
Nitrate Increases Aluminum Toxicity and Accumulation in Root of Wheat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Determination of Al Content of Cell Wall in Root Tips
2.3. Localization of Al in Root Tips
2.4. Membrane Integrity Verification Assay
2.5. Antioxidant Enzyme Activity and Antioxidant Determination
2.6. Cell-Wall Fraction Determination
2.7. Statistical Analysis
3. Results
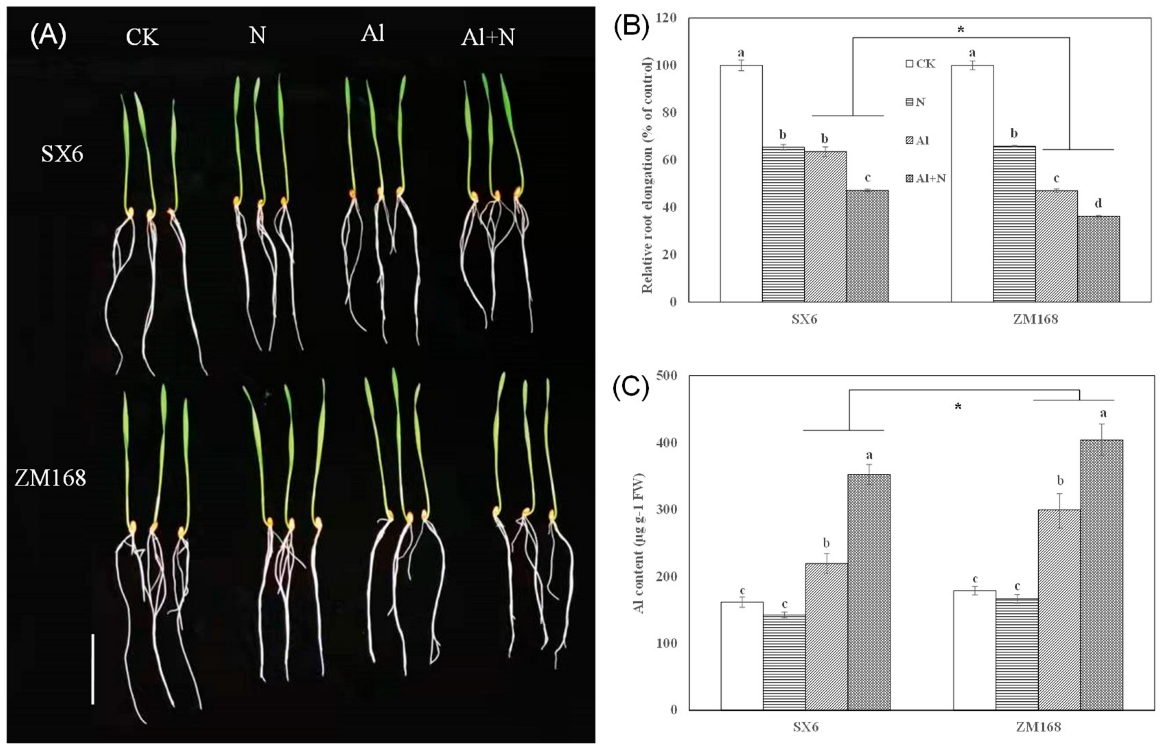
3.1. Effect of N and Al on Root Elongation and Al Accumulation in Root Tips
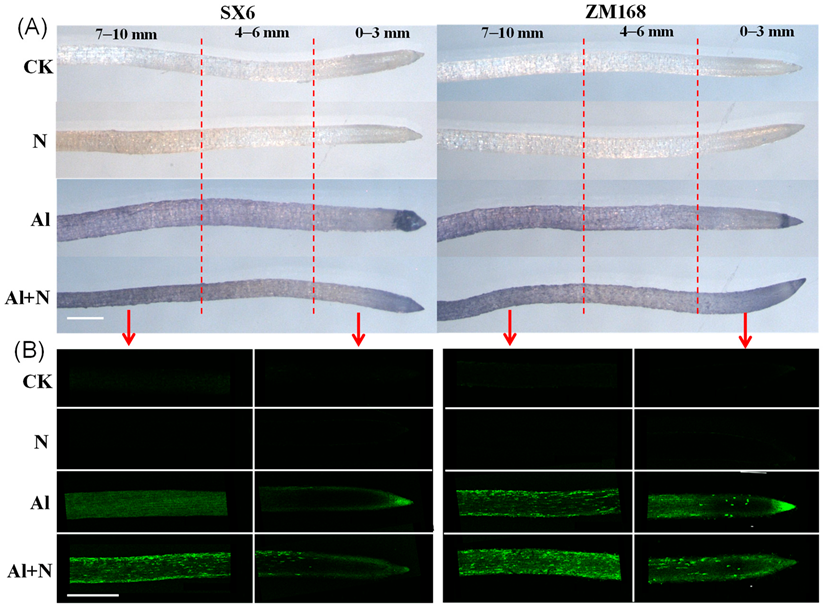
3.2. Al Localization in Root Tips
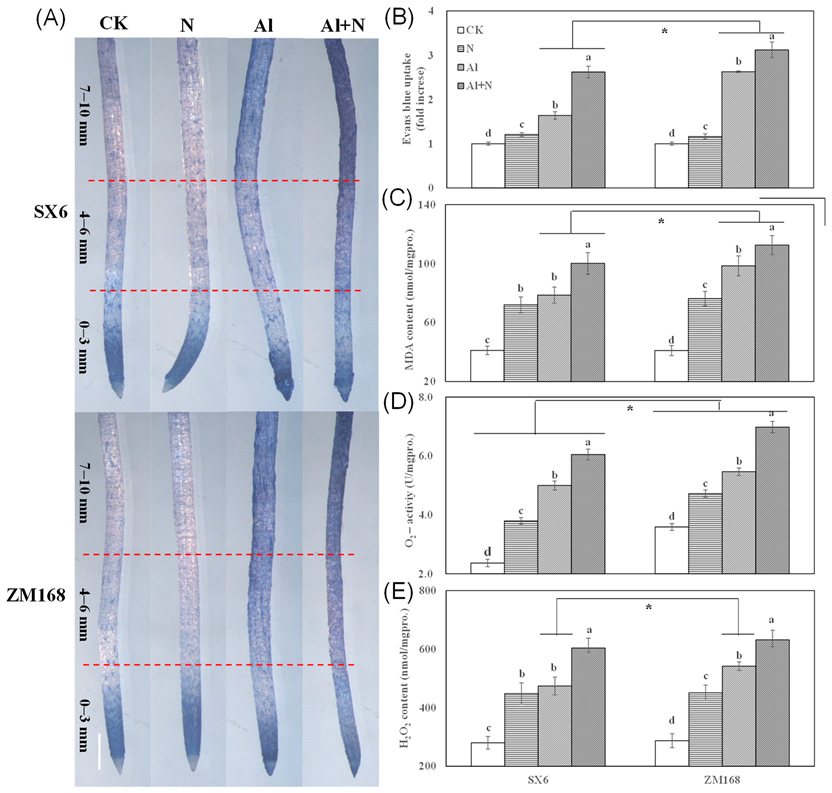
3.3. Oxidative Damage and Reactive Oxygen Species (ROS) in Root Tips
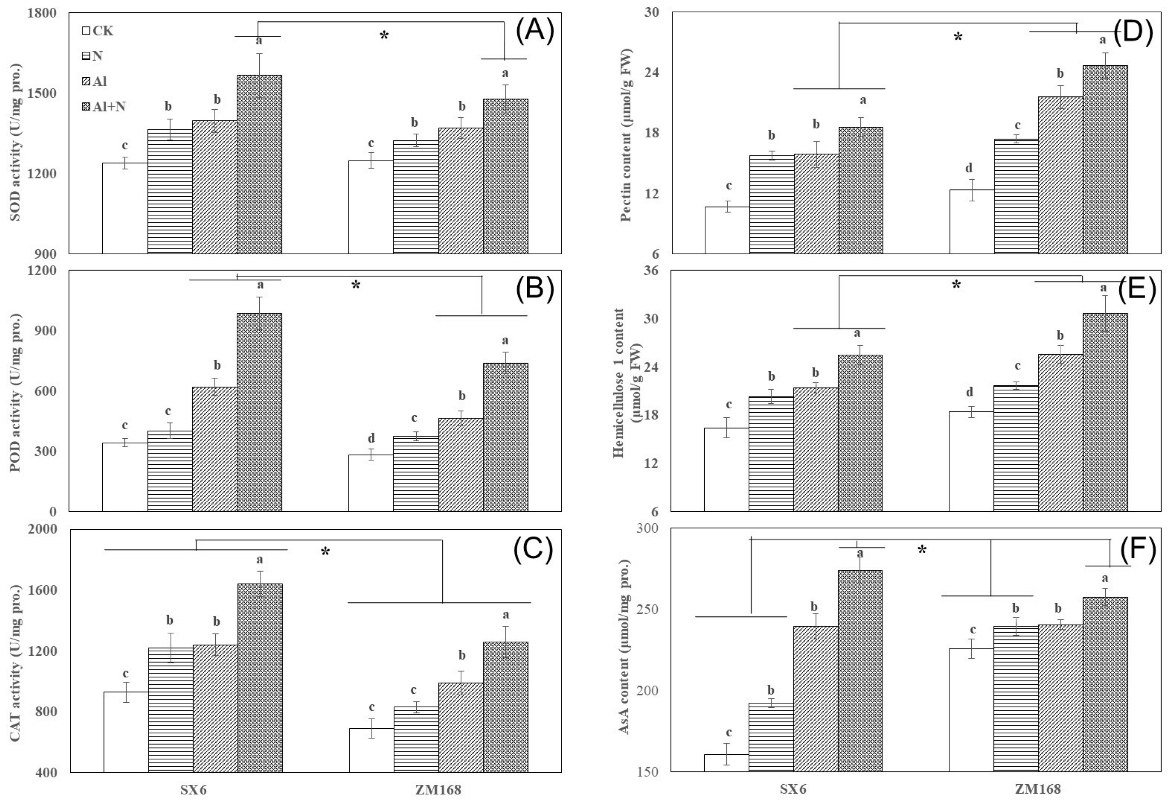
3.4. Antioxidant Defense System and Cell-Wall Fractions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kochian, L.V.; Pineros, M.A.; Liu, J.P.; Magalhaes, J.V. Plant adaptation to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.F.; Zhao, X.Q. The sustainable use of acid soils. J. Agric. Food Chem. 2019, 9, 16–20. [Google Scholar]
- Liu, J.; Piñeros, M.A.; Kochian, L.V. The role of aluminum sensing and signaling in plant aluminum resistance. J. Integr. Plant Biol. 2014, 56, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Li, Y.L.; Mai, J.; Tao, L.; Qu, M.; Liu, J.Y.; Shen, R.F.; Xu, G.; Feng, Y.M.; Xiao, H.D. Boron alleviates aluminium toxicity by promoting root alkalization in transition zone via polar auxin transport. Plant Physiol. 2018, 177, 1254–1266. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Xu, F.J.; Lv, T.; Zhou, W.W.; Chen, Y.; Jin, C.W.; Lu, L.L.; Lin, X.Y. Spatial responses of antioxidative system to aluminum stress in roots of wheat (Triticum aestivum L.) plants. Sci. Total Environ. 2018, 627, 462–469. [Google Scholar] [CrossRef]
- Sun, C.L.; Lv, T.; Huang, L.; Liu, X.X.; Jin, C.W.; Lin, X.Y. Melatonin ameliorates aluminum toxicity through enhancing aluminum exclusion and reestablishing redox homeostasis in roots of wheat. J. Pineal Res. 2020, 68, e12642. [Google Scholar] [CrossRef]
- Ryan, P.R.; Ditomaso, J.M.; Kochian, L.V. Aluminium toxicity in roots: An investigation of spatial sensitivity and the role of the root cap. J. Exp. Bot. 1993, 44, 437–446. [Google Scholar] [CrossRef]
- Sivaguru, M.; Horst, W.J. The distal part of the transition zone is the most aluminum-sensitive apical root zone of maize. Plant Physiol. 1998, 116, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Kollmeier, M.; Dietrich, P.; Bauer, C.S.; Horst, W.J.; Hedrich, R. Aluminum activates a citrate-permeable anion channel in the aluminum-sensitive zone of the maize root apex. A comparison between an aluminum-sensitive and an aluminum-resistant cultivar. Plant Physiol. 2001, 126, 397–410. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F. Syndrome of aluminum toxicity and diversity of aluminum resistance in higher plants. Int. Rev. Cytol. 2007, 264, 225–252. [Google Scholar]
- Rengel, Z.; Zhang, W.H. Role of dynamics of intracellular calcium in aluminium-toxicity syndrome. New Phytol. 2003, 159, 295–314. [Google Scholar] [CrossRef]
- Horst, W.J.; Wang, Y.; Eticha, D. The role of root apoplast in aluminum-induced inhibition of root elongation and in aluminum resistance of plants: A review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Ryan, P.R.; Tyerman, S.D.; Sasaki, T.; Furuichi, T.; Yamamoto, Y.; Zhang, W.H.; Delhaize, E. The identification of aluminium-resistance genes provides opportunities for enhancing crop production on acid soils. J. Exp. Bot. 2011, 62, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.P.; Luo, X.Y.; Shaff, J.; Liang, C.Y.; Jia, X.M.; Li, Z.Y.; Magalhaes, J.; Kochian, L.V. A promoter-swap strategy between the AtALMT and AtMATE genes increased Arabidopsis aluminum resistance and improved carbon-use efficiency for aluminum resistance. Plant J. 2012, 71, 327–337. [Google Scholar] [CrossRef]
- Inostroza-Blancheteau, C.; Rengel, Z.; Alberdi, M.; Mora, M.D.L.L.; Aquea, F.; Arce-Johnson, P.; Reyes-Díaz, M. Molecular and physiological strategies to increase aluminum resistance in plants. Mol. Biol. Rep. 2012, 39, 2069–2079. [Google Scholar] [CrossRef]
- Zang, L.L.; Paven, M.C.M.L.; Clochard, T.; Porcher, A.; Satour, P.; Mojović, M.; Vidović, M.; Limami, A.M.; Montrichard, F. Nitrate inhibits primary root growth by reducing accumulation of reactive oxygen species in the root tip in Medicago truncatula. Plant Physiol. Biochem. 2020, 146, 363–373. [Google Scholar] [CrossRef]
- Celis-Arámburo, T.D.J.; Carrillo-Pech, M.; Castro-Concha, L.A.; Miranda-Ham, M.D.L.; Martínez-Estévez, M.; Echevarría-Machado, I. Exogenous nitrate induces root branching and inhibits primary root growth in Capsicum chinense Jacq. Plant Physiol. Biochem. 2011, 49, 1456–1464. [Google Scholar] [CrossRef]
- Chu, X.Q.; Wang, J.G.; Li, M.Z.; Zhang, S.J.; Gao, Y.Y.; Fan, M.; Han, C.; Xiang, F.N.; Li, G.Y.; Wang, Y.; et al. HBI transcription factor-mediated ROS homeostasis regulates nitrate signal transduction. Plant Cell 2021, 33, 3004–3021. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Shen, R.F.; Sun, Q.B. Ammonium under solution culture alleviates aluminum toxicity in rice and reduces aluminum accumulation in roots compared with nitrate. Plant Soil 2009, 315, 107–121. [Google Scholar] [CrossRef]
- Chang, Y.C.; Yamamoto, Y.; Matsumoto, H. Accumulation of aluminium in the cell wall pectin in cultured tobacco (Nicotiana tabacum L.) cells treated with a combination of aluminium and iron. Plant Cell Environ. 1999, 22, 1009–1017. [Google Scholar] [CrossRef]
- Yang, J.L.; Zhu, X.F.; Peng, Y.X.; Zheng, C.; Li, G.X.; Liu, Y.; Shi, Y.Z.; Zheng, S.J. Cell wall hemicellulose contributes significantly to aluminum adsorption and root growth in Arabidopsis. Plant Physiol. 2011, 155, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, X.Q.; Chen, R.F.; Dong, X.Y.; Lan, P.; Ma, J.F.; Shen, R.F. Altered cell wall properties are responsible for ammonium-reduced aluminum accumulation in rice roots. Plant Cell Environ. 2015, 38, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.L.; Lu, L.L.; Yu, Y.; Liu, L.J.; Hu, Y.; Ye, Y.Q.; Jin, C.W.; Lin, X.Y. Decreasing methylation of pectin caused by nitric oxide leads to higher aluminum binding in cell walls and greater aluminum sensitivity of wheat roots. J. Exp. Bot. 2016, 67, 97. [Google Scholar] [CrossRef] [Green Version]
- Osawa, H.; Matsumoto, H. Possible involvement of protein phosphorylation in aluminum-responsive malate efflux from wheat root apex. Plant Physiol. 2001, 126, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.Y.; Guo, Y.Y.; Cai, S.G.; Kuang, L.H.; Shen, Q.F.; Wu, D.Z.; Zhang, G.P. The zinc finger transcription factor ATF1 regulates aluminum tolerance in barley. J. Exp. Bot. 2020, 71, 6512–6523. [Google Scholar] [CrossRef]
- Baker, C.J.; Mock, N.M. An improved method for monitoring cell death in cell suspension and leaf disc assays using Evans blue. Plant Cell Tissue Organ. Cult. 1994, 39, 7–12. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Silva, S.; Rodriguez, E.; Pinto-Carnide, O.; Martins-Lopes, P.; Matos, M.; Guedes-Pinto, H.; Santos, C. Zonal responses of sensitive vs. tolerant wheat roots during Al exposure and recovery. J. Plant Physiol. 2012, 169, 760–769. [Google Scholar] [CrossRef]
- Klug, B.; Horst, W.J. Spatial characteristics of aluminum uptake and translocation in roots of buckwheat (Fagopyrum esculentum). Physiol. Plant. 2010, 139, 181–191. [Google Scholar] [CrossRef]
- Rangel, A.F.; Rao, I.M.; Horst, W.J. Spatial aluminum sensitivity of root apices of two common bean (Phaseolus vulgaris L.) genotypes with contrasting aluminum resistance. J. Exp. Bot. 2007, 58, 3895–3904. [Google Scholar] [CrossRef] [PubMed]
- Samuels, T.D.; Kucukakyuz, K.; Rincon-Zachary, M. Al partitioning patterns and root growth as related to Al sensitivity and Al tolerance in wheat. Plant Physiol. 1997, 113, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.; Crevecoeur, M.; Penel, C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: Possible interaction with peroxidases. New Phytol. 2007, 174, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Tsukagoshi, H.; Busch, W.; Benfey, P.N. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell 2010, 143, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabaldi, L.A.; Cargnelutti, D.; Gonçalves, J.F.; Pereira, L.B.; Castro, G.Y.; Maldaner, J.; Rauber, R.; Rossato, L.V.; Bisognin, D.A.; Schetinger, M.R.C.; et al. Oxidative stress is an early symptom triggered by aluminum in Al-sensitive potato plantlets. Chemosphere 2009, 76, 1402–1409. [Google Scholar] [CrossRef] [PubMed]
- Delisle, G.; Champoux, M.; Houde, M. Characterization of oxalate oxidase and cell death in Al-sensitive and tolerant wheat roots. Plant Cell Physiol. 2001, 42, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kobayashi, Y.; Devi, S.R.; Rikiishi, S.; Matsumoto, H. Aluminum toxicity is associated with mitochondrial dysfunction and the production of reactive oxygen species in plant cells. Plant Physiol. 2002, 128, 63–72. [Google Scholar] [CrossRef]
- Tamás, L.; Dudíková, J.; Ďurčeková, K.; Huttová, J.; Mistrík, I.; Zelinová, V. The impact of heavy metals on the activity of some enzymes along the barley root. Environ. Exp. Bot. 2008, 62, 86–91. [Google Scholar] [CrossRef]
- Giannakoula, A.; Moustakas, M.; Syros, T.; Yupsanis, T. Aluminum stress induces upregulation of an efficient antioxidant system in the Al-tolerant maize line but not in the Al-sensitive line. Environ. Exp. Bot. 2010, 67, 487–494. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.F.; Shi, Y.Z.; Lei, G.J.; Fry, S.C.; Zhang, B.C.; Zhou, Y.H.; Braam, J.; Jiang, T.; Xu, X.Y.; Mao, C.Z.; et al. XTH31, encoding an in vitro XEH/ XET-active enzyme, regulates aluminum sensitivity by modulating in vivo XET action, cell wall xyloglucan content, and aluminum binding capacity in Arabidopsis. Plant Cell 2012, 24, 4731–4747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Shen, R.; Nagao, S.; Tanimoto, E. Aluminum targets elongating cells by reducing cell wall extensibility in wheat roots. Plant Cell Physiol. 2004, 45, 583–589. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Bai, C.; Zhang, X.; Ding, Y. Nitrate Increases Aluminum Toxicity and Accumulation in Root of Wheat. Agriculture 2022, 12, 1946. https://doi.org/10.3390/agriculture12111946
Ma Y, Bai C, Zhang X, Ding Y. Nitrate Increases Aluminum Toxicity and Accumulation in Root of Wheat. Agriculture. 2022; 12(11):1946. https://doi.org/10.3390/agriculture12111946
Chicago/Turabian StyleMa, Yan, Caihong Bai, Xincheng Zhang, and Yanfeng Ding. 2022. "Nitrate Increases Aluminum Toxicity and Accumulation in Root of Wheat" Agriculture 12, no. 11: 1946. https://doi.org/10.3390/agriculture12111946