
Stimulation of Early Post-Emergence Growth of Alopecurus myosuroides and Apera spica-venti Following Spray Application of ACCase Inhibitors

Abstract
:1. Introduction
2. Materials and Methods
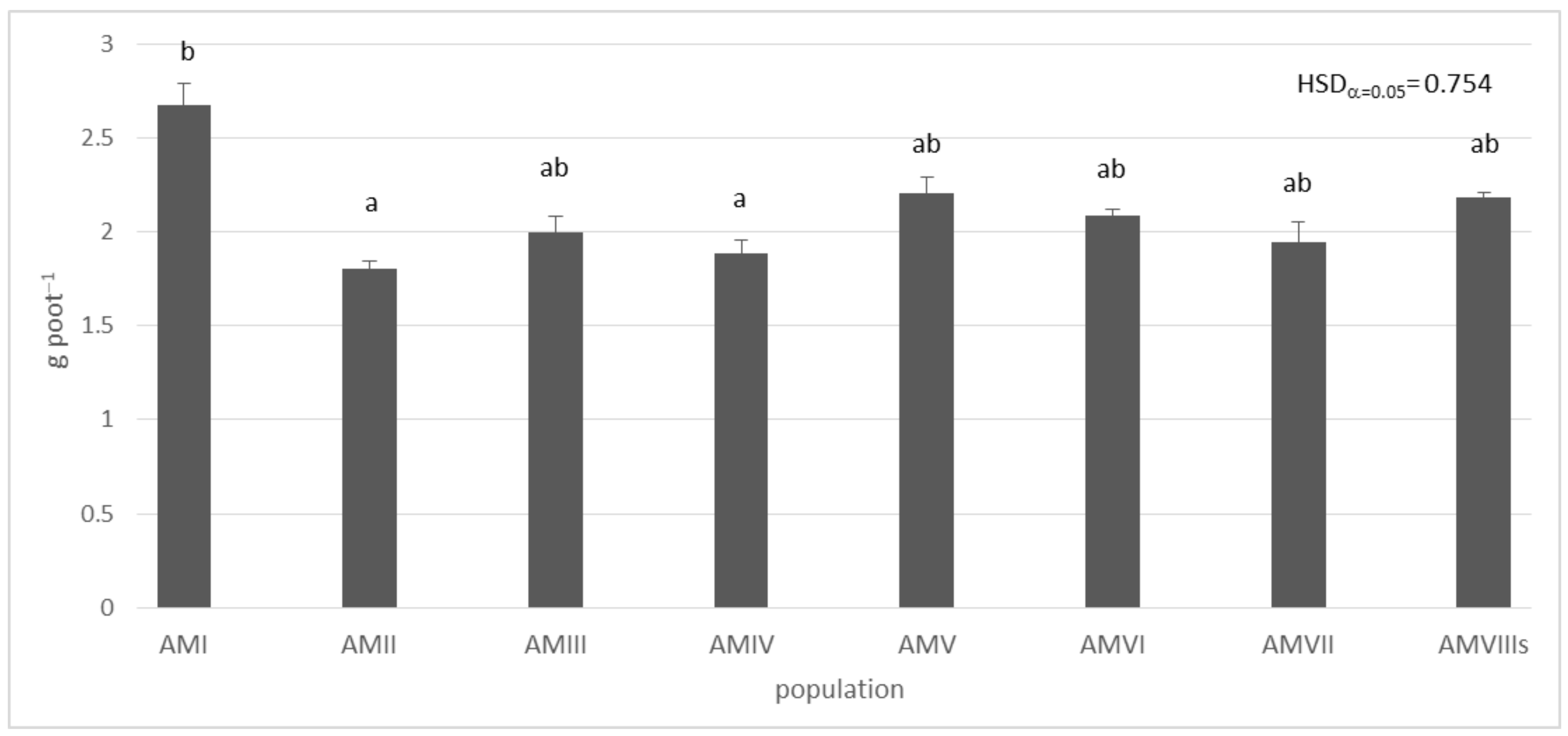
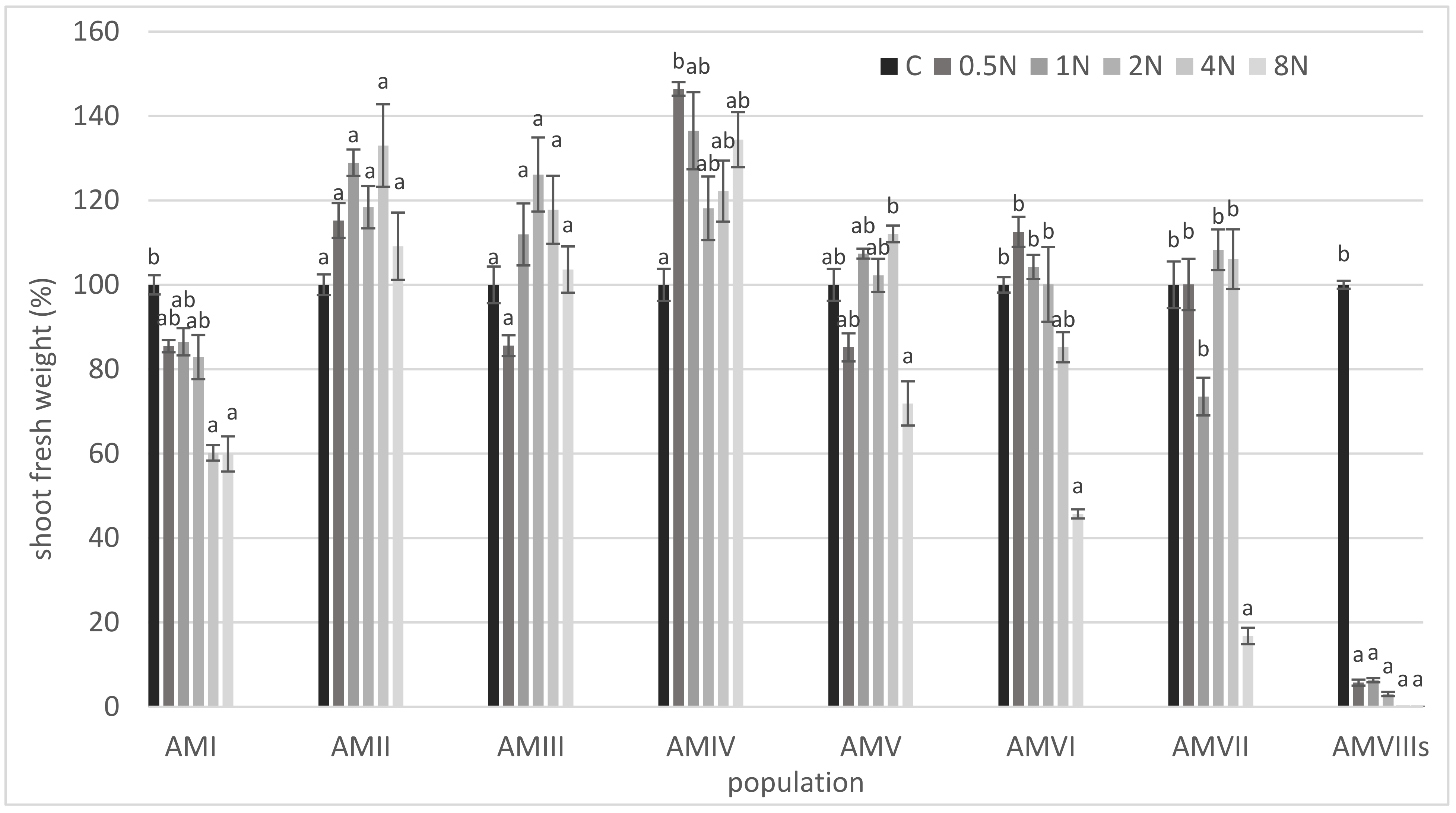
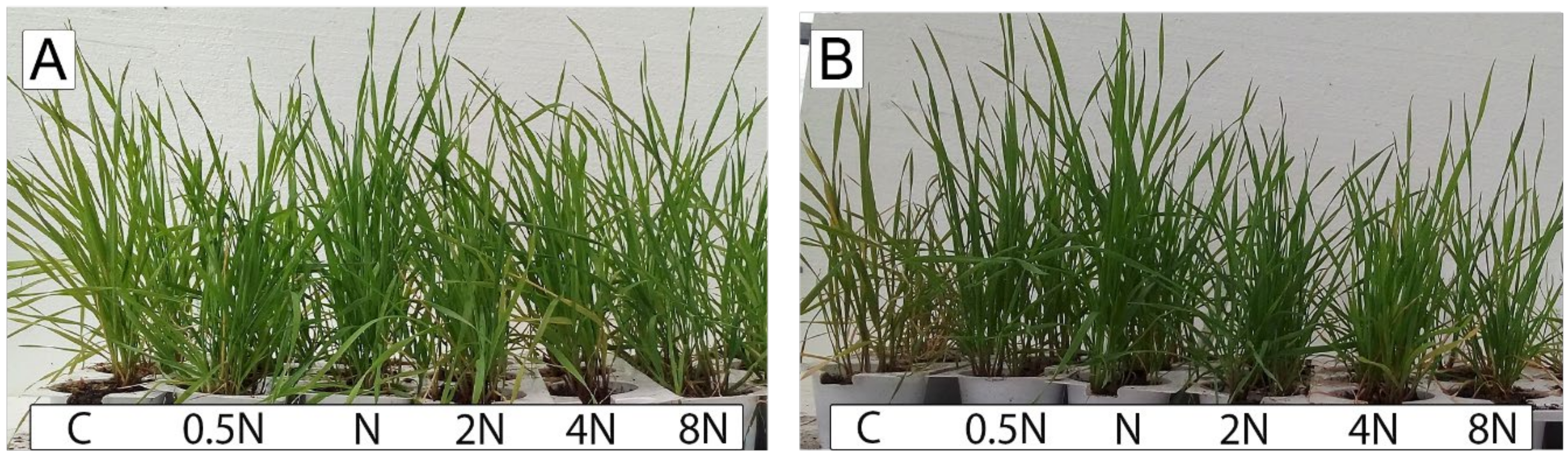
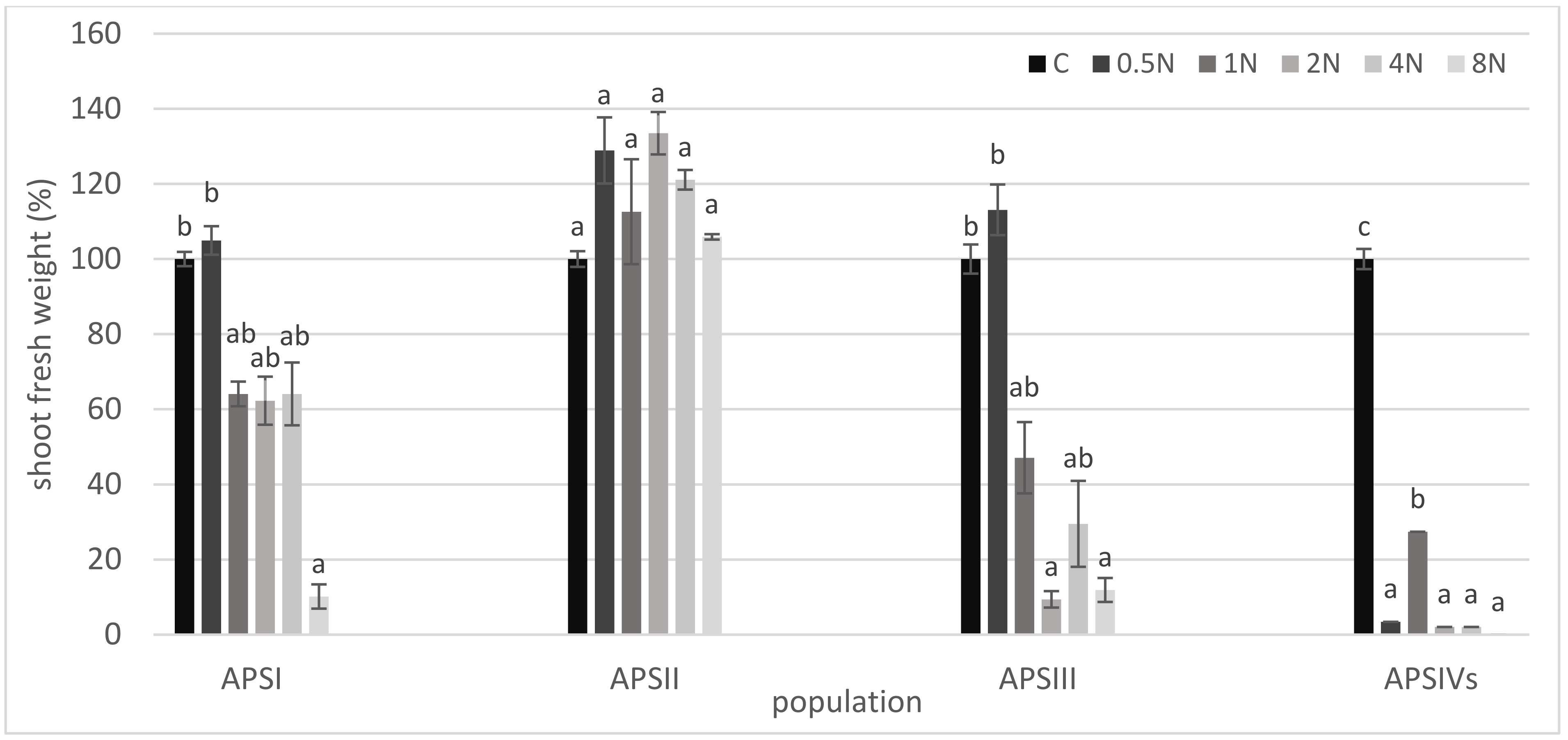
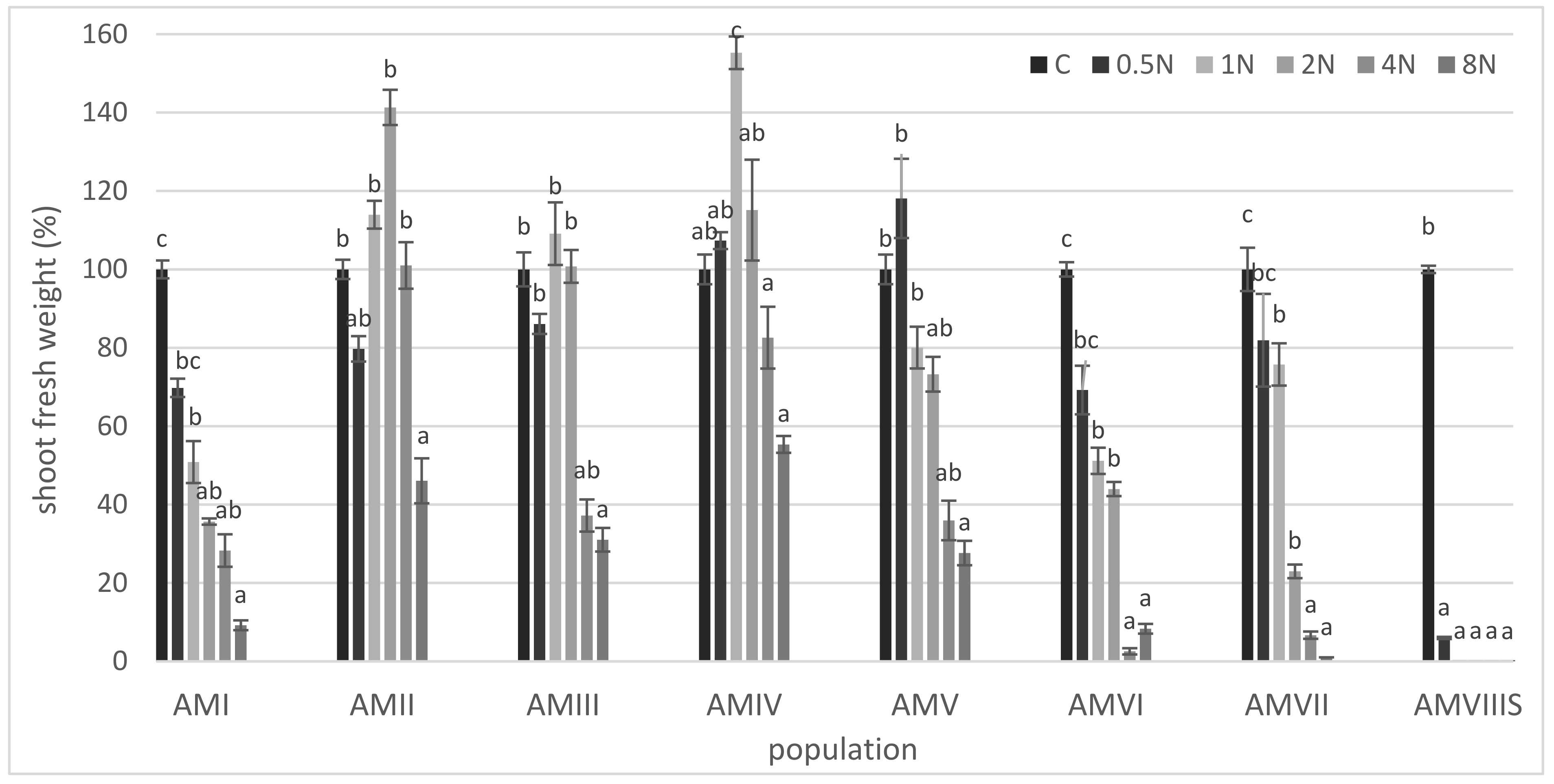
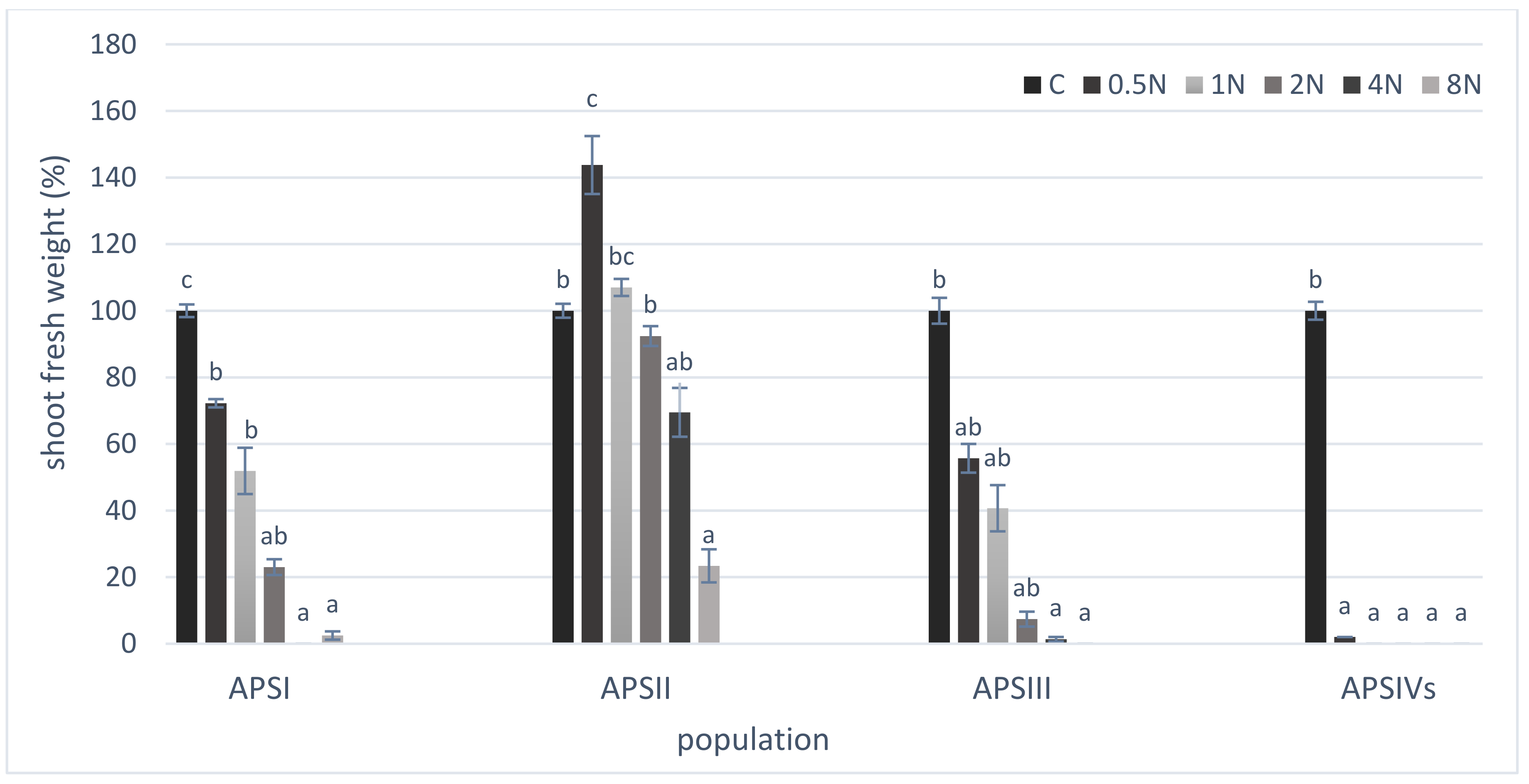
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraehmer, H.; Laber, B.; Rosinger, C.; Schulz, A. Herbicides as weed control agents: State of the art: I. weed control research and safener technology: The path to modern agriculture. Plant Physiol. 2014, 166, 1119–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stankiewicz-Kosyl, M.; Synowiec, A.; Haliniarz, M.; Wenda-Piesik, A.; Domaradzki, K.; Parylak, D.; Wrochna, M.; Pytlarz, E.; Gala-Czekaj, D.; Marczewska-Kolasa, K.; et al. Herbicide resistance and management options of Papaver rhoeas L. and Centaurea cyanus L. in Europe: A Review. Agronomy 2020, 10, 874. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Ann. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitarafan, Z.; Andreasen, C. Seed production and retention at maturity of blackgrass (Alopecurus myosuroides) and silky windgrass (Apera spica-venti) at wheat harvest. Weed Sci. 2020, 68, 151–156. [Google Scholar] [CrossRef]
- Hull, R.; Tatnell, L.V.; Cook, S.K.; Beffa, R.; Moss, S.R. Current status of herbicide-resistant weeds in the UK. Asp. App. Biol. 2014, 127, 261–272. [Google Scholar]
- Massa, D.; Gerhards, R. Investigation on herbicide resistance in European silky bentgrass (Apera spica-venti) populations. J. Plant Dis. Prot. 2011, 118, 31–39. [Google Scholar] [CrossRef]
- Melander, B. Population Dynamics of Apera spica-venti as Influenced by Cultural Methods. In Proceedings of the Brighton Crop Protection Conference—Weeds, Brighton, UK, 22–25 November 1993; pp. 107–112. [Google Scholar]
- Babineau, M.; Mahmood, K.; Mathiassen, S.K.; Kudsk, P.; Kristensen, M. De novo transcriptome assembly analysis of weed Apera spica-venti from seven tissues and growth stages. BMC Genom. 2017, 18, 128. [Google Scholar] [CrossRef] [Green Version]
- Hamouzová, K.; Soukup, J.; Jursik, M.; Hamouz, P.; Venclová, V.; Tumová, P. Cross-resistance to three frequently used sulfonylurea herbicides in populations of Apera spica-venti from the Czech Republic. Weed Res. 2011, 51, 113–122. [Google Scholar] [CrossRef]
- Adamczewski, K.; Kaczmarek, S.; Kierzek, R.; Matysiak, K. Significant increase of weed resistance to herbicides in Poland. J. Plant Prot. Res. 2019, 59, 139–150. [Google Scholar]
- Petersen, J.; Raffen, H. Evolution of herbicide resistance in Alopecurus myosuroides and Apera spica-venti in German cereal during the last 15 years. Jul.-Kühn-Arch. 2020, 464, 326–332. [Google Scholar]
- Massa, D.; Kaiser, Y.I.; Andújar-Sánchez, D.; Carmona-Alférez, R.; Mehrtens, J.; Gerhards, R. Development of a Geo-Referenced Database for Weed Mapping and Analysis of Agronomic Factors Affecting Herbicide Resistance in Apera spica-venti L. Beauv. (Silky Windgrass). Agronomy 2013, 3, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Stankiewicz-Kosyl, M.; Wrochna, M.; Tołłoczko, M. Increase in resistance to sulfonylurea herbicides in Alopecurus myosuroides populations in north-eastern Poland. Zemdirbyste 2020, 107, 249–254. [Google Scholar] [CrossRef]
- Maréchal, P.Y.; Henriet, F.; Vancutsem, F.; Bodson, B. Ecological review of black-grass (Alopecurus myosuroides Huds.) propagation abilities in relationship with herbicide resistance. Biotechnol. Agron. Soc. Environ. 2012, 16, 103–113. [Google Scholar]
- Synowiec, A.; Jop, B.; Domaradzki, K.; Podsiadło, C.; Gawęda, D.; Wacławowicz, R.; Wenda-Piesik, A.; Nowakowski, M.M.; Bocianowski, J.; Marcinkowska, K.; et al. Environmental Factors Effects on Winter Wheat Competition with Herbicide-Resistant or Susceptible Silky Bentgrass (Apera spica-venti L.) in Poland. Agronomy 2021, 11, 871. [Google Scholar] [CrossRef]
- Wenda-Piesik, A.; Synowiec, A.; Marcinkowska, K.; Wrzesińska, B.; Podsiadło, C.; Domaradzki, K.; Kuc, P.; Kwiecińska-Poppe, E. Intra- and interspecies competition of blackgrass and wheat in the context of herbicidal resistance and environmental conditions in Poland. Sci. Rep. 2022, 12, 8720. [Google Scholar] [CrossRef]
- Delye, C.; Michel, S.; Berard, A.; Chauvel, B.; Brunel, D.; Guillemin, J.-P.; Dessaint, F.; Le Corre, V. Geographical variation in resistance to acetyl-coenzyme A carboxylase inhibiting herbicides across the range of the arable weed Alopecurus myosuroides (black-grass). New Phytol. 2010, 186, 1005–1017. [Google Scholar] [CrossRef]
- Rosenhauer, M.; Jaser, B.; Felsenstein, F.; Petersen, J. Development of target-site resistance (TSR) in Alopecurus myosuroides in Germany between 2004–2012. J. Plant Dis. Prot. 2013, 120, 179–187. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Neve, P.; Powles, S.B. Fitness costs associated with evolved herbicide resistance alleles in plants. New Phytol. 2009, 184, 751–767. [Google Scholar] [CrossRef] [Green Version]
- Babineau, M.; Mathiassen, S.K.; Kristensen, M.; Kudsk, P. Fitness of ALS-inhibitors herbicide resistant population of loose silky bentgrass (Apera spica-venti). Front. Plant Sci. 2017, 8, 1660. [Google Scholar] [CrossRef]
- Belz, R.G.; Duke, S.O. Herbicides and plant hormesis. Pest Manag. Sci. 2014, 70, 698–707. [Google Scholar] [CrossRef]
- Pfleeger, T.; Blakeley-Smith, M.; King, G.; Lee, E.H.; Plocher, M.; Olszyk, D. The effects of glyphosate and aminopyralid on a multi-species plant, field trial. Ecotoxicology 2012, 21, 1771–1787. [Google Scholar] [CrossRef] [PubMed]
- Belgers, J.D.M.; Van Lieverlee, R.J.; Van der Plas, L.J.T.; Van den Brink, P.J. Effects of 2,4-D on the growth of nine aquatic macrophytes. Aquat. Bot. 2007, 86, 260–268. [Google Scholar] [CrossRef]
- Beltz, R.G.; Farooq, M.F.; Wagner, J. Does selective hormesis impact herbicide resistance evolution in weeds? ACCase-resistant populations of Alopecurus myosuroides Huds. as a case study. Pest Manag. Sci. 2018, 74, 1880–1891. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G.; Carbonari, C.A.; Duke, S.O. The potential influence of hormesis on evolution of resistance to herbicides. Curr. Opin. Environ. Sci. Health 2022, 27, 100360. [Google Scholar] [CrossRef]
- European Food Safety Authority. Conclusion on the peer review of fenoxaprop-P. EFSA Sci. Rep. 2007, 121, 1–76. [Google Scholar]
- European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment of the active substance pinoxaden. EFSA J. 2013, 11, 3269. [Google Scholar] [CrossRef]
- R Core Team. R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 2 September 2022).
- EFSA PPR Panel (EFSA Panel on Plant Protection Products and their Residues). Scientific Opinion addressing the state of the science on risk assessment of plant protection products for non-target terrestrial plants. EFSA J. 2014, 12, 3800. [Google Scholar] [CrossRef]
- Warne, M.S.J.; Van Dam, R. NOEC and LOEC data should no longer be generated or used. Australas. J. Ecotoxicol. 2008, 14, 1–5. [Google Scholar]
- Azimonti, G.; Galimberti, F.; Marchetto, F.; Menaballi, L.; Ullucci, S.; Pellicioli, F.; Caffi, A.; Ceriani, L.; Ippolito, A.; Moretto, A.; et al. Comparison of NOEC values to EC10/EC20 values, including confidence intervals, in aquatic and terrestrial ecotoxicological risk assessment. EFSA Support. Publ. 2015, 12, 906E. [Google Scholar] [CrossRef] [Green Version]
- Vila-Aiub, M.M.; Gundel, P.E.; Preston, C. Experimental methods for estimation of plant fitness costs associated with herbicide-resistance genes. Weed Sci. 2015, 63, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.; Auteri, D.; Barmaz, S.; Brancato, A.; Brocca, D.; Bura, L.; Carrasco Cabrera, L.; Chiusolo, A.; Court Marques, D.; Crivellente, F.; et al. Conclusion on the peer review of the pesticide risk assessment of the active substance azadirachtin (Margosa extract). EFSA J. 2018, 16, e05234. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Fenoxaprop-P-ethyl | Pinoxaden | ||||
|---|---|---|---|---|---|---|
| ER50 | ER25 | ER10 | ER50 | ER25 | ER10 | |
| AMI | >662.40 (>8 N) | 101.56 | 27.54 | 36.82 | 23.04 | 5.88 |
| AMII | >662.40 | 52.24 | 8.59 | 351.03 | 55.25 | 42.92 |
| AMIII | >662.40 | 65.69 | 59.71 | 203.74 | 188.41 | 174.23 |
| AMIV | >662.40 | >662.40 | >662.40 | 312.78 | 182.08 | 146.13 |
| AMV | >662.40 | >662.40 | 642.61 | 129.34 | 75.62 | 44.88 |
| AMVI | >662.40 | 396.71 | 246.34 | 50.20 | 29.40 | 10.26 |
| AMVII | 600.69 | 583.5 | 536.31 | 86.75 | 62.47 | 44.99 |
| APSI | 341.38 | 337.75 | 148.97 | 36.80 | 24.30 | 10.90 |
| APSII | >662.40 | >662.40 | >662.40 | 291.22 | 160.29 | 88.23 |
| APSIII | 82.51 | 74.33 | 68.97 | 22.32 | 12.94 | 5.26 |
| Population | Fenoxaprop-P-ethyl | Pinoxaden | ||
|---|---|---|---|---|
| ER10 (Biomass) | ERrepro10 | ER10 (Biomass) | ERrepro10 | |
| AMI | 27.54 | 9.18 | 5.88 | 1.96 |
| AMII | 8.59 | 2.86 | 42.92 | 14.31 |
| AMIII | 59.71 | 19.90 | 174.23 | 58.08 |
| AMIV | >662.40 | 220.80 | 146.13 | 48.71 |
| AMV | 642.61 | 214.20 | 44.88 | 14.96 |
| AMVI | 246.34 | 82.11 | 10.26 | 3.42 |
| AMVII | 536.31 | 178.77 | 44.99 | 15.00 |
| APSI | 148.97 | 49.66 | 10.90 | 3.63 |
| APSII | >662.40 | 220.80 | 88.23 | 29.41 |
| APSIII | 68.97 | 22.99 | 5.26 | 1.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wrochna, M.; Stankiewicz-Kosyl, M.; Wińska-Krysiak, M. Stimulation of Early Post-Emergence Growth of Alopecurus myosuroides and Apera spica-venti Following Spray Application of ACCase Inhibitors. Agriculture 2023, 13, 483. https://doi.org/10.3390/agriculture13020483
Wrochna M, Stankiewicz-Kosyl M, Wińska-Krysiak M. Stimulation of Early Post-Emergence Growth of Alopecurus myosuroides and Apera spica-venti Following Spray Application of ACCase Inhibitors. Agriculture. 2023; 13(2):483. https://doi.org/10.3390/agriculture13020483
Chicago/Turabian StyleWrochna, Mariola, Marta Stankiewicz-Kosyl, and Marzena Wińska-Krysiak. 2023. "Stimulation of Early Post-Emergence Growth of Alopecurus myosuroides and Apera spica-venti Following Spray Application of ACCase Inhibitors" Agriculture 13, no. 2: 483. https://doi.org/10.3390/agriculture13020483