
Genetic Variations among Fleabane (Conyza bonariensis (L.) Cronquist) Populations in Jordan and Their Susceptibility Levels to Contact Herbicides
 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Conyza Bonariensis Populations
2.2. DNA Analysis of Conyza spp. Populations
2.2.1. Extraction Quantification and Qualification of Total Genomic DNA
2.2.2. PCR Conditions
2.2.3. Preparation of Agarose Gel
2.2.4. Sequencing Protocol
2.3. Field Trial at Tal-al-Ramel
2.4. Herbicide Treatment Pots Experiments
2.4.1. Experiment 1
2.4.2. Experiment 2
2.5. Statistic Analysis
3. Results
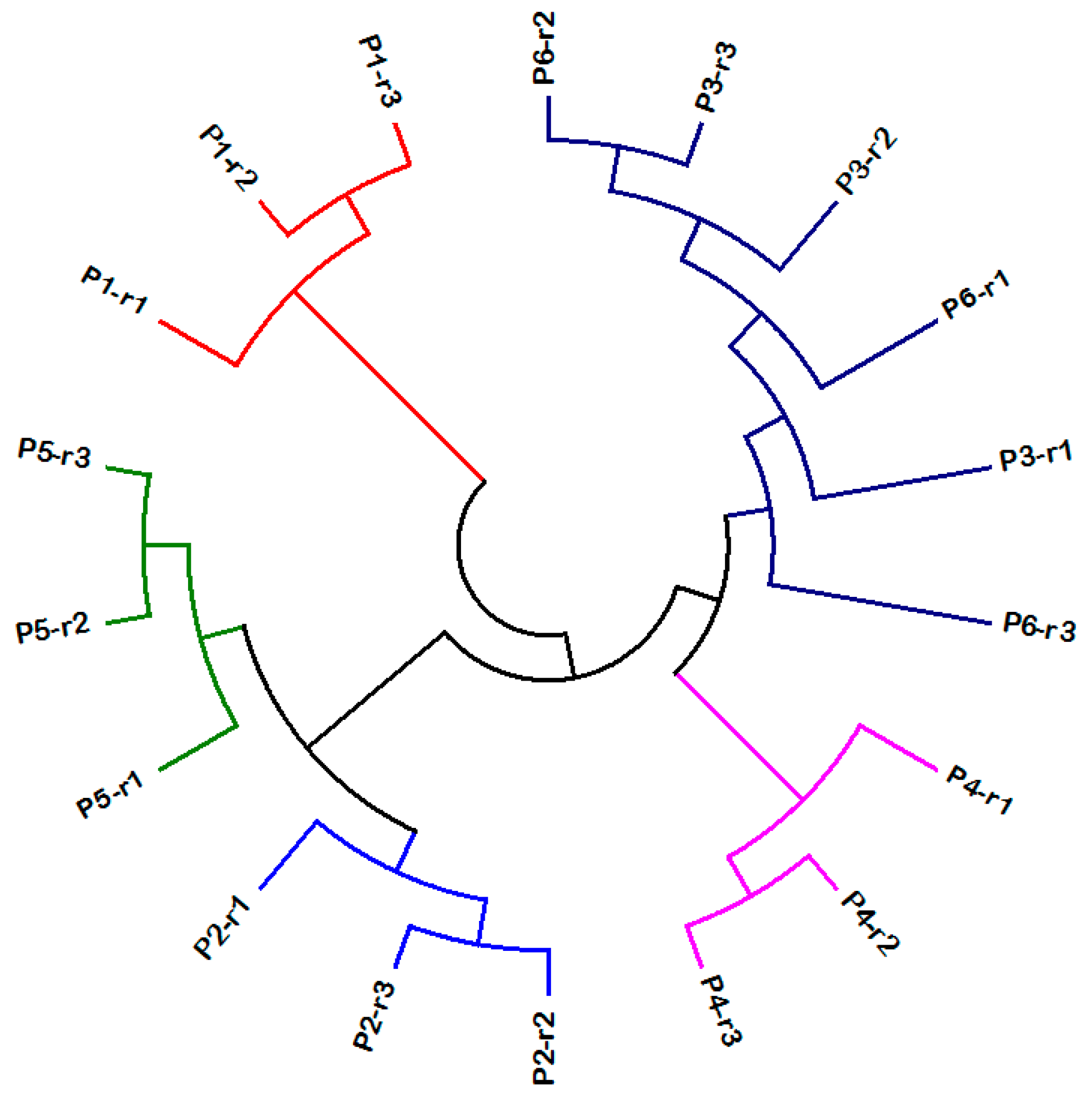
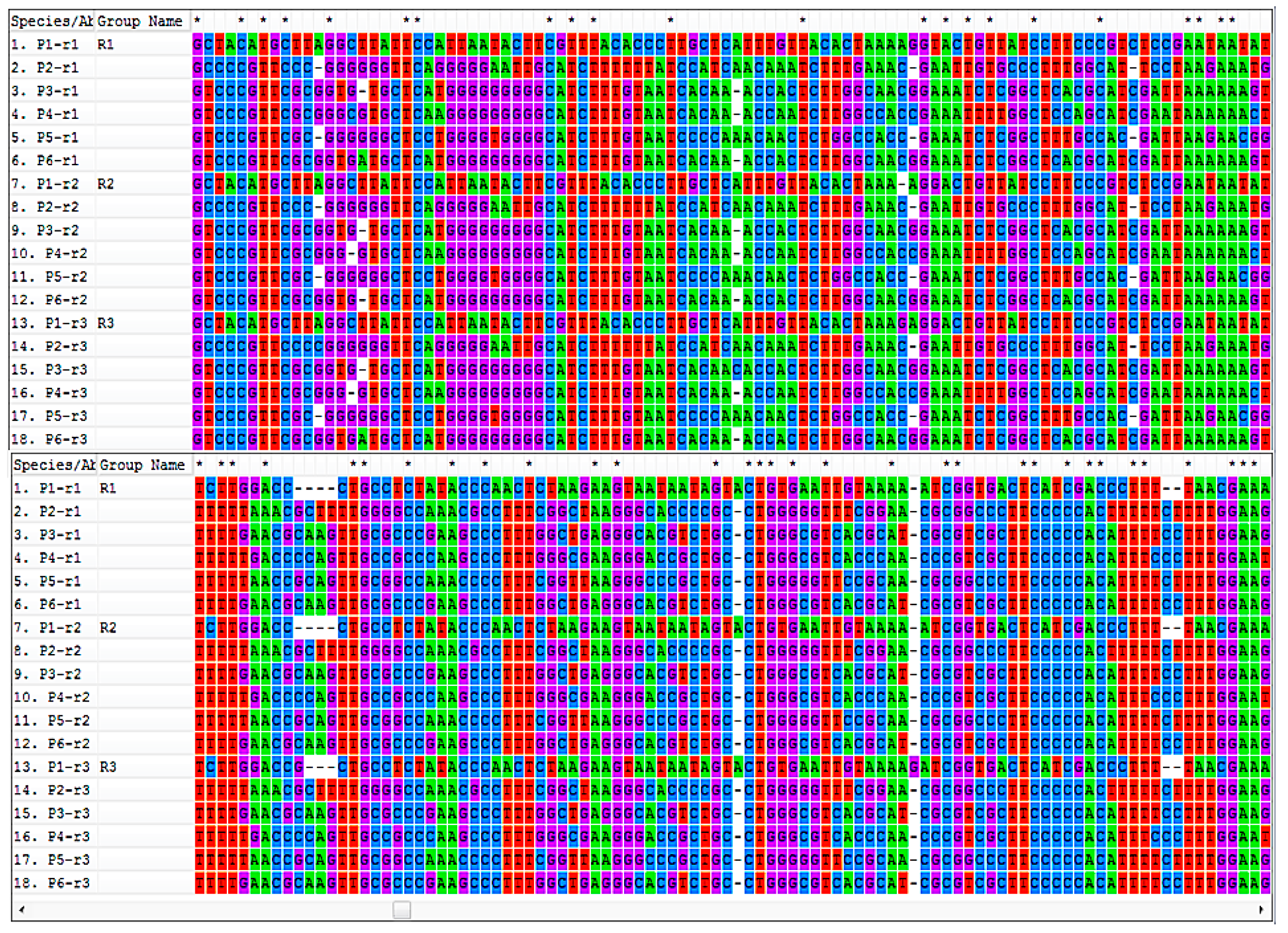
3.1. Molecular Analysis of C. Bonariensis Seed Samples
3.2. Field Trial at Tal-al-Ramel
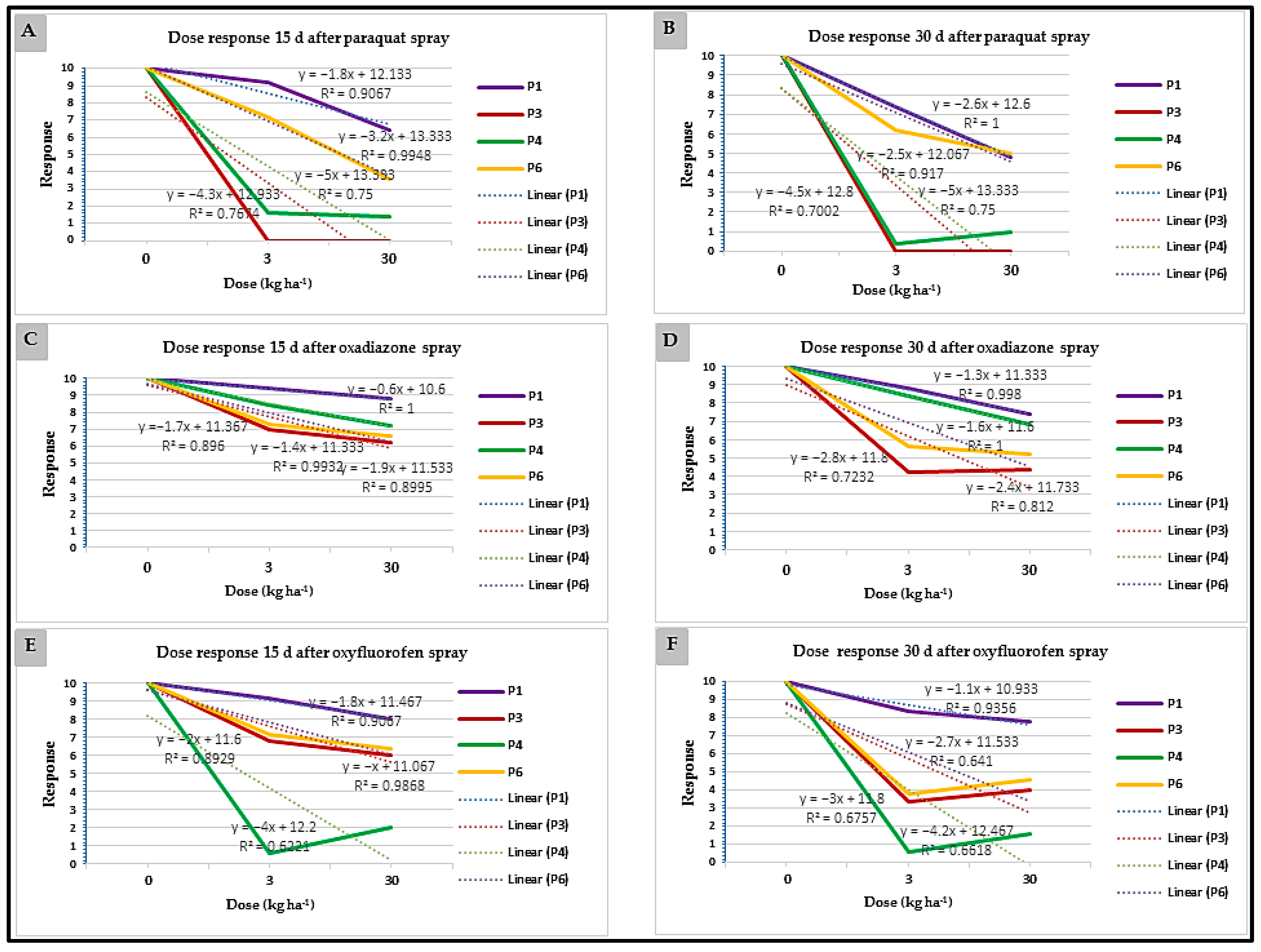
3.3. Glasshouse Experiments
3.3.1. Experiment 1
3.3.2. Experiment 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heap, I. The International Survey of Herbicide-Resistant Weeds. 2017. Available online: https://weedscience.org/Summary/Species.aspx (accessed on 8 May 2020).
- Smith, C.M.; Namuth, D. Herbicide Resistance: Mechanisms, Inheritance, and Molecular Genetics. eLearn and Grow Library, 2005, PP, 7. Available online: http://plantand soil.unl.edu/croptechnology2005/pageincludes/printModule.jsp?inform (accessed on 22 August 2021).
- Gunsolus, J.L. Herbicide Resistant Weeds. Regents of the University of Minnesota. University of Minnesota Extension. 2008. Available online: http://www.extension.umn.edu/distribution/cropsystems/DC6077.html (accessed on 14 March 2020).
- Qasem, J.R. Herbicide-Resistant Weeds: The Technology and Weed Management. In Herbicides—Current Research and Case Studies in Use; Price, A.J., Kelton, J.A., Eds.; InTech: Rang-Du-Fliers, France, 2013; pp. 445–471. Available online: https://www.intechopen.com/articles/show/title/herbicide-resistant-weeds-the-technology-and-weed-management (accessed on 3 May 2020).
- Holm, L.G.; Doll, J.; Holm, E.; Pancho, J.V.; Herberger, J.P. World Weeds, Natural Histories and Distribution; Wiley: Hoboken, NJ, USA, 1997. [Google Scholar]
- Heap, I. The International Survey of Herbicide Resistant Weeds. 2022. Available online: www.weedscience.org (accessed on 7 November 2022).
- Monsanto Company. Management Guide for Marestail. For Cropland and Orchards in CA and AZ. 2005. Available online: http://www.monsanto.com/weedmanagement/Documents/Marestail.pdf (accessed on 16 June 2016).
- Urbano, J.M.; Borrego, A.; Torres, V.; Leon, J.M.; Jimenez, C.; Dinelli, G.; Barnes, J. Glyphosate-resistant hairy fleabane (Conyza bonariensis) in Spain. Weed Technol. 2007, 21, 396–401. [Google Scholar] [CrossRef]
- Dinelli, G.; Marotti, I.; Bonetti, A.; Catizone, P.; Urbano, J.M.; Barnes, J. Physiological and molecular bases of glyphosate resistance in Conyza bonariensis biotypes from Spain. Weed Res. 2008, 48, 257–265. [Google Scholar] [CrossRef]
- Ferreira, E.A.; Galon, L.; Aspiazú, I.; Silva, A.A.; Concenço, G.; Silva, A.F.; Oliveira, J.A.; Vargas, L. Glyphosate translocation in hairy fleabane (Conyza bonariensis) biotypes. Planta Daninha 2008, 26, 637–643. [Google Scholar] [CrossRef]
- Travlos, I.S.; Chachalis, D. Glyphosate-resistant hairy fleabane (Conyza bonariensis) is reported in Greece. Weed Technol. 2010, 24, 569–573. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide-Resistant Weeds. Weed Science. 2012. Available online: www.weedscience.com (accessed on 8 May 2012).
- Sansom, M.; Saborido, A.A.; Dubois, M. Control of Conyza spp. with glyphosate—A review of the situation in Europe. Plant Prot. Sci. 2013, 49, 44–53. [Google Scholar] [CrossRef]
- Mylonas, F.N.; Giannopolitis, C.N.; Efthimiadis, P.G.; Menexes, G.C.; Madesis, P.B.; Eleftherohorinos, I.G. Glyphosate resistance of molecularly identified Conyza albida and Conyza bonariensis populations. Crop Prot. 2014, 65, 207–215. [Google Scholar] [CrossRef]
- Okada, M.; Hanson, B.D.; Hembree, K.J.; Peng, Y.; Shrestha, A.; Stewart, C.N.; Wright, S.D.; Jasieniuk, M. Evolution and spread of glyphosate resistance in Conyza bonariensis in California and a comparison with closely related Conyza canadensis. Weed Res. 2015, 55, 173–184. [Google Scholar] [CrossRef]
- Kaspary, T.E.; Lamego, F.; Langaro, A.; Ruchel, Q.; Agostinetto, D. Investigation of the mechanism of resistance to glyphosate herbicide in hairy fleabane. Planta Daninha 2016, 34, 555–564. [Google Scholar] [CrossRef]
- Aves, C.S. Herbicide Resistance in Conyza bonariensis (L) Cronquist (Flaxleaf Fleabane) Populations from Northeast Victoria and Its Management in Mixed Farming Systems. Ph.D. Thesis, School of Agriculture, Food and Wine, University of Adelaide, Adelaide, Australia, 2017; p. 100. [Google Scholar]
- Hereward, J.P.; Werth, J.A.; Thornby, D.F.; Keenan, M.; Chauhan, B.S.; Walter, G.H. Complete chloroplast genome of glyphosate resistant Conyza bonariensis (L.) Cronquist from Australia. Mitochondrial DNA Part B Resour. 2017, 2, 444–445. [Google Scholar] [CrossRef]
- Santo, T.L.d.E.; de Mendonga, C.G.; Graichen, F.A.S.; Raetano, C.G.; Teodoro, P.E. Control of Conyza bonariensis with glyphosate associated to adjuvants applied with different spray nozzles. Biosci. J. Uberlandia 2017, 33, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Szigeti, Z.; Rácz, I.; Lásztity, D. Paraquat resistance of weeds—The case of Conyza canadensis (L.) Cronq. Z. Naturforsch. 2001, 56, 319–328. [Google Scholar] [CrossRef]
- Moretti, M.L.; Hanson, B.D. Reduced translocation is involved in resistance to glyphosate and paraquat in Conyza bonariensis and Conyza canadensis from California. Weed Res. 2017, 57, 25–34. [Google Scholar] [CrossRef]
- Vargas, L.; Bianchi, M.A.; Rizzardi, M.A.; Agostinetto, D.; e Dal Magro, T. Conyza bonariensis biotypes resistant to the glyphosate in Southern Brazil. Planta 2007, 25, 573–578. [Google Scholar]
- Shaaltiel, Y.; Gressel, J. Multienzyme oxygen radical detoxifying system correlated with paraquat resistance in Conyza bonariensis. Pestic. Biochem. Physiol. 1986, 26, 22–28. [Google Scholar] [CrossRef]
- Fuerst, E.P.; Nakatani, H.Y.; Dodge, A.D.; Penner, D.; Arntzen, C.J. Paraquat resistance in Conyza. Plant Physiol. 1985, 77, 984–989. [Google Scholar] [CrossRef] [PubMed]
- Thébaud, C.; Abbott, R. Characterization of invasive Conyza species (Asteracae) in Europe: Quantitative trait and isozyme analysis. Am. J. Bot. 1995, 82, 360–368. [Google Scholar] [CrossRef]
- Van Gessel, M.J. Glyphosate-resistant horseweed from Delaware. Weed Sci. 2001, 49, 703–705. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Hernández, F.; Alves, M.F.; Machado, R.M.; Sun, Y.-Y.; Wang, M.-R.; Zhang, C.-B.; Hao, J.-H. Genetic diversity and population structure of invasive and native populations of Erigeron canadensis L. J. Plant Ecol. 2022, 15, 864–876. [Google Scholar] [CrossRef]
- Soares, A.A.F.; Fregonezi, A.M.D.; Mangolin, C.A.; Collet, S.A.; Machado, R.S. Evidence of high gene flow between samples of horseweed (Conyza canadensis) and hairy fleabane (Conyza bonariensis) as revealed by isozyme polymorphisms. Weed Sci. 2015, 63, 604–612. [Google Scholar] [CrossRef]
- Shaik, R.S.; Zhu, X.; Clements, D.R.; Weston, L.A. Understanding invasion history and predicting invasive niches using genetic sequencing technology in Australia: Case studies from Cucurbitaceae and Boraginaceae. Conserv. Physiol. 2016, 4, cow030. [Google Scholar] [CrossRef]
- Alfalahi, A.O.; Saleh, A.T. Molecular detection of Johnsongrass Sorghum halepense L. evalution in Iraq. Asian J. Plant Sci. 2018, 17, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; p. 2100. [Google Scholar]
- Vogelstein, B.; Gillespie, D. Preparative and analytical purification of DNA from agarose. Proc. Natl. Acad. Sci. USA. 1979, 76, 615–619. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. SAS/STAT® 9.1 User’s Guide. Ver. 9.1; SAS Institute Inc.: Cary, NC, USA, 2004. [Google Scholar]
- Rogers, J.S. Maximum likelihood estimation of phylogenetic trees is consistent when substitution rates vary according to the invariable sites plus gamma distribution. Syst. Biol. 2001, 50, 713–722. [Google Scholar] [CrossRef]
- Walia, S.; Banerjee, T.; Kumar, R. Efficacy of weed management techniques onw control, biomass yield, and soil herbicide residue in transplanted wild marigold (Tagetes minuta L.) under high rainfall conditions of western Himalaya. Agronomy 2021, 11, 2119. [Google Scholar] [CrossRef]
- Scarabel, L.; Farinati, S.; Sattin, M. Occurrence of resistance to ALS inhibitors in European Cyperus Esculentus L.: Characterization and implications for management. Agronomy 2020, 10, 1133. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site of Collection | Conyza spp. | Symbol | Field Description | Geographical Coordinates |
|---|---|---|---|---|
| Tal-al-Ramel | Conyza bonariensis | P1 | Date palm orchard | 32.059821° N, 35.598875° E |
| University of Jordan Research Station | Conyza canadensis | P2 | Cultivated fields | 32.145968° N, 35.696554° E |
| Conyza bonariensis | P3 | Cultivated fields | ||
| Al-Twal | Conyza bonariensis | P4 | Uncultivated fields | 32.147574° N, 35.692870° E |
| Conyza canadensis | P5 | Uncultivated fields | ||
| The University of Jordan Campus | Conyza bonariensis | P6 | Gardens | 32.0161° N, 35.8695° E |
| Common Name | Trade Name and a.i Percentage | Chemical Name | Rate of Application (kg ha−1) | Mode of Action |
|---|---|---|---|---|
| Paraquat | Gramaxon 20% (v/v) | 1,1′-dimethyl-(4,4′-bipyridiniom) dichloride | 2.5 | Contact |
| Oxadiazon | Ronstar 25%(v/v) | 2-tert-butyl-4-(2,4-dichloro-5-isopropyloxyphenyl)-1,3,4-oxadiazolin-5-one | 5 | Contact |
| Oxyfluorfen | Goal 24%(v/v) | [2-chloro-N-[[4-methoxy-6-methyl-I,3,5-triazine-2-yl)-amino]carbboennyzl]esulphonamide | 3.3 | Contact |
| Treatments | 15 Days after Herbicides Application (DAT) |
|---|---|
| Untreated (control) | 10.0 a |
| Paraquat (2.5 kg ha−1) | 10.0 ± 0.4 a |
| Paraquat (5 kg ha−1) | 10.0 ± 0.4 a |
| Paraquat (25 kg ha−1) | 9.0 ± 0.6 ab |
| Oxadiazon (5.0 kg ha−1) | 7.8 ± 0.5 b |
| Oxadiazon (10.0 kg ha−1) | 7.4 ± 0.6 b |
| Oxadiazon (50 kg ha−1) | 7.8 ± 0.6 b |
| Oxyfluorfen (3.3 kg ha−1) | 9.8 ± 0.6 a |
| Oxyfluorfen (6.6 kg ha−1) | 10.0 ± 0.5 a |
| Oxyfluorfen (33 kg ha−1) | 7.2 ± 0.7 b |
| LSD ≤ p (0.05) | 1.8 |
| Treatments | Recommended Rates of Application (kg ha−1) | First Spray 15 October 2019 Score Out of 10 | Second Spray 30 October 2019 Score Out of 10 |
|---|---|---|---|
| Date palm population (Tal-al-Ramel) of C. bonariensis (P 1) | |||
| Control | 0.0 | 10.0 ± 0.6 a | 10.0 ± 0.2 a |
| Paraquat | 2.5 | 9.2 ± 0.3 ab | 7.4 ± 0.5 dc |
| Oxadiazon | 5.0 | 9.4 ± 0.5 ab | 8.8 ± 0.2 ab |
| Oxyfluorfen | 3.0 | 9.2 ± 0.3 ab | 8.4 ± 0.3 bc |
| University of Jordan Research Station Population of C. bonariensis (P3) | |||
| Control | 0.0 | 10.0 ± 0.3 a | 10.0 ± 0.3 a |
| Paraquat | 2.5 | 0.0 ± 1.4 e | 0.0 ± 1.2 g |
| Oxadiazon | 5.0 | 7.0 ± 1.2 c | 4.2 ± 1.0 f |
| Oxyfluorfen | 3.0 | 6.8 ± 0.9 c | 3.4 ± 1.3 f |
| Al-Twal Population of C. bonariensis (P4) | |||
| Control | 0.0 | 10.0 ± 0.8 a | 10.0 ± 0.3 a |
| Paraquat | 2.5 | 1.6 ± 1.3 d | 0.4 ± 1.4 g |
| Oxadiazon | 5.0 | 8.4 ± 0.4 b | 8.4 ± 0.3 bc |
| Oxyfluorfen | 3.0 | 0.6 ± 0.6 de | 0.6 ± 1.2 g |
| The University of Jordan Campus Population of C. bonariensis (P6) | |||
| Control | 0.0 | 10.0 ± 0.5 a | 10.0 ± 0.4 a |
| Paraquat | 2.5 | 7.2 ± 0.3 c | 6.2 ± 0.6 de |
| Oxadiazon | 5.0 | 7.3 ± 0.6 c | 5.6 ± 0.6 e |
| Oxyfluorfen | 3.0 | 7.2 ± 0.3 c | 3.8 ± 0.8 f |
| LSD (p = 0.05) | - | 1.2 | 1.2 |
| Treatments | High Rates of Application (kg ha−1) | First Spray 15 October 2019 Score Out of 10 | Second Spray 30 October 2019 Score Out of 10 |
|---|---|---|---|
| Date palm population of C. bonariensis (P1) | |||
| Control | 0 | 10.0 ±0.4 a | 10.0 ±0.2 a |
| Paraquat | 25 | 6.4 ± 0.8 cd | 4.8 ± 1.3 c |
| Oxadiazon | 50 | 8.8 ± 0.6 ab | 7.4 ± 0.5 b |
| Oxyfluorfen | 30 | 8.0 ± 0.7 bc | 7.8 ± 0.6 b |
| University of Jordan Research Station Population of C. bonariensis (P3) | |||
| Control | 0 | 10.0 ± 0.3 a | 10.0 ± 0.3 a |
| Paraquat | 25 | 0.0 ± 1.3 g | 0.0 ± 1.2 e |
| Oxadiazon | 50 | 6.2 ± 0.8 cd | 4.4 ± 0.9 c |
| Oxyfluorfen | 30 | 6.0 ± 0.7 d | 4.0 ± 0.8 c |
| Al-Twal Population of C. bonariensis (P4) | |||
| Control | 0 | 10.0 ± 0.3 a | 10.0 ± 0.2 a |
| Paraquat | 25 | 1.4 ± 0.5 fg | 1.0 ± 1.3 de |
| Oxadiazon | 50 | 7.2 ± 0.6 bcd | 6.8 ± 0.7 b |
| Oxyfluorfen | 30 | 2.0 ± 0.3 ef | 1.6 ± 1.3 d |
| The University of Jordan Campus Population of C. bonariensis (P6) | |||
| Control | 0 | 10.0 ± 0.3 a | 10.0 ± 0.1 a |
| Paraquat | 25 | 3.6 ± 1.2 e | 5.0 ± 1.0 c |
| Oxadiazon | 50 | 6.6 ± 0.8 cd | 5.2 ± 0.6 c |
| Oxyfluorfen | 30 | 6.4 ± 0.7 cd | 4.6 ± 0.7 c |
| LSD (p = 0.05) | - | 1.9 | 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qasem, J.R.; Alfalahi, A.O.; Alsubeie, M.S.; Almehemdi, A.F.; Synowiec, A. Genetic Variations among Fleabane (Conyza bonariensis (L.) Cronquist) Populations in Jordan and Their Susceptibility Levels to Contact Herbicides. Agriculture 2023, 13, 435. https://doi.org/10.3390/agriculture13020435
Qasem JR, Alfalahi AO, Alsubeie MS, Almehemdi AF, Synowiec A. Genetic Variations among Fleabane (Conyza bonariensis (L.) Cronquist) Populations in Jordan and Their Susceptibility Levels to Contact Herbicides. Agriculture. 2023; 13(2):435. https://doi.org/10.3390/agriculture13020435
Chicago/Turabian StyleQasem, Jamal Ragheb, Ayoob Obaid Alfalahi, Moodi Saham Alsubeie, Ali Fadaam Almehemdi, and Agnieszka Synowiec. 2023. "Genetic Variations among Fleabane (Conyza bonariensis (L.) Cronquist) Populations in Jordan and Their Susceptibility Levels to Contact Herbicides" Agriculture 13, no. 2: 435. https://doi.org/10.3390/agriculture13020435