plant Sci. 2016, 7, 1035. [Google Scholar] [CrossRef] [Green Version]
- Hassan, N.; Ebeed, H.; Aljaarany, A. Exogenous application of spermine and putrescine mitigate adversities of drought stress in wheat by protecting membranes and chloroplast ultra-structure. Physiol. Mol. Biol. Plants 2020, 26, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Abdeldaym, E.A.; Sabra, M.A. The root endophytic fungus Piriformospora indica improves growth performance, physiological parameters and yield of tomato under water stress condition. Middle East J. Agric. Res. 2018, 7, 1090–1101. [Google Scholar]
- Raza, M.A.S.; Saleem, M.F.; Shah, G.M.; Khan, I.H.; Raza, A. Exogenous application of glycinebetaine and potassium for improving water relations and grain yield of wheat under drought. J. Soil Sci. Plant Nutr. 2014, 14, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Talaat, N.B.; Shawky, B.T.; Ibrahim, A.S. Alleviation of drought-induced oxidative stress in maize (Zea mays L.) plants by dual application of 24-epibrassinolide and spermine. Environ. Exp. Bot. 2015, 113, 47–58. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, R.; Gheith, E.S.M.S.; Hussain, H.A.; Hussain, S.; Irfan, M.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Effects of salicylic acid, zinc and glycine betaine on morpho-physiological growth and yield of maize under drought stress. Sci. Rep. 2021, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, H.A.M.; Hassanein, R.A.; Khalil, S.I.; El-Khawas, S.A.; El-Bassiouny, H.M.S.; El-Monem, A.A.A. Effect of arginine or putrescine on growth, yield and yield components of late sowing wheat. J. Appl. Sci. Res. 2010, 6, 177–183. [Google Scholar]
- Liu, Y.; Liang, H.; Lv, X.; Liu, D.; Wen, X.; Liao, Y. Effect of polyamines on the grain filling of wheat under drought stress. Plant Physiol. Biochem. 2016, 100, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Alobaidy, M.G. Effect of Putrescine and Humic Acid on Cotton Plant Growing under Salinity Stress Conditions. Master’s Thesis, Faculty of Agriculture, Cairo University, Giza, Egypt, 2013. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Root Length (cm) | Leaves Number | Flag Leaf Area (cm2) | Tillers Number | |
|---|---|---|---|---|---|---|
| Irrigation | Putrescine | |||||
| Control | 0.0 mM | 60 ± 0.58 e | 6.00 ± 0.1 d | 4.67 ± 0.33 de | 14.5 ± 0.35 cde | 2.00 ± 0.1 bc |
| 0.25 mM | 72 ± 1.15 a | 7.33 ± 0.67 cd | 8.33 ± 0.33 abc | 18.1 ± 0.64 bc | 2.33 ± 0.33 bc | |
| 0.50 mM | 74 ± 0.58 a | 8.67 ± 0.33 bc | 9.33 ± 0.33 ab | 19.7 ± 1.11 ab | 3.00 ± 0.1 b | |
| 1.0 mM | 71 ± 0.58 ab | 9.67 ± 0.33 abc | 10.33 ± 0.67 a | 23.3 ± 1.92 a | 4.67 ± 0.33 a | |
| Drought | 0.0 mM | 49 ± 0.58 f | 8.00 ± 0.1 bcd | 3.00 ± 0.1 e | 11.2 ± 0.73 e | 1.67 ± 0.33 c |
| 0.25 mM | 64 ± 0.58 d | 9.00± 0.1 bc | 5.67 ± 0.33 d | 12.8 ± 0.24 de | 2.0 0± 0.1 bc | |
| 0.50 mM | 66 ± 0.58 cd | 10.0 ± 1.0 ab | 6.67 ± 0.33 cd | 15.8 ± 0.61 bcd | 2.67 ± 0.33 bc | |
| 1.0 mM | 68 ± 0.58 bc | 12.0 ± 0.58 a | 8.00 ± 0.57 bc | 19.0 ± 0.13 ab | 3.00 ± 0.1 b | |
| Treatments | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) | |
|---|---|---|---|---|---|
| Irrigation | Putrescine | ||||
| Control | 0.0 mM | 6.3 ± 0.13 cd | 1.51 ± 0.03 cd | 0.82 ± 0.08 bcd | 0.27 ± 0.02 b |
| 0.25 mM | 7.7 ± 0.49 bc | 1.85 ± 0.12 bc | 0.83 ± 0.03 bcd | 0.30 ± 0.02 b | |
| 0.50 mM | 9.3 ± 1.01 ab | 2.22 ± 0.24 ab | 1.03 ± 0.03 ab | 0.42 ± 0.03 a | |
| 1.0 mM | 10.2 ± 0.35 a | 2.45 ± 0.08 a | 1.18 ± 0.06 a | 0.52 ± 0.02 a | |
| Drought | 0.0 mM | 3.4 ± 0.04 e | 0.83 ± 0.01 e | 0.47 ± 0.02 e | 0.14 ± 0.03 c |
| 0.25 mM | 5.2 ± 0.35 de | 1.25 ± 0.09 de | 0.62 ± 0.01 de | 0.23 ± 0.01 bc | |
| 0.50 mM | 6.4 ± 0.11 cd | 1.54 ± 0.03 cd | 0.77 ± 0.03 cd | 0.27 ± 0.03 b | |
| 1.0 mM | 6.9 ± 0.53 cd | 1.65 ± 0.13 cd | 0.94 ± 0.07 bc | ± 0.03 a | |
| No. | MW | L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 180 | + | + | + | + | + | + | + | + |
| 2 | 135 | + | + | + | + | + | + | + | + |
| 3 | 118 | + | + | + | + | + | + | + | + |
| 4 | 110 | - | - | - | - | - | - | + | + |
| 5 | 100 | + | + | + | + | + | + | + | + |
| 6 | 80 | + | + | + | + | + | + | + | + |
| 7 | 70 | + | + | + | + | + | + | + | + |
| 8 | 66 | + | + | + | + | + | + | + | + |
| 9 | 65 | + | + | + | + | + | + | + | + |
| 10 | 64 | - | - | - | - | - | - | + | - |
| 11 | 63 | - | - | + | - | - | + | + | - |
| 12 | 61 | + | + | + | + | + | + | + | + |
| 13 | 60 | + | + | + | + | + | + | + | + |
| 14 | 54 | - | - | - | - | + | - | - | - |
| 15 | 52 | + | + | + | + | - | - | - | - |
| 16 | 48 | + | + | + | + | + | + | + | + |
| 17 | 46 | - | - | + | - | - | + | - | - |
| 18 | 37 | + | + | + | + | + | + | + | + |
| 19 | 36 | - | + | + | + | + | + | + | + |
| 20 | 34 | + | + | + | + | + | + | + | + |
| 21 | 33 | + | + | + | + | + | + | + | + |
| 22 | 32 | - | - | - | - | - | - | + | + |
| 23 | 31 | + | + | + | + | + | + | + | + |
| 24 | 30 | + | + | + | + | + | + | + | + |
| 25 | 28 | + | + | + | + | + | + | + | + |
| 26 | 26 | - | - | - | - | - | - | + | + |
| 27 | 23 | + | + | + | + | + | + | + | + |
| 28 | 21 | + | + | + | + | + | + | + | + |
| 29 | 20 | + | - | - | - | - | - | - | - |
| 30 | 18 | + | + | + | + | + | + | + | + |
| 31 | 17 | + | + | + | + | + | + | + | + |
| 32 | 16 | - | + | + | + | + | + | + | + |
| 33 | 15 | + | + | + | + | + | + | + | + |
| 34 | 12 | + | + | + | + | + | + | + | + |
| Total | 25 | 26 | 28 | 26 | 25 | 27 | 31 | 28 | |
| Treatments | Spike Length | Number of Spikelets | Number of Grains | Weight of Spike (g) | Weight of Grains/Spike (g) | |
|---|---|---|---|---|---|---|
| Irrigation | Putrescine | |||||
| Control | 0.0 mM | 11.67 ± 0.33 a | 10.67 ± 0.33 bc | 25.33 ± 0.67 c | 1.18 ± 0.02 b | 0.68 ± 0.04 cd |
| 0.25 mM | 12.17 ± 0.60 a | 14 ± 0.57 a | 27.67 ± 1.45 bc | 1.57 ± 0.02 a | 0.92 ± 0.10 bc | |
| 0.5 mM | 12.67 ± 0.67 a | 14 ± 1.15 a | 31 ± 0.57 b | 1.69 ± 0.08 a | 1.11 ± 0.07 ab | |
| 1.0 mM | 11 ± 0.57 ab | 13.33 ± 0.33 a | 37.67 ± 1.45 a | 1.72 ± 0.01 a | 1.31 ± 0.10 a | |
| Drought | 0.0 mM | 8.33 ± 0.33 b | 9 ± 0.1 c | 17.67 ± 0.33 d | 0.90 ± 0.01 c | 0.42 ± 0.03 d |
| 0.25 mM | 11.33 ± 0.33 a | 12 ± 0.1 ab | 18 ± 0.57 d | 1.21 ± 0.02 b | 0.58 ± 0.03 d | |
| 0.5 mM | 9.83 ± 0.17 ab | 12 ± 0.1 ab | 26 ± 1 c | 1.24 ± 0.04 b | 0.68 ± 0.03 cd | |
| 1.0 mM | 11 ± 1.15 ab | 14.33 ± 0.33 a | 32 ± 0.57 b | 1.31 ± 0.02 b | 0.63 ± 0.03 cd | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, H.-A.A.; Alshammari, S.O.; Abd El-Sadek, M.E.; Kenawy, S.K.M.; Badawy, A.A. The Promotive Effect of Putrescine on Growth, Biochemical Constituents, and Yield of Wheat (Triticum aestivum L.) Plants under Water Stress. Agriculture 2023, 13, 587. https://doi.org/10.3390/agriculture13030587
Hussein H-AA, Alshammari SO, Abd El-Sadek ME, Kenawy SKM, Badawy AA. The Promotive Effect of Putrescine on Growth, Biochemical Constituents, and Yield of Wheat (Triticum aestivum L.) Plants under Water Stress. Agriculture. 2023; 13(3):587. https://doi.org/10.3390/agriculture13030587
Chicago/Turabian StyleHussein, Hebat-Allah A., Shifaa O. Alshammari, Marwa E. Abd El-Sadek, Sahar K. M. Kenawy, and Ali A. Badawy. 2023. "The Promotive Effect of Putrescine on Growth, Biochemical Constituents, and Yield of Wheat (Triticum aestivum L.) Plants under Water Stress" Agriculture 13, no. 3: 587. https://doi.org/10.3390/agriculture13030587