
Survival and Demography of the Tomato Borer (Tuta absoluta) Exposed to Citrus Essential Oils and Major Compounds †
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Essential Oils
2.3. Chemical Characterization of the EOs
2.4. Acute Toxicity of EOs against T. absoluta
2.5. Time–Concentration–Mortality Responses of T. absoluta to the EOs
2.6. Acute Toxicity of the Major Compounds of the EOs against T. absoluta
2.7. Life History Tables of T. absoluta Treated with LC50 of Citrus spp. EOs
2.8. Statistical Analysis
3. Results
3.1. Chemical Characterization of EOs
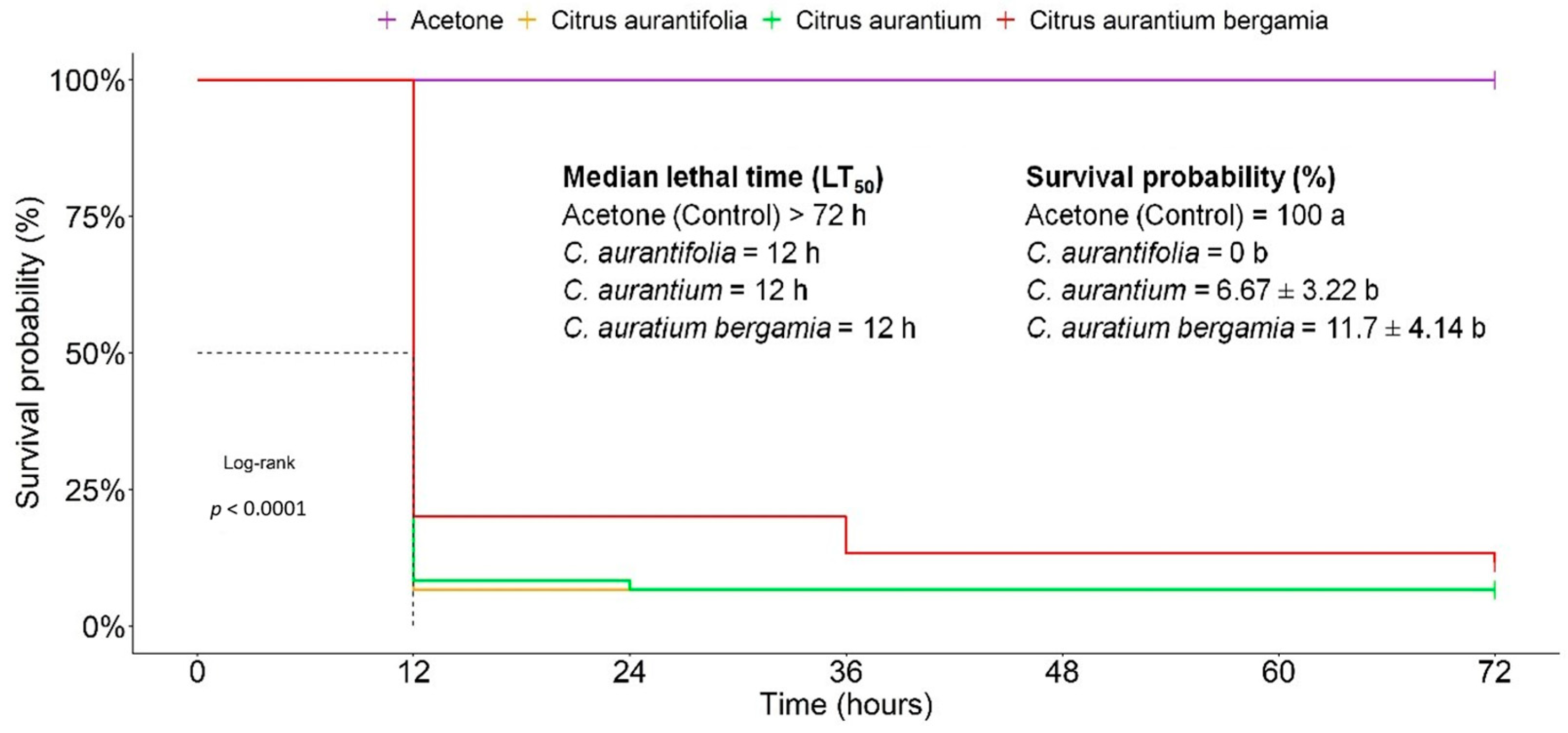
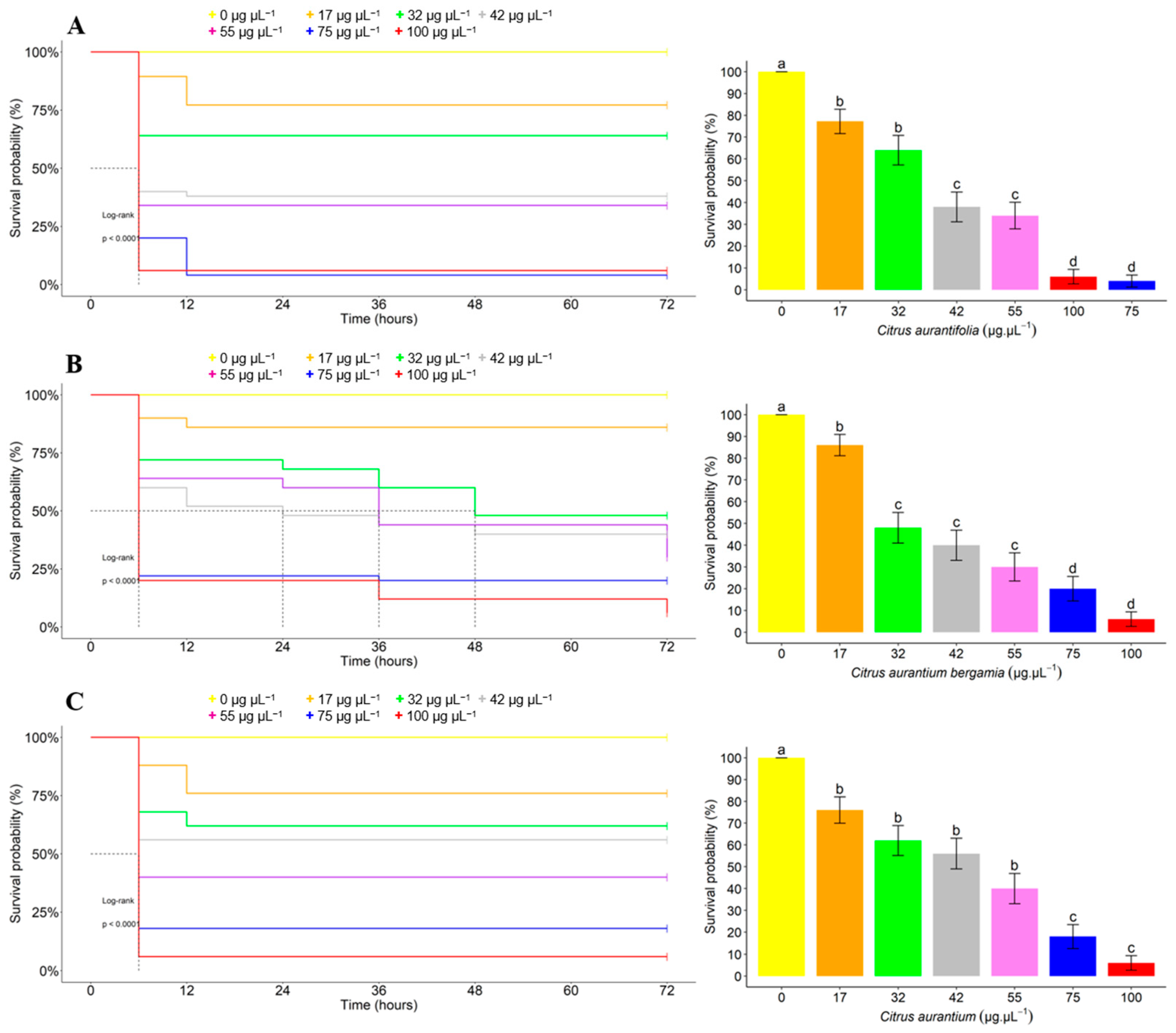
3.2. Acute Toxicity of EOs against T. absoluta
3.3. Time–Concentration–Mortality Responses of T. absoluta to the EOs
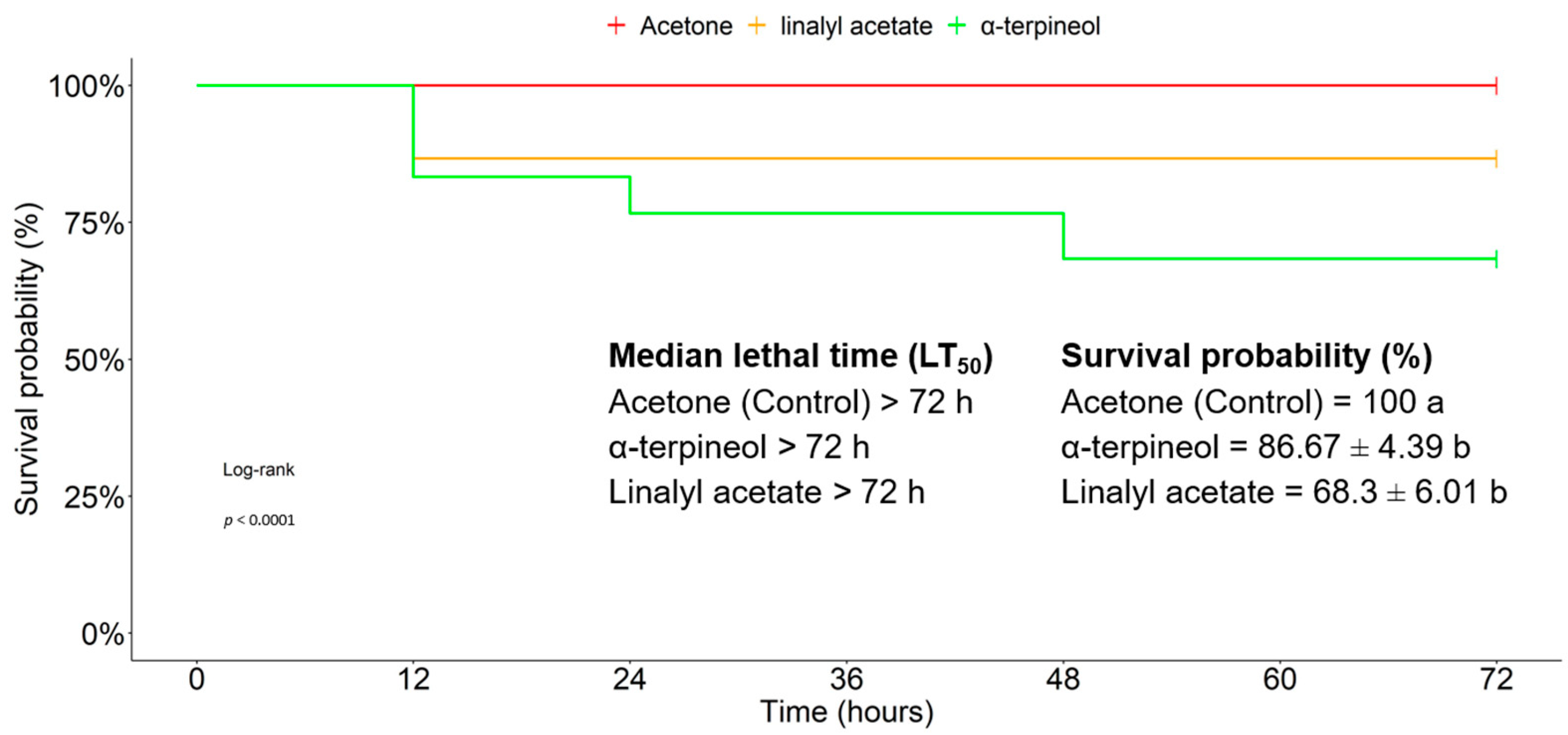
3.4. Acute Toxicity of Major Compounds of EOs against T. absoluta
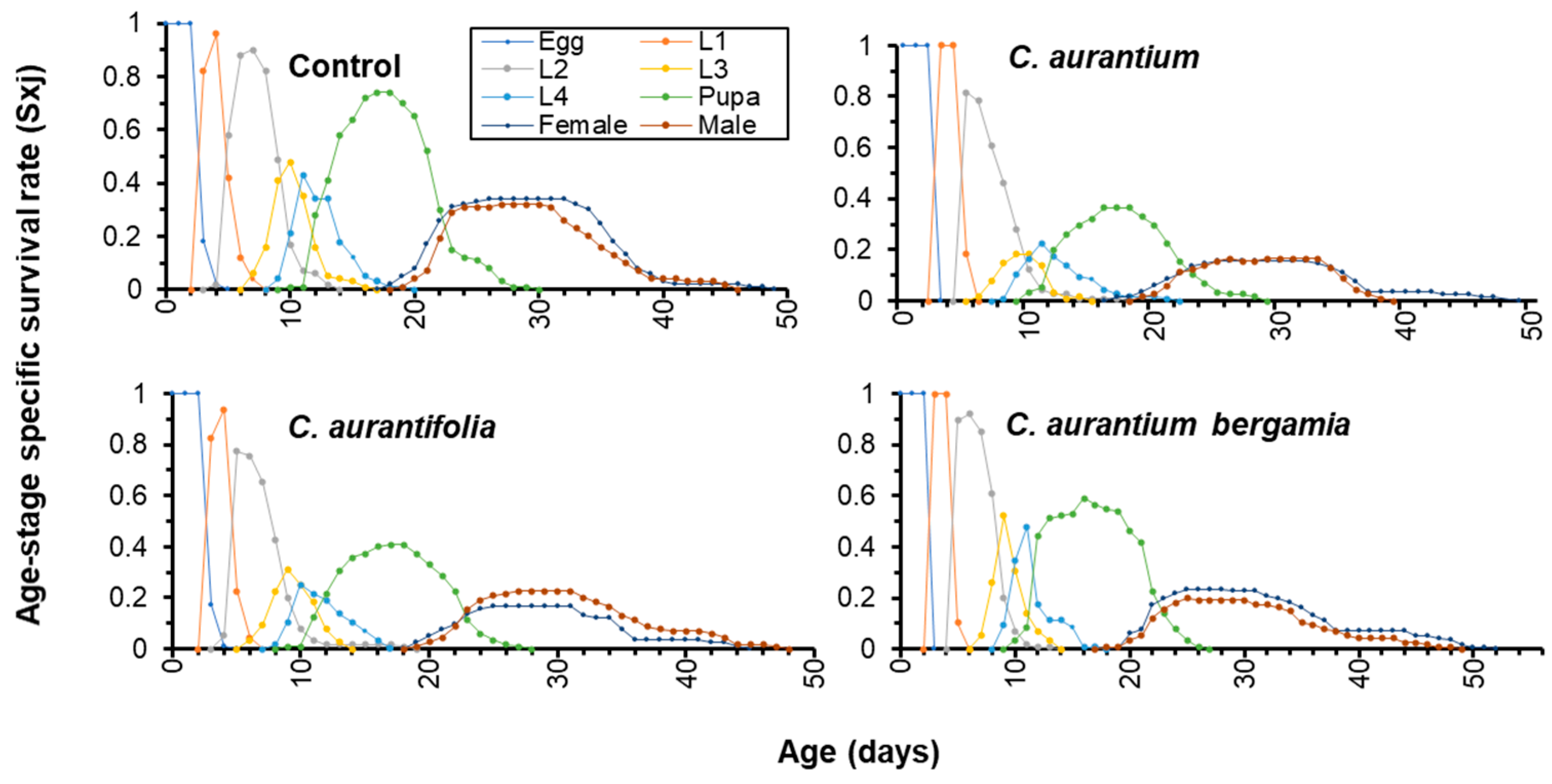
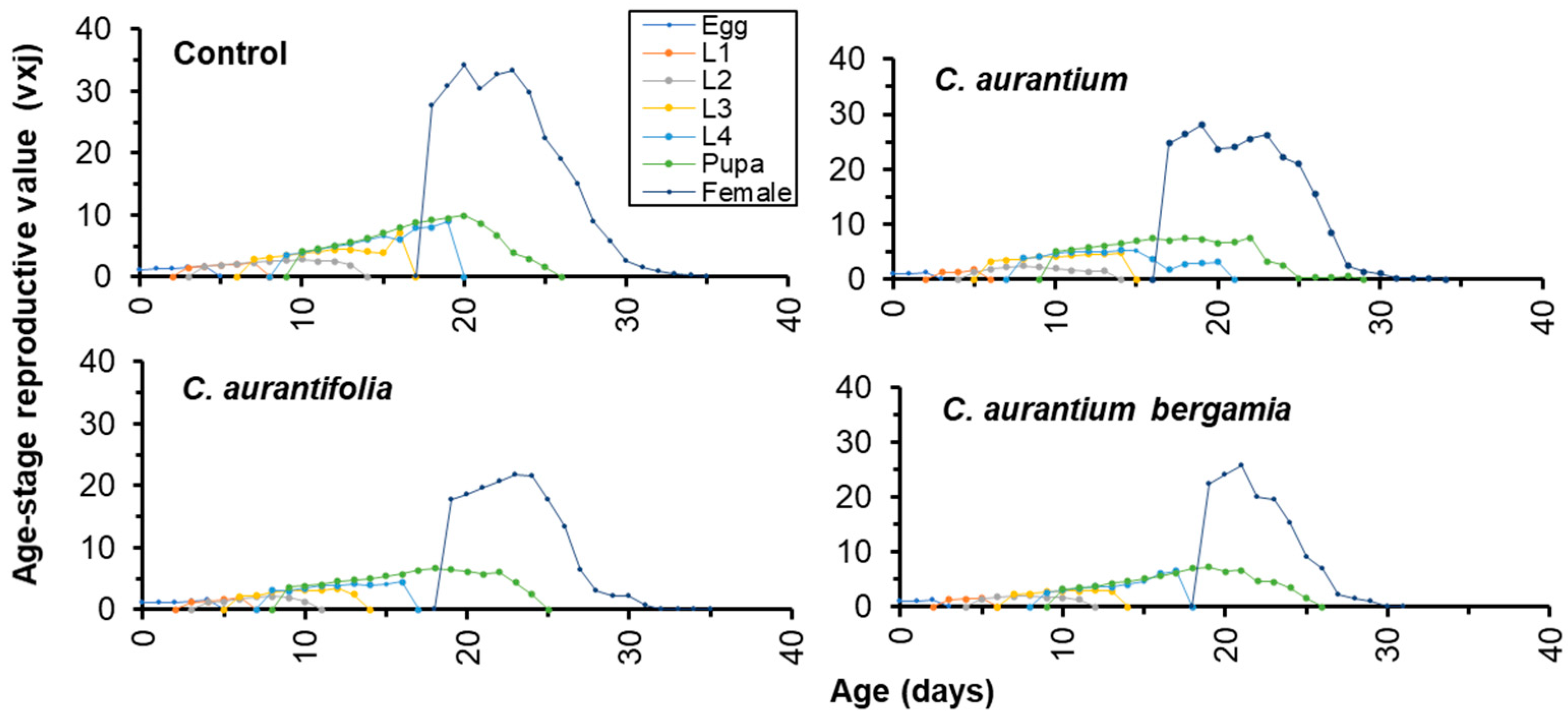
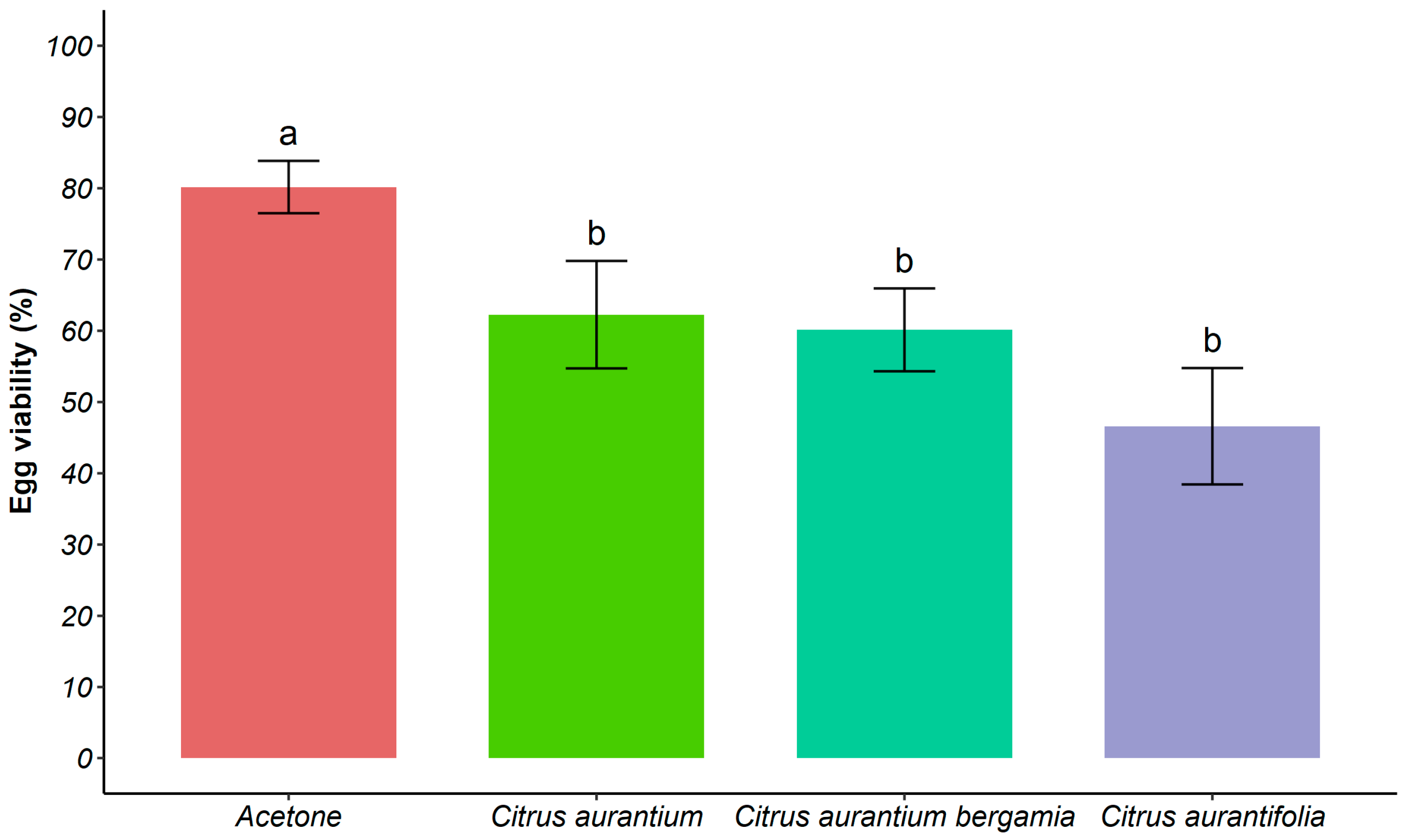
3.5. Life History Tables of T. absoluta Treated with LC50 of Citrus spp. EOs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tropea Garzia, G.; Siscaro, G.; Biondi, A.; Zappalà, L. Tuta absoluta, a South American Pest of Tomato Now in the EPPO Region: Biology, Distribution and Damage. EPPO Bull. 2012, 42, 205–210. [Google Scholar] [CrossRef]
- Biondi, A.; Guedes, R.N.C.; Wan, F.-H.; Desneux, N. Ecology, Worldwide Spread, and Management of the Invasive South American Tomato Pinworm, Tuta absoluta: Past, Present, and Future. Annu. Rev. Entomol. 2018, 63, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological Invasion of European Tomato Crops by Tuta absoluta: Ecology, Geographic Expansion and Prospects for Biological Control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Picanço, M.C. The Tomato Borer Tuta absoluta in South America: Pest Status, Management and Insecticide Resistance. EPPO Bull. 2012, 42, 211–216. [Google Scholar] [CrossRef]
- Han, P.; Bayram, Y.; Shaltiel-Harpaz, L.; Sohrabi, F.; Saji, A.; Esenali, U.T.; Jalilov, A.; Ali, A.; Shashank, P.R.; Ismoilov, K.; et al. Tuta absoluta Continues to Disperse in Asia: Damage, Ongoing Management and Future Challenges. J. Pest Sci. 2019, 92, 1317–1327. [Google Scholar] [CrossRef]
- Zhang, G.-F.; Ma, D.-Y.; Wang, Y.-S.; Gao, Y.-H.; Liu, W.-X.; Zhang, R.; Fu, W.-J.; Xian, X.-Q.; Wang, J.; Kuang, M.; et al. First Report of the South American Tomato Leafminer, Tuta absoluta (Meyrick), in China. J. Integr. Agric. 2020, 19, 1912–1917. [Google Scholar] [CrossRef]
- Santana, P.A.; Kumar, L.; Da Silva, R.S.; Picanço, M.C. Global Geographic Distribution of Tuta absoluta as Affected by Climate Change. J. Pest Sci. 2019, 92, 1373–1385. [Google Scholar] [CrossRef]
- Zhang, G.F.; Ma, D.Y.; Liu, W.X.; Wang, Y.S.; Fu, W.J.; Wang, J.; Gao, Y.H.; Wang, F.H. The Arrival of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). China. J Biosaf. 2019, 28, 200–203. [Google Scholar]
- Campos, M.R.; Biondi, A.; Adiga, A.; Guedes, R.N.C.; Desneux, N. From the Western Palaearctic Region to beyond: Tuta absoluta 10 Years after Invading Europe. J. Pest Sci. 2017, 90, 787–796. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Roditakis, E.; Campos, M.R.; Haddi, K.; Bielza, P.; Siqueira, H.A.A.; Tsagkarakou, A.; Vontas, J.; Nauen, R. Insecticide Resistance in the Tomato Pinworm Tuta absoluta: Patterns, Spread, Mechanisms, Management and Outlook. J. Pest Sci. 2019, 92, 1329–1342. [Google Scholar] [CrossRef]
- Guedes, R. The Tomato Borer Tuta absoluta: Insecticide Resistance and Control Failure. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2012, 7. [Google Scholar] [CrossRef]
- Campos, M.R.; Rodrigues, A.R.S.; Silva, W.M.; Silva, T.B.M.; Silva, V.R.F.; Guedes, R.N.C.; Siqueira, H.A.A. Spinosad and the Tomato Borer Tuta absoluta: A Bioinsecticide, an Invasive Pest Threat, and High Insecticide Resistance. PLoS ONE 2014, 9, e103235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desneux, N.; Han, P.; Mansour, R.; Arnó, J.; Brévault, T.; Campos, M.R.; Chailleux, A.; Guedes, R.N.C.; Karimi, J.; Konan, K.A.J.; et al. Integrated Pest Management of Tuta absoluta: Practical Implementations across Different World Regions. J. Pest Sci. 2022, 95, 17–39. [Google Scholar] [CrossRef]
- Elliott, M.; Farnham, A.W.; Janes, N.F.; Needham, P.H.; Pulman, D.A. Synthetic Insecticide with a New Order of Activity. Nature 1974, 248, 710–711. [Google Scholar] [CrossRef] [PubMed]
- Koul, O.; Walia, S.; Dhaliawal, G. Essential Oils as Green Pesticides: Potential and Constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar]
- Isman, M.B. Commercial Development of Plant Essential Oils and Their Constituents as Active Ingredients in Bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Isman, M.B. Pesticides Based on Plant Essential Oils: Phytochemical and Practical Considerations. ACS Symp. Ser. 2016, 1218, 13–26. [Google Scholar] [CrossRef]
- Sammour, E.; Kandil, M.A.-H.; Abdel-Aziz, N.F.; El Maguied, E.; Agamy, A.M.E.B.; Abdelmaksound, N.M. Field Evaluation of New Formulation Types of Essential Oils against Tuta absoluta and Their Side Effects on Tomato Plants. Acta Scientific Agriculture (ISSN: 2581-365X) Field Evaluation of New Formulation Types of Essential Oils against Tuta absoluta An. Acta Sci. Agric. 2018, 2, 14–22. [Google Scholar]
- Moreno, S.C.; Carvalho, G.A.; Picanço, M.C.; Morais, E.G.F.; Pereira, R.M. Bioactivity of Compounds from Acmella Oleracea against Tuta Absoluta (Meyrick) (Lepidoptera: Gelechiidae) and Selectivity to Two Non-Target Species. Pest Manag. Sci. 2012, 68, 386–393. [Google Scholar] [CrossRef] [Green Version]
- Soares, M.A.; Campos, M.R.; Passos, L.C.; Carvalho, G.A.; Haro, M.M.; Lavoir, A.V.; Biondi, A.; Zappalà, L.; Desneux, N. Botanical Insecticide and Natural Enemies: A Potential Combination for Pest Management against Tuta absoluta. J. Pest Sci. 2019, 92, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhao, J.; Zheng, Y.; Weng, Q.; Biondi, A.; Desneux, N.; Wu, K. Assessment of Potential Sublethal Effects of Various Insecticides on Key Biological Traits of the Tobacco Whitefly, Bemisia Tabaci. Int. J. Biol. Sci. 2013, 9, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brevik, K.; Lindström, L.; McKay, S.D.; Chen, Y.H. Transgenerational Effects of Insecticides—Implications for Rapid Pest Evolution in Agroecosystems. Curr. Opin. Insect Sci. 2018, 26, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, H. Life-Table Analysis Incorporating Both Sexes and Variable Development Rates Among Individuals. Entomol. Ambient. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, Y.H.H.; Huang, Y.; Chi, H. The Age-Stage, Two-Sex Life Table with an Offspring Sex Ratio Dependent on Female Age. J. Agri. Fore. 2011, 60, 337–345. [Google Scholar]
- Chi, H.L.; Chi, H.S.I.N.; Liu, H. Two New Methods for the Study of Insect Population Ecology Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Stark, J.D.; Sugayama, R.L.; Kovaleski, A. Why Demographic and Modeling Approaches Should Be Adopted for Estimating the Effects of Pesticides on Biocontrol Agents. BioControl 2007, 52, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and Damage Projection of Spodoptera litura (F.) on Peanuts (Arachishypogaea L.) under Different Conditions Using the Age-Stage, Two-Sex Life Table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatograpy/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; pp. 804–806. [Google Scholar]
- Genç, H. The Tomato Leafminer, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae): Pupal Key Characters for Sexing Individuals. Turkish J. Zool. 2016, 40, 801–805. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2015. Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 3 April 2020).
- Efron, B.; Tibshirani, R.J. Volunteer Motivation, Satisfaction, and Management at an Elite Sporting Competition. In An Introduction to the Bootstrap; Farrell, J., Johnston, M., Twynam, D., Eds.; Chapman and Hall: New York, NY, USA, 1998. [Google Scholar]
- Lumley, T.; Elizabeth, A.; Cynthia, C.; Therneau, M.T.M. Package Survival. 2020. Available online: https://cran.r-project.org/web/packages/survival/index.html (accessed on 3 April 2020).
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing 2020. Available online: https://cran.r-project.org/bin/windows/base/ (accessed on 3 April 2020).
- Ritz, C.; Strebig, J.C. Package “Drc”: Analysis of Dose-Response Curves. 2016. Available online: https://cran.r-project.org/web/packages/drc/drc.pdf (accessed on 3 April 2020).
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biometrical. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacoli, F.; Bell, V.A.; Cargnus, E.; Pavan, F. Insecticidal Activity of Natural Products against Vineyard Mealybugs (Hemiptera: Pseudococcidae). Crop. Prot. 2018, 111, 50–57. [Google Scholar] [CrossRef]
- Campolo, O.; Puglisi, I.; Barbagallo, R.N.; Cherif, A.; Ricupero, M.; Biondi, A.; Palmeri, V.; Baglieri, A.; Zappalà, L. Side Effects of Two Citrus Essential Oil Formulations on a Generalist Insect Predator, Plant and Soil Enzymatic Activities. Chemosphere 2020, 257, 127252. [Google Scholar] [CrossRef] [PubMed]
- Kabiri, M.; Abad, R.; Besheli, B.A. Insecticidal Potential of Essential Oil from the Leaves of Citrus aurantium L. against Oryzaephilus surinamensis (F.), Lasioderma serricorne (L.) and Sitophilus oryzae (L.). J. Entomol. Zool. Stud. 2016, 4, 865–869. [Google Scholar]
- Oboh, G.; Ademosun, A.O.; Olumuyiwa, T.A.; Olasehinde, T.A.; Ademiluyi, A.O.; Adeyemo, A.C. Insecticidal Activity of Essential Oil from Orange Peels (Citrus sinensis) against Tribolium confusum, Callosobruchus maculatus and Sitophilus oryzae and Its Inhibitory Effects on Acetylcholinesterase and Na+/K+-ATPase Activities. Phytoparasitica 2017, 45, 501–508. [Google Scholar] [CrossRef]
- López, M.D.; Pascual-Villalobos, M.J. Mode of Inhibition of Acetylcholinesterase by Monoterpenoids and Implications for Pest Control. Ind. Crops Prod. 2010, 31, 284–288. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential Oils in Insect Control: Low-Risk Products in a High-Stakes World. Annu. Rev. Entomol. 2011, 57, 405–424. [Google Scholar] [CrossRef]
- Liu, X.C.; Li, Y.P.; Li, H.Q.; Deng, Z.W.; Zhou, L.; Liu, Z.L.; Du, S.S. Identification of Repellent and Insecticidal Constituents of the Essential Oil of Artemisia Rupestris L. Aerial Parts against Liposcelis Bostrychophila Badonnel. Molecules 2013, 18, 10733–10746. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Wang, W.; Dai, J.; Zhu, L. Chemical Composition and Biological Activity against Tribolium Castaneum (Coleoptera: Tenebrionidae) of Artemisia Brachyloba Essential Oil. Ind. Crops Prod. 2019, 128, 29–37. [Google Scholar] [CrossRef]
- Calandra, M.J.; Wang, Y. An HPLC Method for the Detection of Hydro Peroxides Derived from Linalyl Acetate in Citrus Oils, Using Post-Column Luminol-Mediated Chemiluminescence Detection. Flavour Fragr. J. 2017, 32, 178–183. [Google Scholar] [CrossRef]
- Soares Rodrigues, G.C.; Dos Santos Maia, M.; Muratov, E.N.; Scotti, L.; Scotti, M.T. Quantitative Structure-Activity Relationship Modeling and Docking of Monoterpenes with Insecticidal Activity against Reticulitermes chinensis Snyder and Drosophila melanogaster. J. Agric. Food Chem. 2020, 68, 4687–4698. [Google Scholar] [CrossRef] [PubMed]
- de Elguea-Culebras, G.O.; Sánchez-Vioque, R.; Berruga, M.I.; Herraiz-Peñalver, D.; Santana-Méridas, O. Antifeedant Effects of Common Terpenes from Mediterranean Aromatic Plants on Leptinotarsa decemlineata. J. Soil Sci. Plant Nutr. 2017, 17, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Campolo, O.; Malacrinò, A.; Zappalà, L.; Laudani, F.; Chiera, E.; Serra, D.; Russo, M.; Palmeri, V. Fumigant Bioactivity of Five Citrus Essential Oils against Tribolium confusum. Phytoparasitica 2014, 42, 223–233. [Google Scholar] [CrossRef]
- Zarrad, K.; Ben Hamouda, A.; Bouslama, T.; Laarif, A. Chemical Composition and Insecticidal Effects of Citrus Aurantium of Essential Oil and Its Powdery Formulation against Tuta absoluta. Tunis. J. Plant Prot. 2017, 12, 83–94. [Google Scholar]
- Tchameni, S.N.; Mbiakeu, S.N.; Sameza, M.L.; Jazet, P.M.D.; Tchoumbougnang, F. Using Citrus Aurantifolia Essential Oil for the Potential Biocontrol of Colocasia esculenta (Taro) Leaf Blight Caused by Phytophthora colocasiae. Environ. Sci. Pollut. Res. 2018, 25, 29929–29935. [Google Scholar] [CrossRef]
- Werdin González, J.O.; Gutiérrez, M.M.; Ferrero, A.A.; Fernández Band, B. Essential Oils Nanoformulations for Stored-Product Pest Control-Characterization and Biological Properties. Chemosphere 2014, 100, 130–138. [Google Scholar] [CrossRef]
- Tadeo, F.R.; Cercós, M.; Colmenero-Flores, J.M.; Iglesias, D.J.; Naranjo, M.A.; Ríos, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; et al. Molecular Physiology of Development and Quality of Citrus. Adv. Bot. Res. 2008, 47, 147–223. [Google Scholar] [CrossRef]
- Kesterson, J.W.; Attaway, J.A.; Wolford, R.W. Physicochemical Properties of Citrus Essential Oils from Florida. J. Agric. Food Chem. 1971, 19, 1097–1105. [Google Scholar] [CrossRef]
- Isman, M.B.; Grieneisen, M.L. Botanical Insecticide Research: Many Publications, Limited Useful Data. Trends Plant Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Karpouhtsis, I.; Pardali, E.; Feggou, E.; Kokkini, S.; Scouras, Z.G.; Mavragani-Tsipidou, P. Insecticidal and Genotoxic Activities of Oregano Essential Oils. J. Agric. Food Chem. 1998, 46, 1111–1115. [Google Scholar] [CrossRef]
- Cal, K. Skin Penetration of Terpenes from Essential Oils and Topical Vehicles. Planta Med. 2006, 72, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.B.; Philogfene, B.J.R. Invited Review: Insecticide Synergists: Role, Importance, and Perspectives. J. Toxicol. Environ. Health 1993, 38, 199–233. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, R.L. Mode of Action of Insecticide Synergists. Annu. Rev. Entomol. 1967, 12, 229–256. [Google Scholar] [CrossRef]
- Richards, O.W. The Theoretical and Practical Study of Natural Insect Populations. Annu. Rev. Entomol. 1961, 6, 147–162. [Google Scholar] [CrossRef]
- Goundoudaki, S.; Tsitsipis, J.A.; Margaritopoulos, J.T.; Zarpas, K.D.; Divanidis, S. Performance of the Tobacco Aphid Myzus persicae (Hemiptera: Aphididae) on Oriental and Virginia Tobacco Varieties. Agric. For. Entomol. 2003, 5, 285–291. [Google Scholar] [CrossRef]
- Stark, J.D.; Banks, J.E. Population-Level Effects of Pesticides and Other Toxicants on Arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef]
- Kandil, M.A.; Abdel-kerim, R.N.; Moustafa, M.A.M. Lethal and Sub-Lethal Effects of Bio-and Chemical Insecticides on the Tomato Leaf Miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Egypt. J. Biol. Pest Control. 2020, 30, 76. [Google Scholar] [CrossRef]
- Bariola, L.A. Pink Bollworms (Lepidoptera: Gelechiidae): Effects of Low Concentrations of Selected Insecticides on Mating and Fecundity in the Laboratory1. J. Econ. Entomol. 1984, 77, 1278–1282. [Google Scholar] [CrossRef]
- Beach, M.R.; Todd, J.W. Toxicity of Avermectin to Larva and Adult Soybean Looper (Lepidoptera: Noctuidae) and Influence on Larva Feeding and Adult Fertility and Fecundity. J. Econ. Entomol. 1985, 78, 1125–1128. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil | RI a | Compound | Percentages | Method of Identification b |
|---|---|---|---|---|
| C. aurantifolia | 921 | tert-Butylbenzene | 2.25 | RI, GC-MS |
| 925 | Limonene | 11.88 | RI, GC-MS | |
| 1010 | α-Fenchol | 2.41 | RI, GC-MS | |
| 1030 | 3-terpinen-1-ol | 4.5 | RI, GC-MS | |
| 1040 | β-Terpineol | 3.18 | RI, GC-MS | |
| 1061 | 2,3,3-Trimethyl-1,4-pentadieno | 2.2 | RI, GC-MS | |
| 1087 | α-Terpinol * | 44.74 | RI, GC-MS | |
| 1094 | CTK1F4019 (C10H16) | 3.88 | RI, GC-MS | |
| 1404 | CTK5J8343 (C12H20) | 2.14 | RI, GC-MS | |
| Unknown compounds | 22.82 | |||
| C. aurantium | 998 | Linalool | 28.04 | RI, GC-MS |
| 1087 | α-Terpineol | 6.22 | RI, GC-MS | |
| 1126 | Nerol | 0.71 | RI, GC-MS | |
| 1154 | Linalyl acetate * | 55.45 | RI, GC-MS | |
| 1263 | Neryl acetate | 6.93 | RI, GC-MS | |
| Unknown compounds | 2.65 | |||
| C. aurantiumbergamia | 925 | Limonene | 4.99 | RI, GC-MS |
| 998 | Linalool | 29.94 | RI, GC-MS | |
| 1153 | Linalyl acetate * | 58.12 | RI, GC-MS | |
| Unknown compounds | 6.95 |
| Essential Oil | df | χ2 | p | * LC50 | CL 95% | * LC90 | CL 95% | b ** ± SE |
|---|---|---|---|---|---|---|---|---|
| C. aurantifolia | 10 | 12.29 | 0.27 | 33.75 | 29.31–38.18 | 88.59 | 68.77–108.42 | 2.28 ± 0.28 |
| C. aurantium | 10 | 10.07 | 0.43 | 38.78 | 33.62–43.94 | 113.36 | 82.41–144.31 | 2.05 ± 0.27 |
| C. aurantium bergamia | 10 | 4.16 | 0.94 | 35.05 | 30.43–39.68 | 94.89 | 72.46–117.33 | 2.21 ± 0.28 |
| Parameter | Control (Acetone) | Citrus aurantifolia | Citrus aurantium | Citrus aurantium bergamia | ||||
|---|---|---|---|---|---|---|---|---|
| N | Mean ± SE | N | Mean ± SE | N | Mean ± SE | N | Mean ± SE | |
| Fecundity (E/F) total | 34 | 46.82 ± 5.76 a | 19 | 24.79 ± 4.05 b | 19 | 30.63 ± 6.1 ab | 30 | 23.1 ± 4.81 b |
| Fecundity (E/F) * | 30 | 53.07 ± 5.61 a | 16 | 29.00 ± 3.77 b | 16 | 36.38 ± 6.26 b | 18 | 38.5 ± 5.57 ab |
| Oviposition (days) | 30 | 3.43 ± 0.30 a | 16 | 2.56 ± 0.26 b | 16 | 2.81 ± 0.25 ab | 18 | 2.28 ± 0.24 b |
| PPOA (days) | 30 | 3.20 ± 0.32 a | 16 | 3.5 ± 0.52 a | 16 | 3.38 ± 0.42 a | 18 | 2.28 ± 0.4 a |
| PPOT (days) | 30 | 24.67 ± 0.37 a | 16 | 25.38 ± 0.46 a | 16 | 24.88 ± 0.54 a | 18 | 24.33 ± 0.47 a |
| MFD (E/F) | 70 | 30 | 46 | 71 | ||||
| MFT (E/F) | 126 | 54 | 85 | 93 | ||||
| Parameter | Control | Citrus aurantifolia | Citrus aurantium | Citrus aurantium bergamia |
|---|---|---|---|---|
| intrinsic rate of increase (r) | 0.11 ± 0.01 a | 0.05 ± 0.01 b | 0.06 ± 0.01 b | 0.07 ± 0.01 b |
| finite rate of increase (λ) | 1.11 ± 0.01 a | 1.05 ± 0.01 b | 1.06 ± 0.01 b | 1.07 ± 0.01 b |
| net reproductive rate (R0) | 15.92 ± 2.94 a | 4.10 ± 1.07 b | 5.06 ± 1.45 b | 6.03 ± 1.56 b |
| mean generation time (T) | 26.18 ± 0.48 ab | 26.83 ± 0.47 a | 26.09 ± 0.62 ab | 25.22 ± 0.47 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Paiva Silva, G.T.; Figueiredo, K.G.; Alves, D.S.; de Oliveira, D.F.; Silva, G.H.; de Souza e Silva, G.T.; de Oliveira, M.S.; Biondi, A.; Carvalho, G.A.
Survival and Demography of the Tomato Borer (Tuta absoluta) Exposed to Citrus Essential Oils and Major Compounds
de Paiva Silva GT, Figueiredo KG, Alves DS, de Oliveira DF, Silva GH, de Souza e Silva GT, de Oliveira MS, Biondi A, Carvalho GA.
Survival and Demography of the Tomato Borer (Tuta absoluta) Exposed to Citrus Essential Oils and Major Compounds
de Paiva Silva, Gabriel Tadeu, Karolina Gomes Figueiredo, Dejane Santos Alves, Denilson Ferreira de Oliveira, Geraldo Humberto Silva, Gabriela Trindade de Souza e Silva, Murilo Silva de Oliveira, Antonio Biondi, and Geraldo Andrade Carvalho.
2023. "Survival and Demography of the Tomato Borer (Tuta absoluta) Exposed to Citrus Essential Oils and Major Compounds