1. Introduction
The greatest challenge facing modern agriculture science is maintaining food production, in order to meet the needs of a growing global population without jeopardizing future generations’ access to natural resources. The current level of agricultural intensification has reached a tipping point, with far-reaching, irreversible effects on the global environment and a significant decline in the range of ecosystem services once provided by nature [
1]. Barley (
Hordeum vulgare L.) is the preferred crop in arid regions, particularly among farmers with limited agricultural resources and low income. It is commonly used for human consumption, animal feed, and malting [
2]. As a result of their low glycemic index and high nutritional value, barley crops are currently the subject of worldwide interest [
3]. In comparison to other small grains such as wheat and rice, it is generally believed that barley yields are less susceptible to weather fluctuations [
4]. In Iraq, barley is primarily grown in areas with limited precipitation and rain-fed conditions. As a result of climatic changes, crops such as wheat and rice are competing for a shrinking amount of arable land. Farmers believe that barley can replace some wheat cultivars, so they are cultivating it instead, particularly during this dry year [
5]. In order to improve the yield and growth parameters of barley, agronomists, crop physiologists, and other researchers must urgently discover sustainable techniques and new innovations. Despite the fact that barley is considered to be stress-resistant [
6], its productivity in harsh environmental conditions is negatively impacted by a number of factors, including water limitation, agronomic practices, heat stress, and so on [
7].
Pesticides, nitrates (from nitrogen-rich fertilizers), and phosphorus are the most serious agricultural pollutants. Chemical fertilizers can certainly increase agricultural yield, and they are regarded as a crucial factor influencing the final quality of barley products [
8]. Unfortunately, the use of this type of fertilizer comes at a terrible price, as it degrades the soil and pollutes the environment, in addition to being expensive. The greatest opportunity for expanding food production is to improve yields and quality through the strategic application of mineral and organic fertilizers, plant protection products, and water supply. It is critical that this process is carried out in a manner that is safe for both the environment and consumers [
9]. In recent years, there has been a great deal of focus on identifying various natural (organic) sources of biostimulants for enhancing crop productivity and achieving sustainable agriculture [
10,
11]. There are numerous sources of biostimulants used frequently in agriculture, including humic acid [
12], chitosan and chitin derivatives [
13], seaweed extracts [
14], and plant extracts [
15].
Stimulants are beneficial, but they cannot replace chemical fertilizer in long-term agricultural output. Plant extracts of moringa can either inhibit (at high concentrations) or stimulate (at low concentrations) plant development and growth [
16,
17]. Moringa leaf extract (MLE) obtained from moringa (
Moringa oleifera) is one of the most popular plant biostimulants that can be used as a substitute and natural source of mineral nutrition and fertilizer, because it contains stimulant compounds, such as cytokinins such as zeatin, antioxidants such as ascorbic acid, flavonoid, amino acids, vitamins C and A, and phenolics, and micro- and macronutrients [
18]. Besides, Yasmeen et al. [
19] showed that the leaf extraction of such a plant can also provide the balance among nutrients, phytohormones, and antioxidants. Zeatin is the main hormone detected in MLE, and, so far, its concentration is thousands of times higher compared to the most studied plants [
20]. To improve the productivity and growth of many plants grown in normal conditions, this kind of natural biostimulant has been applied as a foliar application [
21,
22,
23]. To our knowledge, few small-scale experiments have been conducted to investigate the effect of moringa mlant extract on barely crops for productivity [
24]. At field scale, no study has been reported using a combined application of moringa (leaf, root, and seed) extract. Recently, an investigation into a collection of barley accessions grown in Iraq by our own research group stated different patterns of response at early stages, phenotypically, physiologically, and biochemically, for drought tolerance [
25]. In this regard, the current investigation was planned to study the effects of moringa organ extract on the growth and yield of a collection of barley accessions.
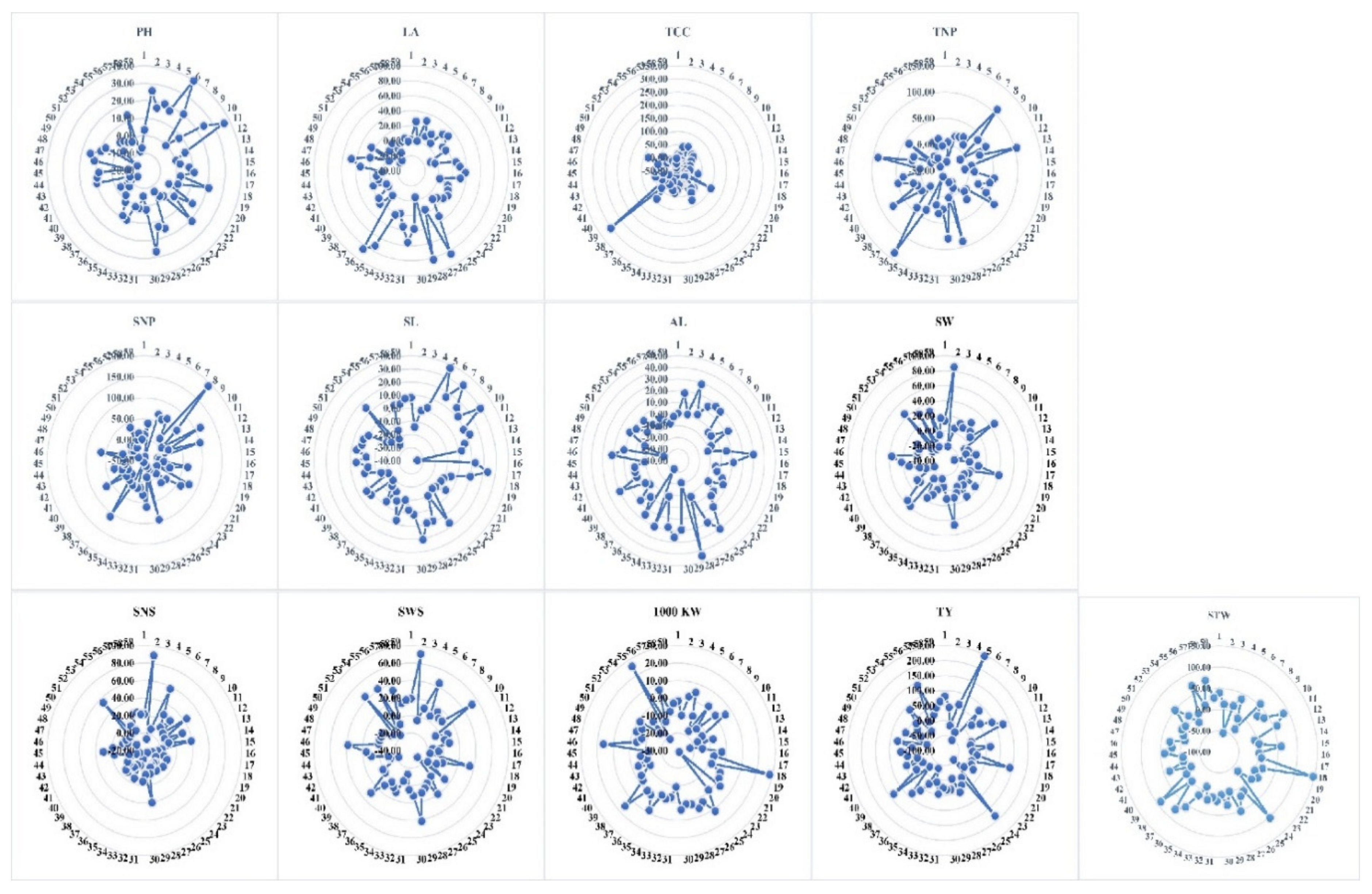
4. Discussion
Plant scientists are now focusing on biostimulants and how to use them in their research to increase crop yields. Plant stimulants have been shown to improve plant health and yield quality by increasing nutrient uptake, changing plant physiology, and making plants more resilient to stress [
19,
22]. The ultimate goal of any breeding strategy is to increase barley and other cereal yields. To increase the yield of contributing factors, several strategies are being implemented. Crop yield in cereals is primarily determined by measuring the most important traits that are strongly related to the final yield product, such as spike length, spike number per plant, seed weight per spike, 1000-kernel weight, seed number per spike, spike weight, total yield, and harvest index. Foliar application of moringa plant extract is well-documented because it is important in improving yield contributing parameters in many plant species [
21,
23,
28,
29]. Moringa leaf extract is measured as one of the essential plant biostimulants due to the presence of phenols, antioxidants, essential nutrients, phytohormones, and ascorbates [
22]. When compared to the respective control conditions, exogenous application of moringa plant part extract had a positive impact on these parameters in our experiment.
In this study, MOE had a significant positive impact on plant height. As previously stated, the moringa plant possesses an abundance of phytohormones, including gibberellin [
30]. Gibberellin’s metabolism and signaling are both essential for controlling plant height. The presence of this phytohormone enhanced internode elongation, leading to an increase in cell division and cell elongation [
31]. Similar to our study, Rehman et al. [
32] discovered a significant increase in wheat height due to the use of moringa extract.
Some traits, such as leaf area [
33], awn length [
34], and chlorophyll content [
35], have been shown to play a major role in increasing photosynthesis under normal and stressful conditions. The primary organ, which takes a huge portion of the energy in photosynthesis, is the flag leaf. The characteristics of flag leaf are considered essential selection criteria for high grain yields in barley [
36]. For this reason, the lower leaves are mostly covered by the upper plant parts and, therefore, do not directly take part in absorbing the radiation of solar energy. After the application of moringa plant extract, a significant increase in leaf area was observed, probably due to the presence of the critical phytohormones in their nature. Several phytohormones with an obvious portion were detected in moringa leaf extract by Ali et al. [
30], including gibberellins, auxin, and cytokinins. It is well-documented that gibberellins improve plant height, while auxins improve the elongation of cells and promote the growth of stems, and cytokinins play a critical role in the promotion of cell division and modification of apical dominance [
37]. In accordance with our findings, Chattha et al. [
38] found similar outcomes in the case of using this type of extract on the wheat plant. Additionally, Ali et al. [
39] showed a significant increase in the measurement of this trait on wheat varieties, after conducting the same exogenous application of moringa leaf extract.
Since chlorophyll is required to convert light energy into stored chemical energy, crop growth and yield are directly affected by chlorophyll content [
40]. Correlations between leaf area, chlorophyll content, and yield were shown by many studies for barley cultivars [
41,
42,
43]. This is probably due to capturing lighter chloroplast, while including a denser chloroplast. New opportunities to predict total chlorophyll content (TCC) at the various crop growth stages have been provided with the development of remote sensing equipment (SPAD), which is widely accepted by researchers [
44,
45]. In the regulation of photosynthesis and many physiological processes, salicylic acid (SA) plays a main role under stress conditions in maintaining these regulations within plant cells [
37]. Until very recently, for barley genotypes, a foliar application of combination gibberellic acid and (SA) with a concentration of (110 mg/l and 1.5 mM) showed a significant increase in different plant physiological properties, including total chlorophyll content [
46]. Many essential developmental processes are modulated by the presence or absence of cytokinins, including leaf development in the last phase, well-known as senescence, which is associated with the breakdown of chlorophyll and photosynthetic collapse. All of these undesirable changes can be slowed by cytokinins [
47]. Taking all the phytohormones present in moringa leaf extract into account, it is possible to conclude that a strong direct correlation is present between the total chlorophyll content and those phytohormones. For all the above reasons, these traits (leaf area and total chlorophyll content) could be used as growth morphological markers for the selection of barley accessions having higher photosynthetic activity.
Cereals have at least two types of tillers (fertile and non-fertile). The first, also known as the productive tiller, causes the formation of spikes and is, thus, necessary for seed yield. The first type depletes the plant’s mineral resources. Since this type of tiller rarely survives until the end of the plant’s life, it cannot produce a yield [
48]. To assess the final productivity of studied cereals, it is critical to measure the fertile tiller number per plant at this point. The obvious increase in tiller number in our results was most likely due to the presence of a cytokinin growth regulator in the moringa plant extract [
30]. As a result, the trait of tiller number can be carefully chosen for studying the application of moringa plant extract. Afzal et al. [
49] and Rehman et al. [
32] reported that the application of moringa leaf extract increased the studied yield traits, including tiller number, in wheat, which is consistent with our findings. In addition to these findings, Koprna et al. [
50], from Palack University Olomouc, stated that cytokinin application has a positive effect on the tiller number of barley varieties.
The number of spikes per plant is one of the most important yield characteristics. The selection of barley genotypes based on the number of spikes per plant may eventually lead to the selection of accessions with better yielding performance among tested accessions. This trait has a significant impact on barley genotype yield [
51]. This could be due to these barley accessions’ ability to respond strongly to this management. In our study, the variation in the number of spikes per plant can be attributed to the genetic potential of barley accessions and their diverse responses to foliar moringa application. In the current foliar treatment, the three barley accessions, AC28, AC47, and AC36, with values of 25.33, 25.00, and 23.67, respectively, had greater potential to produce a large SNP. The current study’s findings are consistent with the findings of another group that investigated the effect of moringa extract on this specific yield trait. Afzal et al. [
49] investigated the effects of three different foliar applications, moringa leaf, sorghum water extract, and salicylic acid, at concentrations of 3%, 0.075%, and 0.01%, respectively, on wheat plants under current heat stress. They applied the foliar application three times in one month, beginning with the tillering stage. Among the tested foliar applications, moringa extract and salicylic acid significantly improved this trait’s performance. Similarly, Khan et al. [
22] demonstrated a significant impact of moringa leaf extract alone and in combination with other plant growth promoters such as ascorbic acid and salicylic acid for this trait on wheat, by administering this treatment twice during the tillering and flowering stages.
The presence of various phytohormones and secondary metabolites in moringa plant parts may be linked to the longer spike length in the current study [
30]. Similarly, Khan et al. [
22] observed a significant increase in spike length on the wheat plant in the field, as a result of using the same application method. Furthermore, Zaheer et al. [
52] studied wheat cultivars using various foliar applications, including cytokinins at 25 mg L
−1 concentrations, used under drought stress conditions at three different growth stages (tiller formation, flowering, and grain filling). The longevity of spikes in their study was significantly improved in the presence of this application. As a result, it is perfectly reasonable to apply moringa foliar to increase spike length.
In our study of the awn length trait, a significant increase was observed when moringa extract was applied foliarly. As a result, increasing awn length could eventually lead to increased barley crop productivity. After the flag leaf, the awns of barley are the most important photosynthetic organs. This organ is the closest plant part to the developing grains in spikelets within the spike, acting as a source of assimilation for grain formation. The photosynthesis of barley spike organs (including awn) accounts for more than 75% of the accumulation of kernel dry weight [
53]. It has been long-established that under normal growth conditions in barley, the awns organ can achieve more than 90% of spike photosynthesis [
54]. As a result, these plant parts can significantly increase the proportion of net photosynthesis, resulting in a higher value of grain dry matter. Awn removal in barley genotypes had a significant effect on grain yield performance, transpiration rate, and net photosynthetic rate, all of which were reduced [
55].
The increased spike weight of plants sprayed with moringa organ extract in our research was due to increased spike length, number of seeds per spike, and other yield-contributing factors previously described. A cheap, rich, and natural source of important secondary metabolic products and plant phytohormones plays an important role in barley trait improvement. The application of the moringa plant part as a foliar spray significantly increased the studied parameters in our study due to the phenomenon of remaining green for a longer period of time during grain filling. This could be due to the high concentration of cytokinin hormone in moringa extract, which is the most general coordinator between senescence and remaining green traits, ultimately improving final yield productivities.
Foliar application of moringa plant part extract had a positive effect on the trait of seed number per spike in tested barley accessions. The grain number and final yield are thought to be positively correlated with the dry weight of the spike during the spike growth phase, possibly due to improved photosynthetic capacity [
56]. Zhang et al. [
57] used CRISPR/Cas9 gene-editing techniques to determine the roles of cytokinin oxidase and dehydrogenase in rice among eleven candidate CKXs families for their effects on grain number, leaf senescence, and regulating the source of leaf and sink of grain. They discovered that OsCKX11 knockout significantly increased cellular cytokine levels, resulting in a delayed leaf senescence phenotype. Furthermore, the mutant OsCKX11 showed a significant increase in grain number, when compared to the wild type. It is possible that OsCKX11 regulates both grain number and photosynthesis. Previous research, as mentioned above, demonstrated the positive regulation of cytokinin in increasing the number of seeds per spike. The significant findings in our study for this trait may be linked to the presence of these essential phytohormones in moringa organ extract. As a consequence, the higher the cytokinin content, the greater the number of seeds detected in this study. It is quite clear that the combination of the activity of particular phytohormones as well as the nutritional condition of the reproductive meristem both have significant effects on final grain number [
48].
MOE application positively affected seed weight per spike in the majority of barley accessions, indicating efficient nutrient use by the plant and translocation of these substrates into reproductive plant parts [
58]. Similar to our results, a considerable increase in the seed weight per pod in pea plants [
59], seeds in maize kernel [
60], and snap bean [
61] was detected due to the treatment of moringa leaf extract.
The majority of barley accessions reacted negatively to the MOE application. This reduction in 1000-KW was caused by producing a large number of seeds with small kernels that were less dense and had a low amount of food reserves, because embryo size and reserved nutrient quantity determine the quantity and quality of the seed [
62].
The study of total yield and straw weight performance for its production is dependent on the genetic characteristics of the cereal crop, the nutrient status of the soil texture, the exogenous application of growth promoters, and the environmental conditions of the crop plants [
22]. Under MOE, we discovered statistically significant positive values for nearly all of the explored yield traits. This is likely due to the presence of cytokinins in moringa leaves, which stimulate carbohydrate metabolism [
29,
63]. In addition, this characteristic creates a new sink source, leading to an increase in dry matter content. From accession to accession, the total yield of cereal grain and the values of its constituents vary. These differences in yield are strictly correlated with variation in grain number and must, therefore, rely on variation in shoot number, which produces more spikes [
64]. In a similar vein, a team of researchers led by Brockman and Brennan [
21] discovered significant results in grain yield and dry biomass when moringa leaf extract was applied to a greenhouse-grown wheat cultivar. In addition to total yield, straw weight is an important trait for plant breeding because it reveals the plant’s capacity to allocate biomass to reproductive plant parts. It is related to grain yield and biomass in accordance with the multiplicative yield component, wherein grain yield is a product of yield biomass and harvest index [
65]. This study’s hypothesis, that moringa plant extract is a significant plant growth enhancer, is supported by the numerous MOE compositions discovered by other researchers as well as by the growth and productivity characteristics exhibited by plants treated with moringa plant extract.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}