
Effect of Plant Seed Mixture on Overwintering and Floristic Attractiveness of the Flower Strip in Western Poland



Abstract
:1. Introduction
1.1. Environmental Multifunctionality of Flower Strips
1.2. Basics of Flower Strip Establishment
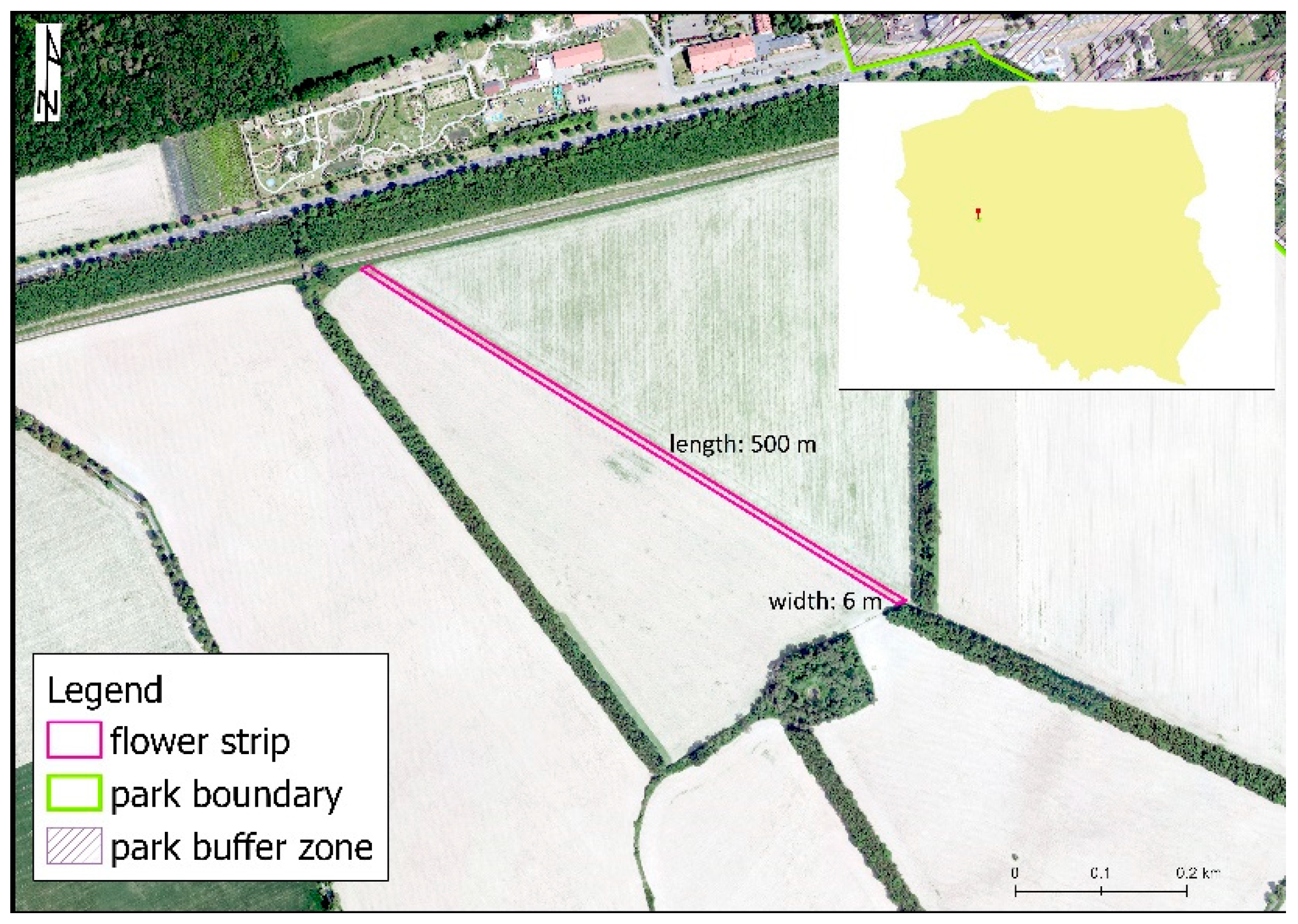
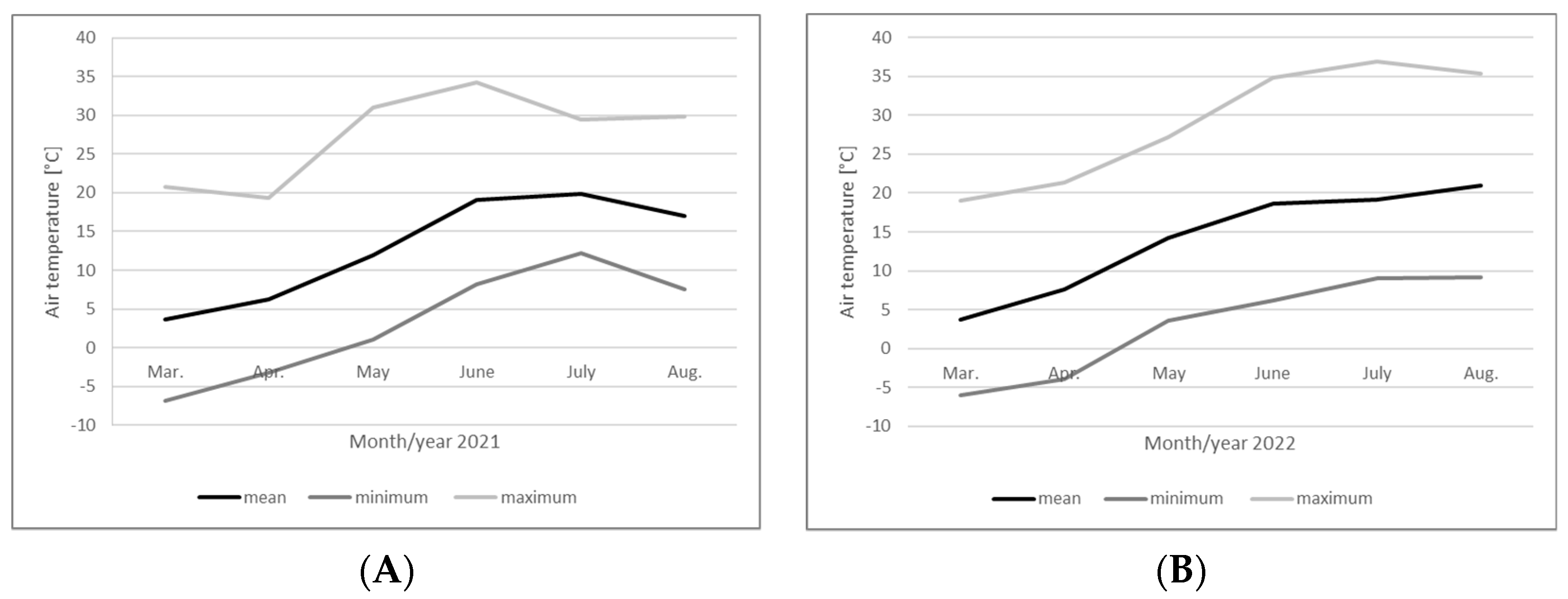
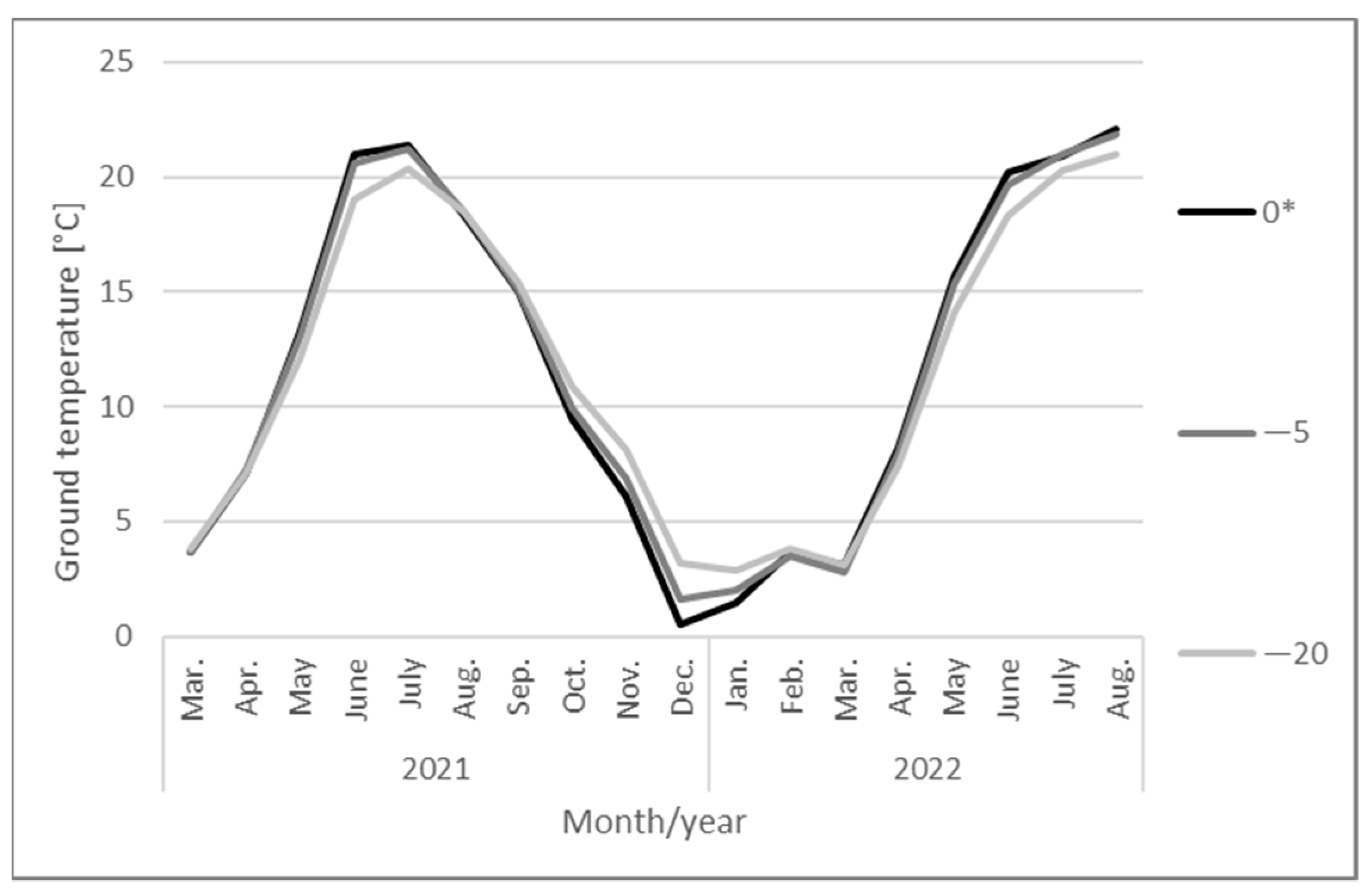
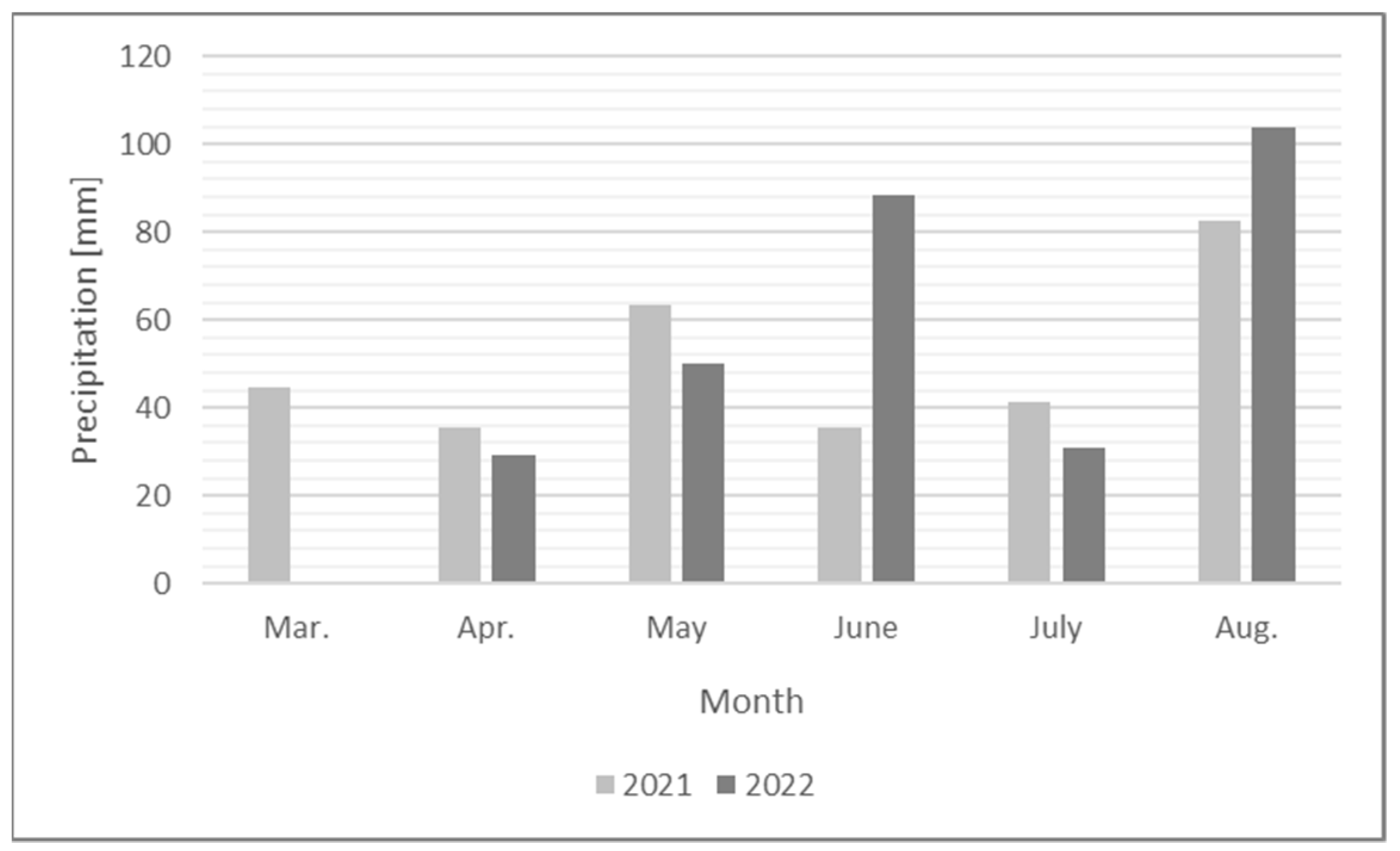
2. Materials and Methods
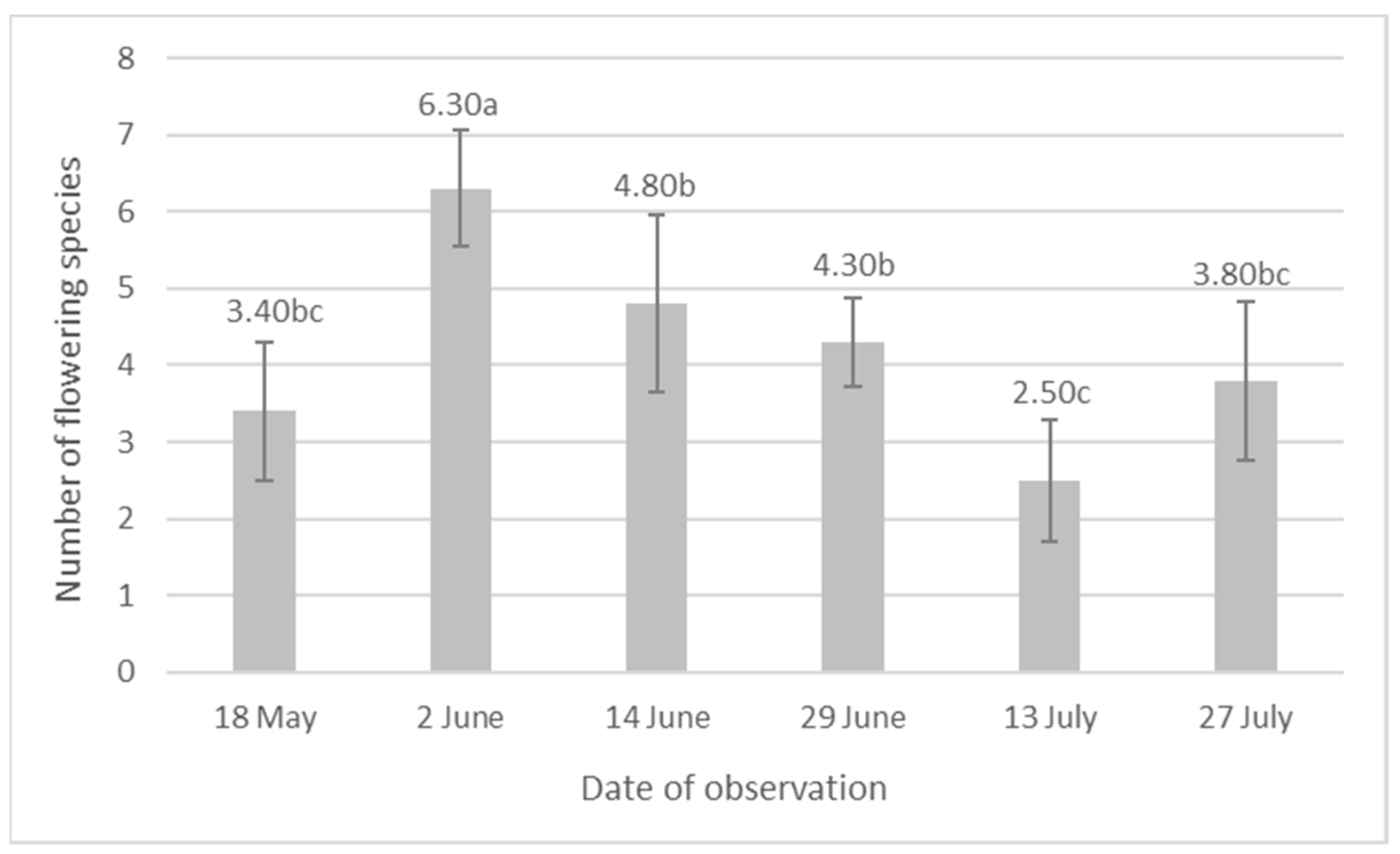
3. Results
4. Discussion
5. Conclusions
- The selection of the appropriate species composition of mixtures intended for flower strips should take into account not only the preferences of beneficial insects but also environmental conditions, which, to a large extent, determine the success of their cultivation. The possibility of their overwintering is important.
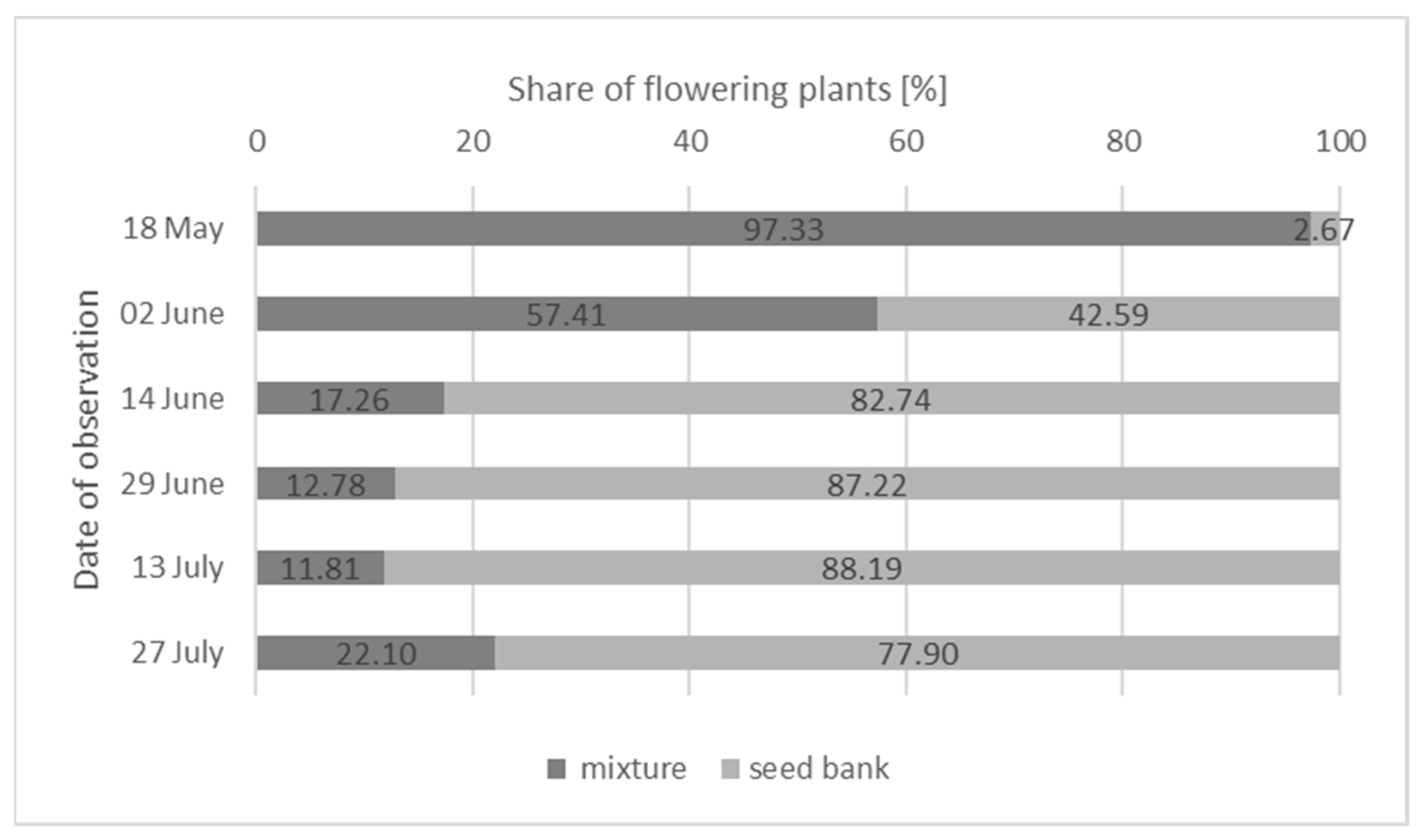
- From the beginning of June to the end of July, the share of flowering plants from the seed bank ranged from 42.59% to 88.19%, while among originally intended plant species, it was only 11.81–57.41%. In May and at the beginning of June, two intended species that were intensively flowering, T. incarnatum with dark-pink inflorescences (in May 70.5%) and P. tanacetifolia with a violet color (at the beginning of June 33.10%), were definitely dominant and are recommended for sowing.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Carvalho, C.R.; de Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Huang, J.; Xu, C.-C.; Ridoutt, B.G.; Wang, X.-C.; Ren, P.-A. Nitrogen and phosphorus losses and eutrophication potential associated with fertilizer application to cropland in China. J. Clean. Product. 2017, 159, 171–179. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Bennekou, S.H. Moving towards a holistic approach for human health risk assessment—Is the current approach fit for purpose? EFSA J. 2019, 17, e170711. [Google Scholar] [CrossRef] [PubMed]
- Kuś, J.; Jończyk, K. Uprawa zbóż w gospodarstwach ekologicznych. Wieś Jutra 2005, 04, 34–36. [Google Scholar]
- Kujawa, K.; Bernacki, Z.; Arczyńska-Chudy, E.; Janku, K.; Karg, J.; Kowalska, J.; Oleszczuk, M.; Sienkiewicz, P.; Sobczyk, D.; Weyssenhoff, D. Kwietne pasy: Rzadko stosowane w Polsce narzędzie wzmacniania integrowanej ochrony roślin uprawnych oraz zwiększania różnorodności biologicznej na terenach rolniczych. Prog. Plant Prot. 2018, 58, 115–128. [Google Scholar]
- Wyss, E.; Pfiffner, L. Biodiversity in Organic Horticulture—An Indicator for Sustainability and a Tool for Pest Management. Acta Hort. 2008, 767, 75–80. [Google Scholar] [CrossRef]
- Fountain, M.T. Impacts of Wildflower Interventions on Beneficial Insects in Fruit Crops: A Review. Insects 2022, 13, 304. [Google Scholar] [CrossRef]
- Buhk, C.; Oppermann, R.; Schanowski, A.; Schanowski, A.; Bleil, R.; Ludemann, J.; Maus, C. Flower strip networks offer promising long term effects on pollinator species richness in intensively cultivated agricultural areas. BMC Ecol. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Pfiffner, L.; Wyss, E. Use of sown wildflower strips to enhance natural enemies of agricultural pests. In Ecological Engineering for Pest Management: Advances in Habitat Manipulation for Arthropods; Gurr, G.M., Wratten, S.D., Altieri, M.A., Eds.; CSIRO Publishing: Collingwood, Australia, 2004; pp. 165–186. [Google Scholar]
- Uyttenbroeck, R.; Hatt, S.; Piqueray, J.; Paul, A.; Bodson, B.; Francis, F.; Monty, A. Creating Perennial Flower Strips: Think Functional! Agric. Agric. Sci. Procedia 2015, 6, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Tschumi, M.; Albrecht, M.; Collatz, J.; Dubsky, V.; Entling, M.H.; Najar-Rodriguez, A.J.; Jacot, K. Tailored flower strips promote natural enemy biodiversity and pest control in potato crops. J. Appl. Ecol. 2016, 53, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- Uyttenbroeck, R.; Hatt, S.; Paul, A.; Boeraeve, B.; Piqueray, J.; Francis, F.; Danthine, S.; Frederich, M.; Dufrêne, M.; Bodson, B.; et al. Pros and cons of flowers strips for farmers. A review. Biotechnol. Agron. Soc. Environ. 2016, 20, 225–235. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2013, 33, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Amy, C.; Noël, G.; Hatt, S.; Uyttenbroeck, R.; Van de Meutter, F.; Genoud, D.; Francis, F. Flower Strips in Wheat Intercropping System: Effect on Pollinator Abundance and Diversity in Belgium. Insects 2018, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Rader, R.; Cunningham, S.A.; Howlett, B.G.; Inouye, D.W. Non-Bee Insects as Visitors and Pollinators of Crops: Biology, Ecology, and Management. Annu. Rev. Entomol. 2020, 7, 391–407. [Google Scholar] [CrossRef] [Green Version]
- Schoch, K.; Tschumi, M.; Lutter, S.; Ramseier, H.; Zingg, S. Competition and Facilitation Effects of Semi-Natural Habitats Drive Total Insect and Pollinator Abundance in Flower Strips. Front. Ecol. Evol. 2022, 10, 854058. [Google Scholar] [CrossRef]
- Wix, N.; Reich, M.; Schaarschmidt, F. Butterfly richness and abundance in flower strips and field margins: The role of local habitat quality and landscape context. Heliyon 2019, 5, e01636. [Google Scholar] [CrossRef] [Green Version]
- Tschumi, M.; Albrecht, M.; Entling, M.H.; Jacot, K. High effectiveness of tailored flower strips in reducing pests and crop plant damage. Proc. R. Soc. B 2015, 282, 20151369. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.K.; Sørensen, H.; Sigsgaard, L. Perennial flower strips in apple orchards promote natural enemies in their proximity. Crop Prot. 2022, 156, 105962. [Google Scholar] [CrossRef]
- Herrera, R.A.; Cotes, B.; Agustí, N.; Tasin, M.; Porcel, M. Using flower strips to promote green lacewings to control cabbage insect pests. J. Pest Sci. 2022, 95, 669–683. [Google Scholar] [CrossRef]
- Serée, L.; Chiron, F.; Valantin-Morison, M.; Barbottin, A.; Gardarin, A. Flower strips, crop management and landscape composition effects on two aphid species and their natural enemies in faba bean. Agric. Ecosyst. Environ. 2022, 331, 107902. [Google Scholar] [CrossRef]
- Baggen, L.R.; Gurr, G.M.; Meats, A. Flowers in tri-trophic systems: Mechanisms allowing selective exploitationby insect natural enemies for conservation biological control. Entomol. Exp. Appl. 1999, 91, 155–161. [Google Scholar] [CrossRef]
- Agro Profil. Magazyn Rolniczy. Available online: https://agroprofil.pl/wiadomosci/jakwplywana-uprawe-maja-pasy-kwietnedane-zaskakuja (accessed on 11 May 2022).
- Barbir, J.; Azpiazu, C.; Badenes-Pérez, F.R.; Fernandez-Quintanilla, C.; Dorado, J. Functionality of Selected Aromatic Lamiaceae in Attracting Pollinators in Central Spain. J. Econ. Entomol. 2016, 109, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.-C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest. Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Nicholson, C.C.; Ricketts, T.H.; Koh, I.; Smith, H.G.; Lonsdorf, E.V.; Olsson, O. Flowering resources distract pollinators from crops: Model predictions from landscape simulations. J. Appl. Ecol. 2019, 56, 618–628. [Google Scholar] [CrossRef]
- Pontin, D.; Wade, M.; Kehrli, P.; Wratten, S. Attractiveness of single and multiple species flower patches to beneficial insects in agroecosystems. Ann. Appl. Biol. 2006, 148, 39–47. [Google Scholar] [CrossRef]
- Jones, J.; Rader, R. Pollinator nutrition and its role in merging the dual objectives of pollinator health and optimal crop production. Phil. Trans. R. Soc. 2022, 377, 20210170. [Google Scholar] [CrossRef]
- Baden-Böhm, F.; Thiele, J.; Dauber, J. Response of honeybee colony size to flower strips in agricultural landscapes depends on areal proportion, spatial distribution and plant composition. Basic Appl. Ecol. 2022, 60, 123–138. [Google Scholar] [CrossRef]
- Ouvrard, P.; Transon, J.; Jacquemart, A.L. Flower-strip agri-environment schemes provide diverse and valuable summer flower resources for pollinating insects. Biodivers. Conserv. 2018, 27, 2193–2216. [Google Scholar] [CrossRef]
- Kujawa, K.; Bernacki, Z.; Kowalska, J.; Kujawa, A.; Oleszczuk, M.; Sienkiewicz, P.; Sobczyk, D. Annual Wildflower Strips as a Tool for Enhancing Functional Biodiversity in Rye Fields in an Organic Cultivation System. Agronomy 2020, 10, 1696. [Google Scholar] [CrossRef]
- Haenke, S.; Scheid, B.; Schaefer, M.; Tscharntke, T. Increasing syrphid fly diversity and density in sown flower strips within simple vs. complex landscapes. J. Appl. Ecol. 2009, 46, 1106–1114. [Google Scholar] [CrossRef]
- Klatt, B.K.; Nilsson, L.; Smith, H.G. Annual flowers strips benefit bumble bee colony growth and reproduction. Biol. Conserv. 2020, 252, 108814. [Google Scholar] [CrossRef]
- Raderschall, C.A.; Lundin, O.; Lindström, S.A.M.; Bommarco, R. Annual flower strips and honeybee hive supplementation differently affect arthropod guilds and ecosystem services in a mass-flowering crop. Agric. Ecosyst. Environ. 2022, 326, 107754. [Google Scholar] [CrossRef]
- Ganser, D.; Knop, E.; Albrecht, M. Sown wildflower strips as overwintering habitat for arthropods: Effective measure or ecological trap? Agric. Ecosyst. Environ. 2019, 275, 123–131. [Google Scholar] [CrossRef]
- Corbet, S.A. Insects, plants and succession: Advantages of long-term set-aside. Agric. Ecosyst. Environ. 1995, 53, 201–217. [Google Scholar] [CrossRef]
- Ullrich, K. The Influence of Wildflower Strips on Plant and Insect (Hetroptera) Diversity in Anarable Landscape. Ph.D. Thesis, Swiss Federal Institute of Technology (ETH), Zürich, Switzerland, 2001. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2021, 23, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Rollin, O.; Bretagnolle, V.; Decourtye, A.; Aptel, J.; Michel, N.; Vaissiere, B.E.; Mickael, H. Differences of floral resource use between honey bees and wild bees in an intensive farming system. Agric. Ecosyst. Environ. 2013, 179, 78–86. [Google Scholar] [CrossRef]
- Piqueray, J.; Gilliaux, V.; Decruyenaere, V.; Cornelis, J.T.; Uyttenbroeck, R.; Mahy, G. Management of Grassland-like Wildflower Strips Sown on Nutrientrich Arable Soils: The Role of Grass Density and Mowing Regime. Environ. Manag. 2019, 63, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Staab, K.; Yannelli, F.A.A.; Lang, M.; Kollmann, J. Bioengineering effectiveness of seed mixtures for road verges: Functional composition as a predictor of grassland diversity and invasion resistance. Ecol. Eng. 2015, 84, 104–112. [Google Scholar] [CrossRef]
- Beyer, N.; Gabriel, D.; Westphal, C. Contrasting effects of past and present mass-flowering crop cultivation on bee pollinators shaping yield components in oilseed rape. Agric. Ecosyst. Environ. 2021, 319, 107537. [Google Scholar] [CrossRef]
- Ditner, N.; Balmer, O.; Beck, J.; Blick, T.; Nagel, P.; Luka, H. Effects of experimentally planting non-crop flowers into cabbage fields on the abundance and diversity of predators. Biodivers. Conserv. 2013, 22, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Hatt, S.; Uyttenbroeck, R.; Lopes, T.; Mouchon, P.; Chen, J.; Piqueray, J.; Monty, A.; Francis, F. Do flower mixtures with high functional diversity enhance aphid predators in wildflower strips? Eur. J. Entomol. 2017, 114, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Schütz, L.; Wenzel, B.; Rottstock, T.; Dachbrodt-Saaydeh, S.; Golla, B.; Kehlenbeck, H. How to promote multifunctionality of vegetated strips in arable farming: A qualitative approach for Germany. Ecosphere 2022, 13, e4229. [Google Scholar] [CrossRef]
- Mei, Z.; de Groot, G.A.; Kleijn, D.; Dimmers, W.; van Gils, S.; Lammertsma, D.; van Kats, R.; Scheper, J. Flower availability drives effects of wildflower strips on ground-dwelling natural enemies and crop yield. Agric. Ecosyst. Environ. 2021, 319, 107570. [Google Scholar] [CrossRef]
- Bilski, Z.; Kajdan-Zysnarska, I. Uprawa roślin bobowatych drobnonasiennych. In Centrum Doradztwa Ekologicznego W Brwinowie; Oddział w Poznaniu: Poznań, Poland, 2019. [Google Scholar]
- Wyłupek, T.; Powroźnik, M.; Widelska, M. Album roślin Miododajnych; Wojewódzki Inspektorat Ochrony Roślin i Nasiennictwa w Lublinie: Lublin, Poland, 2020; pp. 5–53.
- Haaland, C.; Gyllin, M. Butterflies and bumblebees in greenways and sown wildflower strips in southern Sweden. J. Insect Conserv. 2010, 14, 125–132. [Google Scholar] [CrossRef]
- Schmidt, A.; Kirmer, A.; Hellwig, N.; Kiehl, K.; Tischew, S. Evaluating CAP wildflower strips: High-quality seed mixtures significantly improve plant diversity and related pollen and nectar resources. J. Appl. Ecol. 2021, 59, 860–871. [Google Scholar] [CrossRef]
- An Official Website of the Institute of Meteorology and Water Management (IMGW-PIB). Available online: https://www.imgw.pl/wydarzenia/charakterystyka-wybranych-elementow-klimatu-w-polsce-w-sierpniu-2022-roku-podsumowanie (accessed on 29 November 2022).
- Bretagnolle, V.; Gaba, S. Weeds for bees? A review. Agron. Sustain. Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef] [Green Version]
- Rollin, O.; Benelli, G.; Benvenuti, S.; Decourtye, A.; Wratten, S.D.; Canale, A.; Desneux, N. Weed-insect pollinator networks as bio-indicators of ecological sustainability in agriculture. A review. Agron. Sustain. Dev. 2016, 36, 8. [Google Scholar] [CrossRef] [Green Version]
- Brandt, K.; Glemnitz, M.; Schröder, B. The impact of crop parameters and surrounding habitats on different pollinator group abundance on agricultural fields. Agric. Ecosyst. Environ. 2017, 243, 55–66. [Google Scholar] [CrossRef]
- Azpiazu, C.; Medina, P.; Adán, Á.; Sánchez-Ramos, I.; Del Estal, P.; Fereres, A.; Vinuela, E. The Role of Annual Flowering Plant Strips on a Melon Crop in Central Spain. Influence on Pollinators and Crop. Insects 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero-Castaño, A.; Ortiz-Sánchez, F.J.; Vila, M. Mass flowering crops in a patchy agricultural landscape can reduce bee abundance in adjacent shrublands. Agric. Ecosyst. Environ. 2016, 223, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Lindemann-Matthies, P.; Bose, E. Species Richness, Structural Diversity and Species Composition in Meadows Created by Visitors of a Botanical Garden in Switzerland. Landsc. Urban Plan 2007, 79, 281–307. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Family | Life Cycle |
|---|---|---|---|
| 1 | Agrostemma githago L. | Caryophyllaceae | Annual |
| 2 | Borago officinalis L. | Boraginaceae | Annual |
| 3 | Calendula officinalis L. | Asteraceae | Annual |
| 4 | Camelina sativa (L.) Crantz | Brassicaceae | Annual |
| 5 | Carum carvi L. | Apiaceae | Biennial |
| 6 | Centaurea cyanus L. | Asteraceae | Annual |
| 7 | Coriandrum sativum L. | Apiaceae | Annual |
| 8 | Fagopyrum esculentum Moench. | Polygonaceae | Annual |
| 9 | Helianthus annus L. | Asteraceae | Annual |
| 10 | Malva sp. | Malvaceae | Annual |
| 11 | Papaver rhoeas L. | Papaveraceae | Annual |
| 12 | Phacelia tanacetifolia Benth. | Boraginaceae | Annual |
| 13 | Trifolium incarnatum L. | Fabaceae | Annual |
| 14 | Trifolium resupinatum L. | Fabaceae | Annual |
| Plant Origin | Species | Color of Inflorescences | Date of Observations | |||||
|---|---|---|---|---|---|---|---|---|
| 18 May | 2 June | 14 June | 29 June | 13 July | 27 July | |||
| Mixture | Agrostemma githago | Violet | 0.00 | 0.28 | 2.34 | 0.56 | 0.79 | 3.16 |
| Borago officinalis | Blue | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.05 | |
| Calendula officinalis | Orange | 0.00 | 0.42 | 2.05 | 3.33 | 4.72 | 4.21 | |
| Camelina sativa | Yellow | 0.00 | 0.84 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Carum carvi | White | 0.19 | 0.14 | 0.00 | 0.56 | 0.79 | 1.05 | |
| Centaurea cyanus | Blue | 0.00 | 0.84 | 0.88 | 1.11 | 3.94 | 1.05 | |
| Coriandrum sativum | White | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.05 | |
| Malva sp. | Violet/pink | 0.00 | 0.14 | 3.22 | 6.11 | 0.00 | 9.48 | |
| Phacelia tanacetifolia | Violet | 26.47 | 33.10 | 6.73 | 1.11 | 1.57 | 1.05 | |
| Trifolium incarnatum | Dark pink | 70.50 | 21.09 | 1.46 | 0.00 | 0.00 | 0.00 | |
| Trifolium resupinatum | Light pink | 0.17 | 0.56 | 0.58 | 0.00 | 0.00 | 0.00 | |
| Seed bank | Anchusa arvensis L. | Blue | 0.18 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Arabidopsis thaliana (L.) Heynh. | White, yellow | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 42.11 | |
| Capsella bursa-pastoris (L.) Medik. | White | 1.05 | 2.93 | 3.22 | 0.00 | 0.00 | 0.00 | |
| Cirsium arvense (L.) Scop. | Violet | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 24.21 | |
| Conyza canadensis L. | White, yellow | 0.00 | 0.00 | 0.00 | 1.11 | 1.57 | 8.42 | |
| Geranium pusillum L. | Light pink | 0.00 | 17.46 | 1.15 | 20.00 | 36.22 | 1.05 | |
| Matricaria chamomilla L. | White, yellow | 0.54 | 18.43 | 59.63 | 6.11 | 0.00 | 0.00 | |
| Tanacetum parthenium (L.) Sch. Bip. | White, yellow | 0.00 | 0.00 | 0.00 | 20.00 | 49.61 | 0.00 | |
| Torilis japonica (Houtt.) D.C. | White | 0.00 | 0.13 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Tripleurospermum maritimum (L.) W. D. J. Koch | White, yellow | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.11 | |
| Veronica arvensis L. | Blue | 0.00 | 0.99 | 17.84 | 38.89 | 0.00 | 0.00 | |
| Viola tricolor L. | violet, White, yellow | 0.90 | 2.65 | 0.90 | 1.11 | 0.79 | 0.00 | |
| No. | Date of Observations | (SD) | Me (min./max.) | Mean Rank | Kruskal–Wallis Test (5, N = 30) |
|---|---|---|---|---|---|
| 1. | 18 May | 115.60 (±38.29) | 109.50 (58.50/155.50) | 27.2 | H = 23.469 p = 0.0003 |
| 2. | 2 June | 71.60 (±21.04) | 63.00 (53.50/105.00) | 23.0 | |
| 3. | 14 June | 34.20 (±21.25) | 21.50 (18.50/67.00) | 17.6 | |
| 4. | 29 June | 18.00 (±11.44) | 16.00 (6.00/37.00) | 10.9 | |
| 5. | 13 July | 12.70 (±7.96) | 11.00 (2.00/23.50) | 8.8 | |
| 6. | 27 July | 9.50 (±2.98) | 9.00 (5.50/13.50) | 5.5 | |
| Test statistics | ꭓ2 = 18.800 | df = 5 | p = 0.002 | ||
| Significant difference | 1–5 | 1–6 | 2–6 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, J.; Antkowiak, M.; Tymoszuk, A. Effect of Plant Seed Mixture on Overwintering and Floristic Attractiveness of the Flower Strip in Western Poland. Agriculture 2023, 13, 467. https://doi.org/10.3390/agriculture13020467
Kowalska J, Antkowiak M, Tymoszuk A. Effect of Plant Seed Mixture on Overwintering and Floristic Attractiveness of the Flower Strip in Western Poland. Agriculture. 2023; 13(2):467. https://doi.org/10.3390/agriculture13020467
Chicago/Turabian StyleKowalska, Jolanta, Małgorzata Antkowiak, and Alicja Tymoszuk. 2023. "Effect of Plant Seed Mixture on Overwintering and Floristic Attractiveness of the Flower Strip in Western Poland" Agriculture 13, no. 2: 467. https://doi.org/10.3390/agriculture13020467