
Seed Morphology and Germination of Native Tulipa Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.2. Seed Morphological Characteristics Observation
2.3. Seed Germination Test
2.3.1. Effect of Seed Morphology and Temperatures on the Germination
2.3.2. Effect of Seed Coat on the Germination
2.3.3. Effects of Gibberellic Acid (GA3) and Paclobutrazol (PAC) on the Germination
2.4. Statistical Analyses
3. Results
3.1. Seed Morphology of Tulipa Species
3.2. Seed Morphology Cluster Analysis of Tulipa Species
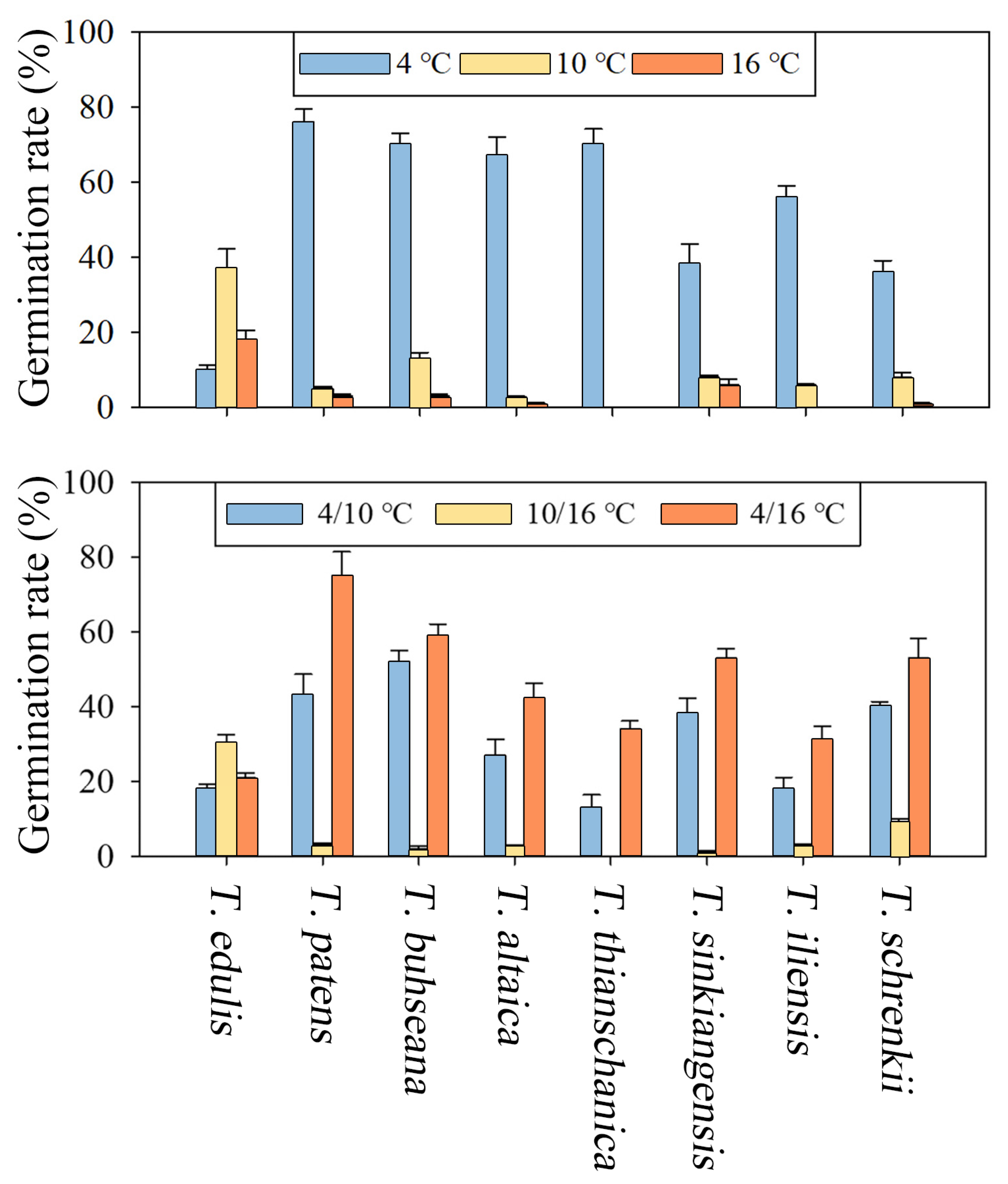
3.3. Effect of Seed Morphology and Temperatures on Germination
3.4. Effect of Seed Coat on Germination
3.5. Effect of GA3 and PAC on Germination
4. Discussion
4.1. Seed Morphology and its Taxonomic Significance for Eight Wild Tulip Species
4.2. Seed Germination Characteristics of Eight Wild Tulip Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Khaleghi, A.; Khadivi, A.; Zonneveld, B.J.M. Morphological variations among and within species of wild tulip (Tulipa L.) from Iran. Genet. Resour. Crop Evol. 2018, 65, 2241–2266. [Google Scholar] [CrossRef]
- Pourkhaloee, A.; Khosh-Khui, M.; Arens, P.; Salehi, H.; Razi, H.; Niazi, A.; Afsharifar, A.; van Tuyl, J. Molecular analysis of genetic diversity, population structure, and phylogeny of wild and cultivated tulips (Tulipa L.) by genic microsatellites. Hortic. Environ. Biotechnol. 2018, 59, 875–888. [Google Scholar] [CrossRef]
- Abedi, R.; Babaei, A.; Karimzadeh, G. Karyological and flow cytometric studies of Tulipa (Liliaceae) species from Iran. Plant Syst. Evol. 2014, 301, 1473–1484. [Google Scholar] [CrossRef]
- Xia, Y.P.; Yang, Y.A.; Yang, X.E.; Gao, X.C.; Li, F. Distribution of 14C-photosynthate and changes of endogenous hormone in the bulb development of tulip (Tulipa gesneriana L.). Acta Hortic. Sin. 2005, 32, 278–283. [Google Scholar]
- Xing, G.; Qu, L.; Zhang, W.; Zhang, Y.; Yuan, X.; Lei, J. Study on interspecific hybridization between tulip cultivars and wild species native to China. Euphytica 2020, 216, 66. [Google Scholar] [CrossRef]
- Marasek, A.; Okazaki, K. Analysis of introgression of the Tulipa fosteriana genome into Tulipa gesneriana using GISH and FISH. Euphytica 2008, 160, 217–230. [Google Scholar] [CrossRef]
- Tang, N.; Shahin, A.; Bijman, P.; Liu, J.; van Tuyl, J.; Arens, P. Genetic diversity and structure in a collection of tulip cultivars assessed by SNP markers. Sci. Hortic. 2013, 161, 286–292. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Govaerts, R.; David, J.C.; Tony, H.; Katherine, B.; Roberts, P.S.; Tuomisto, A.; Buerki, S.; Chase, M.W.; Fay, M.F. Tiptoe through the tulips-cultural history, molecular phylogenetics and classification of Tulipa (Liliaceae). Bot. J. Linn. Soc. 2013, 172, 280–328. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.Y.; Li, X.R.; Hong, D.Y. Amana kuocangshanica (Liliaceae), a new species from southeast China. Bot. J. Linn. Soc. 2007, 154, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Qu, L.; Xing, G.; Zhang, Y.; Su, J.; Zhao, Z.; Wang, W.; Lei, J. Native species of the genus Tulipa and tulip breeding in China. Acta Hortic. 2017, 1171, 357–366. [Google Scholar] [CrossRef]
- Miao, Y.; Zhu, Z.; Guo, Q.; Yang, X.; Liu, L.; Sun, Y.; Wang, C. Dynamic changes in carbohydrate metabolism and endogenous hormones during Tulipa edulis stolon development into a new bulb. J. Plant Biol. 2016, 59, 121–132. [Google Scholar] [CrossRef]
- Xing, G.; Qu, L.; Zhang, Y.; Xue, L.; Su, J.; Lei, J. Collection and evaluation of wild tulip (Tulipa spp.) resources in China. Genet. Resour. Crop Evol. 2017, 64, 641–652. [Google Scholar] [CrossRef]
- Barreto, L.C.; Echternacht, L.; Garcia, Q.S. Seed coat sculpture in Comanthera (Eriocaulaceae) and its implications on taxonomy and phylogenetics. Plant Syst. Evol. 2013, 299, 1461–1469. [Google Scholar] [CrossRef]
- Nonogaki, H.; Barrero, J.M.; Li, C.D. Editorial: Seed dormancy, germination, and pre-harvest sprouting. Front. Plant Sci. 2018, 9, 1783. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wu, F.; Xie, K.; Cheng, Z.; Guo, X.; Zhang, X.; Wang, J.; Lei, C.; Wang, J.; Mao, L.; et al. Transcriptomics Analysis Identified Candidate Genes Colocalized with Seed Dormancy QTLs in Rice (Oryza sativa L.). J. Plant Biol. 2010, 53, 330–337. [Google Scholar] [CrossRef]
- Ajay, S.N.; Animesh, B.; Rajaram, V.G. Comparative seed morphology in the genus Chlorophytum Ker Gawl. (Agavoideae, Asparagaceae) in India. Flora 2020, 273, 1–19. [Google Scholar]
- Rhie, Y.H.; Lee, S.Y. Seed dormancy and germination of Epimedium koreanum Nakai. Sci. Hortic. 2020, 272, 109600. [Google Scholar] [CrossRef]
- Rosbakha, S.; Hülsmannb, L.; Weinbergera, I.; Bleichera, M.; Poschloda, P. Bleaching and cold stratification can break dormancy and improve seed germination in Cyperaceae. Aquat. Bot. 2019, 158, 103128. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, C.C.; Baskin, J.M.; Heshmati, S.; Mirfazeli, M.S. A meta-analysis of the effects of frugivory (endozoochory) on seed germination: Role of seed size and kind of dormancy. Plant Ecol. 2018, 219, 1283–1294. [Google Scholar] [CrossRef]
- Rodriguez-Arevalo, I.; Mattana, E.; García, L.; Liu, U.; Lira, R.; Dávila, P.; Hudson, A.; Pritchard, H.W.; Ulian, T. Conserving seeds of useful wild plants in Mexico: Main issues and recommendations. Genet. Resour. Crop Evol. 2016, 64, 1141–1190. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Junior, A.G.; Mello, A.C.M.P.; Baskin, C.C.; Baskin, J.M.; Oliveira, D.M.; Garcia, Q.S. Why large seeds with physical dormancy become non-dormant earlier than small ones. PLoS ONE 2018, 13, e0202038. [Google Scholar] [CrossRef]
- Ambika, S.; Manonmani, V.; Somasundar, G. Review on Effect of Seed Size on Seedling Vigour and Seed Yield. Res. J. Seed Sci. 2014, 7, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Wyllie-Echeverria, S.; Cox, P.A.; Churchill, A.C.; Brotherson, J.D. Seed size variation within Zostera marina L. (Zosteraceae). Bot. J. Linn. Soc. 2003, 142, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Balkaya, A.; Odabas, M.S. Determination of the seed characteristics in some significant snap bean varieties grown in Samsun, Turkey. Pak. J. Biol. Sci. 2002, 5, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.; Mandal, S.M.; Chakraborty, D. Seed Size Variation: Influence on Germination and Subsequent Seedling Performance in Hyptis suaveolens (Lamiaceae). Res. J. Seed Sci. 2008, 1, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Lando, A.P.; Viana, W.G.; da Silva, R.A.; Costa, C.D.D.; Fraga, H.P.F.; Santos, M.; Mioto, P.T.; Guerra, M.P.; Steiner, N. The Physiological Relationship between Abscisic Acid and Gibberellin During Seed Germination of Trichocline catharinensis (Asteraceae) Is Associated with Polyamine and Antioxidant Enzymes. J. Plant Growth Regul. 2019, 39, 395–410. [Google Scholar] [CrossRef]
- Tang, A.-J.; Tian, M.-H.; Long, C.-L. Seed dormancy and germination of three herbaceous perennial desert ephemerals from the Junggar Basin, China. Seed Sci. Res. 2009, 19, 183–189. [Google Scholar] [CrossRef]
- Rouhi, H.R.; Karimi, F.A.; Shahbodaghlo, A.R.; Sheikhalian, M.; Rahmatabadi, R.; Samadi, M.; Karimi, F. Effects of sulfuric acid, stratification, phytohormone and potassium nitrate on dormancy breaking and germination of water lily tulip (Tulipa kaufmanniana Regel.). Int. J. Agric. Sci. 2012, 2, 136–142. [Google Scholar]
- El-Keblawy, A.; Gairola, S. Dormancy Regulating Chemicals Alleviate Innate Seed Dormancy and Promote Germination of Desert Annuals. J. Plant Growth Regul. 2016, 36, 300–311. [Google Scholar] [CrossRef]
- Chen, F.; Liu, T.; Zhou, L.L. Research on the biological and germinating characters of wild Tulipa. J. Shihezi Univ. 2001, 5, 197–200. (In Chinese) [Google Scholar]
- Zhang, W.; Qu, L.-W.; Zhao, J.; Xue, L.; Dai, H.-P.; Xing, G.-M.; Lei, J.-J. Practical Methods for Breaking Seed Dormancy in a Wild Ornamental Tulip Species Tulipa thianschanica Regel. Agronomy 2020, 10, 1765. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, X.; Liu, Z.; Cheng, Y.; Sun, T.; Yu, X. Mechanism of the Breaking of Seed Dormancy by Flower Thinning in Heracleum moellendorffii Hance. J. Plant Growth Regul. 2018, 38, 870–882. [Google Scholar] [CrossRef]
- Song, D.; Zhou, J.; Lai, L.; Alarcon, I.; Tar’An, B.; Abrams, S. Development of ABA Antagonists to Overcome ABA- and Low Temperature-Induced Inhibition of Seed Germination in Canola, Lentil, and Soybean. J. Plant Growth Regul. 2019, 39, 1403–1413. [Google Scholar] [CrossRef]
- Huang, W.; Mayton, H.S.; Amirkhani, M.; Wang, D.; Taylor, A.G. Seed dormancy, germination and fungal infestation of eastern gamagrass seed. Ind. Crop Prod. 2017, 99, 109–116. [Google Scholar] [CrossRef]
- Li, P.; Lu, R.-S.; Xu, W.-Q.; Ohi-Toma, T.; Cai, M.-Q.; Qiu, Y.-X.; Cameron, K.M.; Fu, C.-X. Comparative Genomics and Phylogenomics of East Asian Tulips (Amana, Liliaceae). Front. Plant Sci. 2017, 8, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vvedenskii, A.I.; Tulipa, L. Flora of the USSR.; Komarov, V.L., Ed.; Izd-vo Akademii Nauk USSR: Leningrad, Russia, 1936; pp. 246–280. [Google Scholar]
- Hall, A.D. The Genus Tulipa; The Royal Horticulture Society: London, UK, 1940; pp. 143–146. [Google Scholar]
- Botschantzeva, Z.P.; Varenkamp, Q. Tulips, Taxonomy, Morphology, Cytology, Phytogeography and Physiology. Taxon 1982, 31, 779. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, biogeography, and evolution of dormancy and germination. Crop Sci. 2014, 40, 564. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Rhie, Y.H.; Lee, S.Y.; Kim, K.S. Seed dormancy and germination in Jeffersonia dubia (Berberidaceae) as affected by temperature and gibberellic acid. Plant Biol. 2014, 17, 327–334. [Google Scholar] [CrossRef]
- Vandelook, F.; Bolle, N.; Van Assche, J.A. Morphological and physiological dormancy in seeds of Aegopodium podagraria (Apiaceae) broken successively during cold stratification. Seed Sci. Res. 2009, 19, 115–123. [Google Scholar] [CrossRef]
- Gao, R.; Hou, J.; Zhao, R.; Yang, X.; Hou, X.; Huo, L.; Hidayati, S.N.; Walck, J.L. Seed dormancy and germination of a critically endangered plant, Elaeagnus mollis, on the Loess Plateau of China. Eur. J. For. Res. 2021, 140, 451–461. [Google Scholar] [CrossRef]
- Gholami, A.; Sharafi, S.; Sharafi, A.; Ghasemi, S. Gemination of different seed size of pinto bean cultivars as affected by salinity and drought stress. Food Agric. Environ. 2009, 7, 555–558. [Google Scholar]
- Menaka, C.; Balamurugan, P. Seed grading techniques in Amaranthus cv. CO5. Plant Arch. 2008, 8, 729–731. [Google Scholar]
- Nonogaki, H. Seed dormancy and germination emerging mechanisms and new hypotheses. Front. Plant Sci. 2014, 5, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, J.; Zhao, L.; Fan, Y.; Shi, S.; He, L.; Hui, W. The Effects of Ascorbic Acid on Breaking the Seed Dormancy of Malus sieversii. J. Plant Growth Regul. 2018, 38, 909–918. [Google Scholar] [CrossRef]
- Subbaraj, A.K.; Funnell, K.A.; Woolley, D.J. Dormancy and Flowering Are Regulated by the Reciprocal Interaction between Cytokinin and Gibberellin in Zantedeschia. J. Plant Growth Regul. 2010, 29, 487–499. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sections | Species | Seed Sources | Latitude and Longitude (N, E) | Altitude (m) | No. of Accessions |
|---|---|---|---|---|---|
| Amana | T. edulis | Dalian city, LP | 38°57′, 121°27′ | 50 | 4 |
| Eriostemones | T. patens | Bole city, XP | 44°87′, 82°35′ | 692 | 4 |
| T. buhseana | Tuoli city, XP | 46°11′, 83°36′ | 635 | 8 | |
| Leiostemones | T. altaica | Tacheng city, XP | 47°01′, 83°12′ | 1147 | 5 |
| T. thianschanica | Zhaosu city, XP | 43°09′, 81°10′ | 1852 | 10 | |
| T. sinkiangensis | Urumchi city, XP | 43°55′, 87°49′ | 937 | 6 | |
| T. iliensis | Yamalic city, XP | 43°47′, 87°33′ | 1080 | 6 | |
| T. schrenkii | Yumin city, XP | 46°12′, 83°02′ | 715 | 5 |
| Characters | Unit or Code |
|---|---|
| Seed size (Seed length × Seed width) | mm |
| E/S (Embryo length/Seed length) | ratio |
| Seed shape | 0 = half-moon; 1 = sector |
| Seed color | 0 = deep-brown; 1 = brown; 2 = light-brown |
| Embryo morphology | 0 = linear, invisible; 1 = linear, visible |
| Testa surface | 0 = stripes; 1 = reticulate |
| Type of murus | 0 = convex; 1 = concave; 2 = flat |
| Type of meshes | 0 = smooth; 1 = granular; 2 = verrucate; 3 = powdery |
| Species | Seed Size (mm) | Embryo Length (mm) | E/S | Seed Color | Seed Shape | Embryo Morphology | Seed Coat Surface | Type of Murus | Type of Meshes |
|---|---|---|---|---|---|---|---|---|---|
| T. edulis | 5.86 × 2.43 | 1.12 ± 0.05f | 0.19 ± 0.01e | Deep- brown (175A) | Half- moon | Linear, invisible | Stripes | Concave | Smooth |
| T. patens | 6.42 × 5.17 | 2.52 ± 0.19cd | 0.40 ± 0.04bc | Brown (N170A) | Sector | Linear, visible | Reticulate | Convex | Powdery |
| T. buhseana | 5.52 × 4.30 | 3.44 ± 0.17a | 0.62 ± 0.01a | Brown (N170A) | Sector | Linear, visible | Reticulate | Concave | Smooth |
| T. altaica | 7.36 × 6.21 | 3.20 ± 0.19ab | 0.44 ± 0.03bc | Deep- brown (175B) | Sector | Linear, visible | Reticulate | Concave | Powdery |
| T. thianschanica | 4.89 × 3.86 | 2.08 ± 0.09de | 0.43 ± 0.01bc | Light- brown (164A) | Sector | Linear, visible | Reticulate | Convex | Verrucate |
| T. sinkiangensis | 6.29 × 5.13 | 2.85 ± 0.23bc | 0.45 ± 0.03b | Brown (N170B) | Sector | Linear, visible | Reticulate | Convex | Granular |
| T. iliensis | 4.53 × 3.54 | 1.65 ± 0.12e | 0.37 ± 0.02d | Light- brown (165B) | Sector | Linear, visible | Reticulate | Flat | Smooth |
| T. schrenkii | 5.43 × 3.79 | 3.18 ± 0.09ab | 0.59 ± 0.02a | Light- brown (166D) | Sector | Linear, visible | Reticulate | Flat | Smooth |
| Indexes | Seed Size | Embryo Length | E/S | Seed Color | Temperatures | Germination (T50) |
|---|---|---|---|---|---|---|
| Seed size | 1 | |||||
| Embryo length | 0.448 * | 1 | ||||
| E/S | 0.298 | 0.926 ** | 1 | |||
| Seed color | 0.133 | 0.218 | 0.102 | 1 | ||
| Temperatures | 0.035 | 0.612 ** | 0.545 ** | 0.743 ** | 1 | |
| Germination (T50) | 0.795 ** | 0.683 ** | 0.842 ** | 0.472 * | 0.944 ** | 1 |
| Species | 4 °C, ISC | 4 °C, WSC | 4/16 °C, ISC | 4/16 °C, WSC |
| T. edulis | 105.10 ± 8.17 a | 95.58 ± 6.88 b | 84.35 ± 8.50 c | 75.18 ± 6.02 d |
| T. patens | 38.83 ± 2.12 c | 36.50 ± 1.23 c | 53.70 ± 2.19 a | 44.12 ± 3.58 b |
| T. buhseana | 29.75 ± 4.13 c | 27.26 ± 3.21 c | 47.36 ± 0.83 a | 35.45 ± 3.14 b |
| T. altaica | 57.27 ± 1.95 c | 49.33 ± 5.21 d | 86.74 ± 3.32 a | 66.72 ± 5.44 b |
| T. thianschanica | 48.13 ± 3.01 b | 49.50 ± 5.08 b | 79.10 ± 3.01 a | 78.53 ± 6.31 a |
| T. sinkiangensis | 82.83 ± 8.45 a | 74.37 ± 7.27 a | 44.40 ± 1.37 b | 41.22 ± 5.24 b |
| T. iliensis | 39.50 ± 1.15 c | 37.97 ± 5.30 c | 57.97 ± 1.50 a | 50.30 ± 2.11 b |
| T. schrenkii | 65.10 ± 1.87 a | 69.53 ± 4.55 a | 52.43 ± 0.85 b | 48.30 ± 3.32 b |
| Treatments (μM) | T. edulis | T. patens | T. buhseana | T. altaica | T. thianschanica | T. sinkiangensis | T. iliensis | T. schrenkii | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 4 °C | CK | 105.10 ± 8.17 b | 38.83 ± 2.12 d | 29.75 ± 4.13 b | 57.27 ± 1.95 b | 48.13 ± 3.01 d | 82.83 ± 8.45 a | 39.50 ± 1.15 c | 65.10 ± 1.87 a | |
| GA3 | 100 | 92.38 ± 7.50 c | 42.17 ± 2.10 c | 24.30 ± 1.80 c | 52.77 ± 1.66 c | 38.67 ± 5.86 e | 58.10 ± 2.88 cd | 45.92 ± 1.25 b | 52.83 ± 1.50 d | |
| 200 | 86.35 ± 9.13 cd | 36.17 ± 2.78 e | 25.37 ± 1.21 c | 56.37 ± 0.85 b | 45.83 ± 4.01 d | 62.00 ± 4.07 c | 45.30 ± 0.79 b | 56.83 ± 1.39 c | ||
| 400 | 106.22 ± 5.92 b | 43.40 ± 0.98 c | 26.33 ± 0.81 c | 57.63 ± 1.96 b | 54.60 ± 4.93 c | 66.70 ± 4.96 b | 38.30 ± 1.05 c | 60.12 ± 1.95 b | ||
| PAC | 100 | 144.66 ± 3.59 a | 75.00 ± 2.95 b | 57.33 ± 1.53 a | 61.97 ± 1.39 a | 68.33 ± 2.52 b | 87.92 ± 5.11 a | 71.07 ± 2.76 a | 62.46 ± 0.95 a | |
| 200 | 156.37 ± 8.50 a | 77.60 ± 4.16 b | 58.67 ± 4.51 a | 67.50 ± 1.35 a | 73.66 ± 4.16 b | 85.63 ± 3.47 a | 74.37 ± 2.16 a | 64.73 ± 5.80 a | ||
| 400 | 148.22 ± 9.20 a | 81.10 ± 9.41 a | 60.60 ± 1.31 a | 64.33 ± 2.75 a | 81.33 ± 5.51 a | 84.10 ± 4.37 a | 72.77 ± 2.55 a | 65.20 ± 0.92 a | ||
| 4/16 °C | CK | 84.35 ± 8.50 b | 53.70 ± 2.19 c | 47.36 ± 0.83 b | 86.74 ± 3.32 b | 79.10 ± 3.01 c | 44.40 ± 1.37 c | 57.97 ± 1.50 c | 52.43 ± 0.85 b | |
| GA3 | 100 | 69.38 ± 5.39 d | 61.40 ± 2.15 b | 33.50 ± 2.29 c | 65.00 ± 2.42 c | 74.67 ± 8.96 cd | 44.03 ± 4.01 c | 53.07 ± 0.96 c | 50.83 ± 9.35 b | |
| 200 | 75.67 ± 9.52 c | 63.27 ± 1.90 b | 33.00 ± 1.00 c | 65.27 ± 2.15 c | 81.33 ± 3.79 c | 43.55 ± 1.03 cd | 53.10 ± 1.95 c | 45.60 ± 2.95 c | ||
| 400 | 82.85 ± 7.31 b | 64.10 ± 2.72 b | 35.23 ± 1.30 c | 68.97 ± 3.18 c | 94.33 ± 6.51 b | 46.03 ± 1.05 b | 52.43 ± 0.71 c | 50.06 ± 3.10 b | ||
| PAC | 100 | 124.40 ± 7.11 a | 113.00 ± 10.16 a | 57.83 ± 5.42 a | 104.53 ± 6.85 a | 99.67 ± 9.71 b | 47.90 ± 1.35 b | 108.53 ± 5.53 b | 68.40 ± 1.71a | |
| 200 | 132.47 ± 6.39 a | 100.67 ± 9.45 a | 57.53 ± 2.32 a | 103.33 ± 6.14 a | 140.32 ± 3.51 a | 46.43 ± 0.71 bc | 130.10 ± 1.75 a | 62.47 ± 3.91 a | ||
| 400 | 128.45 ± 6.44 a | 95.13 ± 8.95 a | 59.13 ± 1.75 a | 105.83 ± 3.93 a | 135.67 ± 7.09 a | 51.80 ± 1.37 a | 123.33 ± 2.65 a | 66.67 ± 4.66 a | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Zhao, J.; Xue, L.; Dai, H.; Lei, J. Seed Morphology and Germination of Native Tulipa Species. Agriculture 2023, 13, 466. https://doi.org/10.3390/agriculture13020466
Zhang W, Zhao J, Xue L, Dai H, Lei J. Seed Morphology and Germination of Native Tulipa Species. Agriculture. 2023; 13(2):466. https://doi.org/10.3390/agriculture13020466
Chicago/Turabian StyleZhang, Wei, Jun Zhao, Li Xue, Hanping Dai, and Jiajun Lei. 2023. "Seed Morphology and Germination of Native Tulipa Species" Agriculture 13, no. 2: 466. https://doi.org/10.3390/agriculture13020466