
Heat Shock Treatment Promoted Callus Formation on Postharvest Sweet Potato by Adjusting Active Oxygen and Phenylpropanoid Metabolism
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
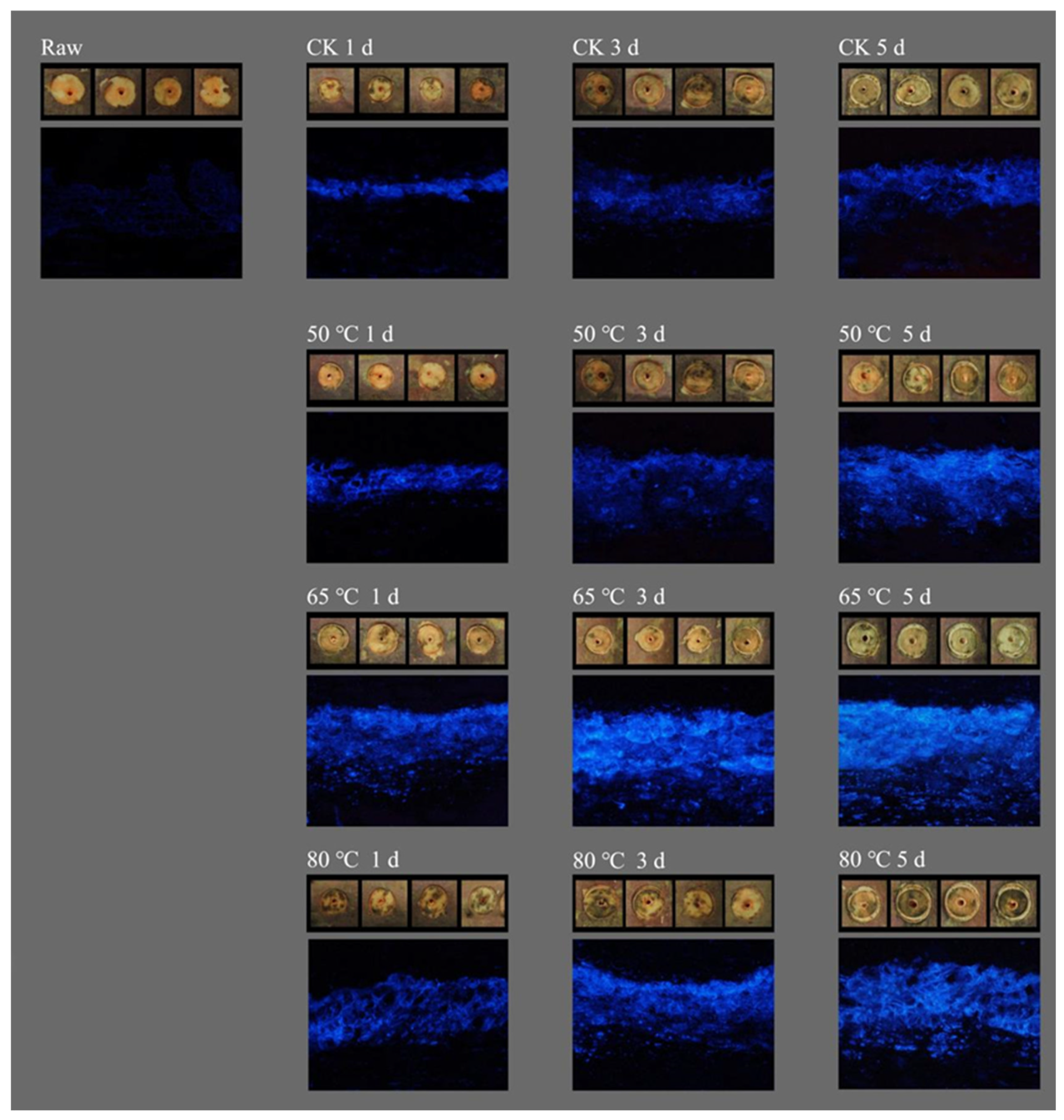
2.1. Effect of RHT Treatment on Callus Wound Healing in Sweet Potato
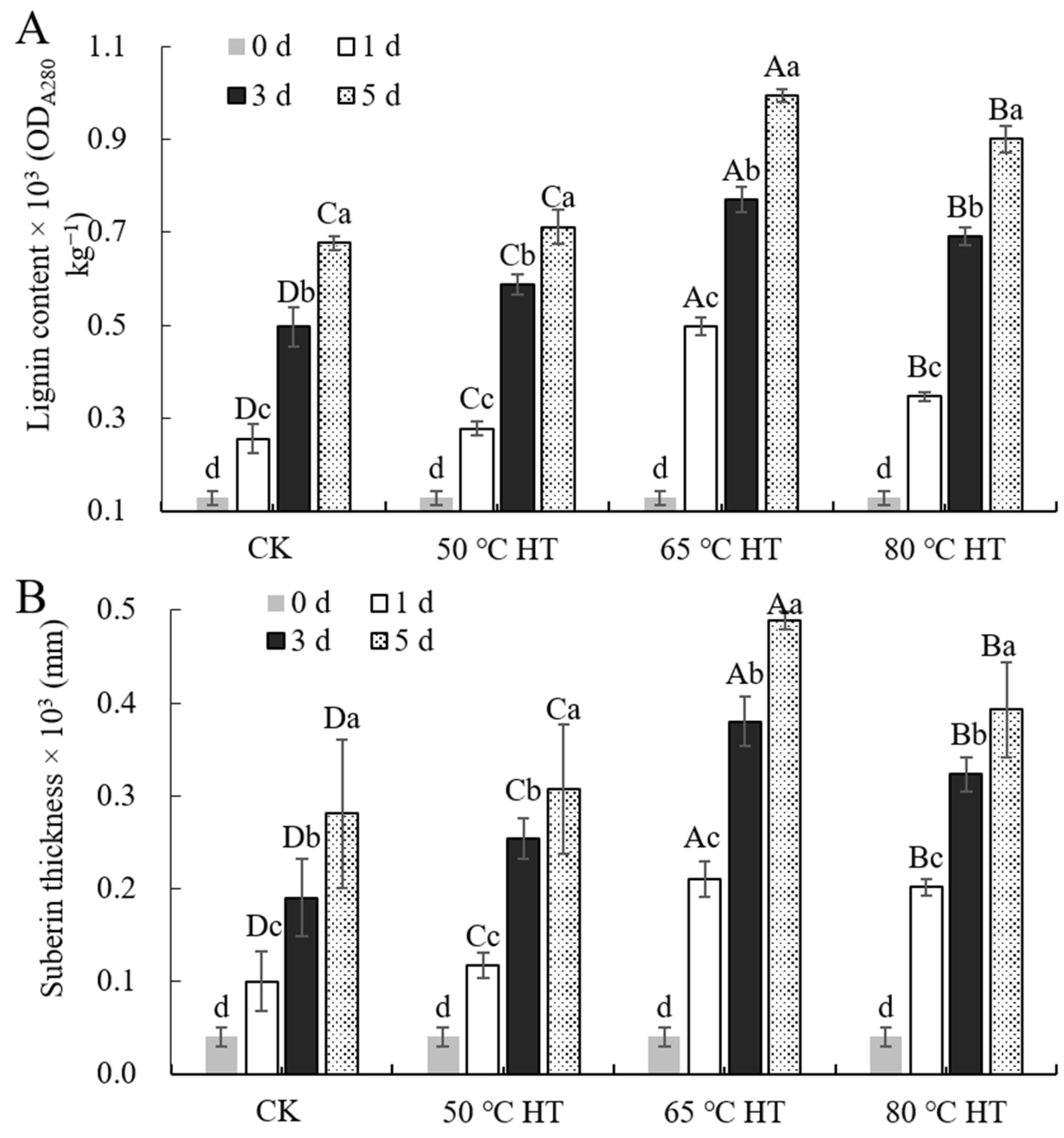
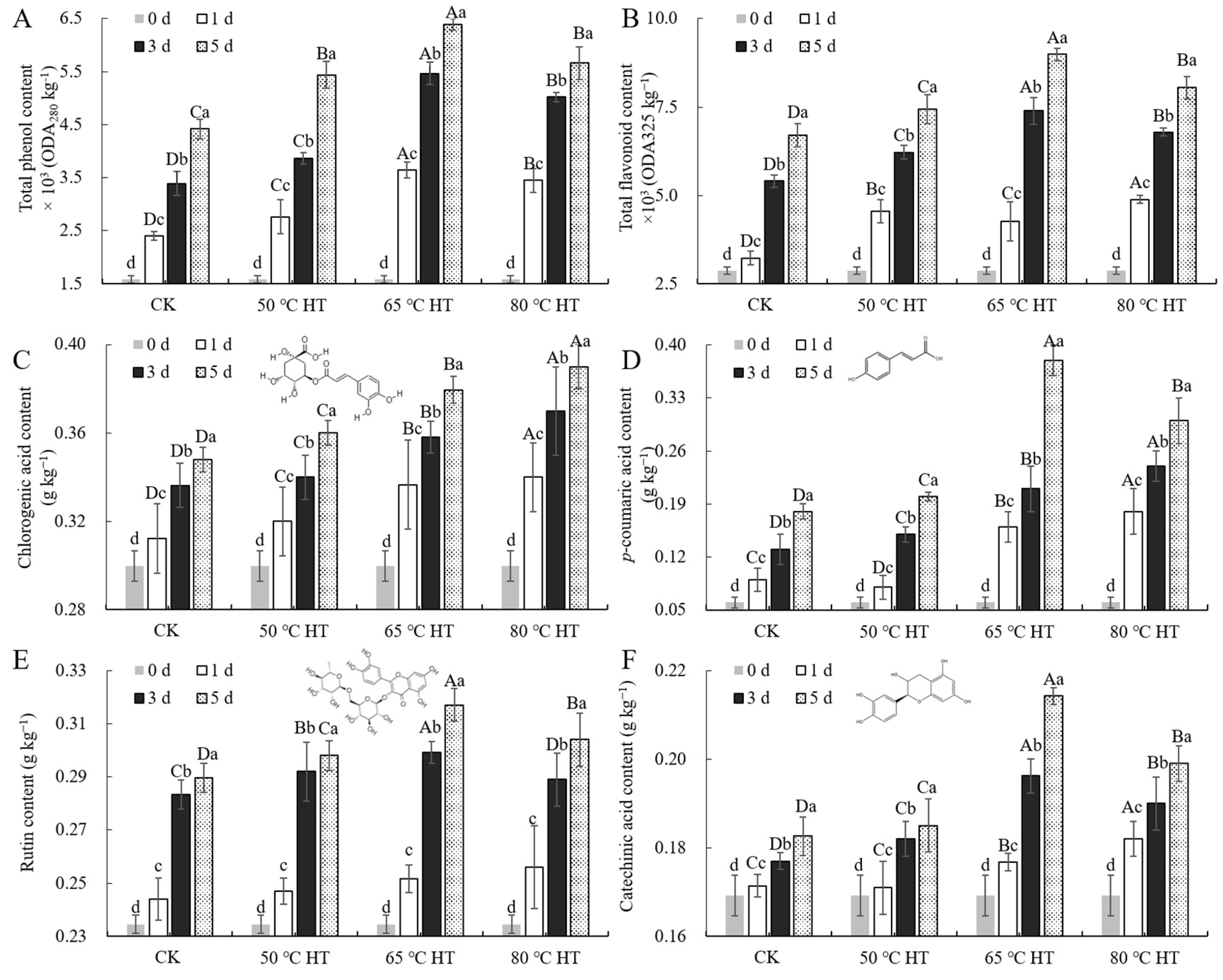
2.2. Effects of RHT Treatment on Phenolic Compounds in Sweet Potato Callus
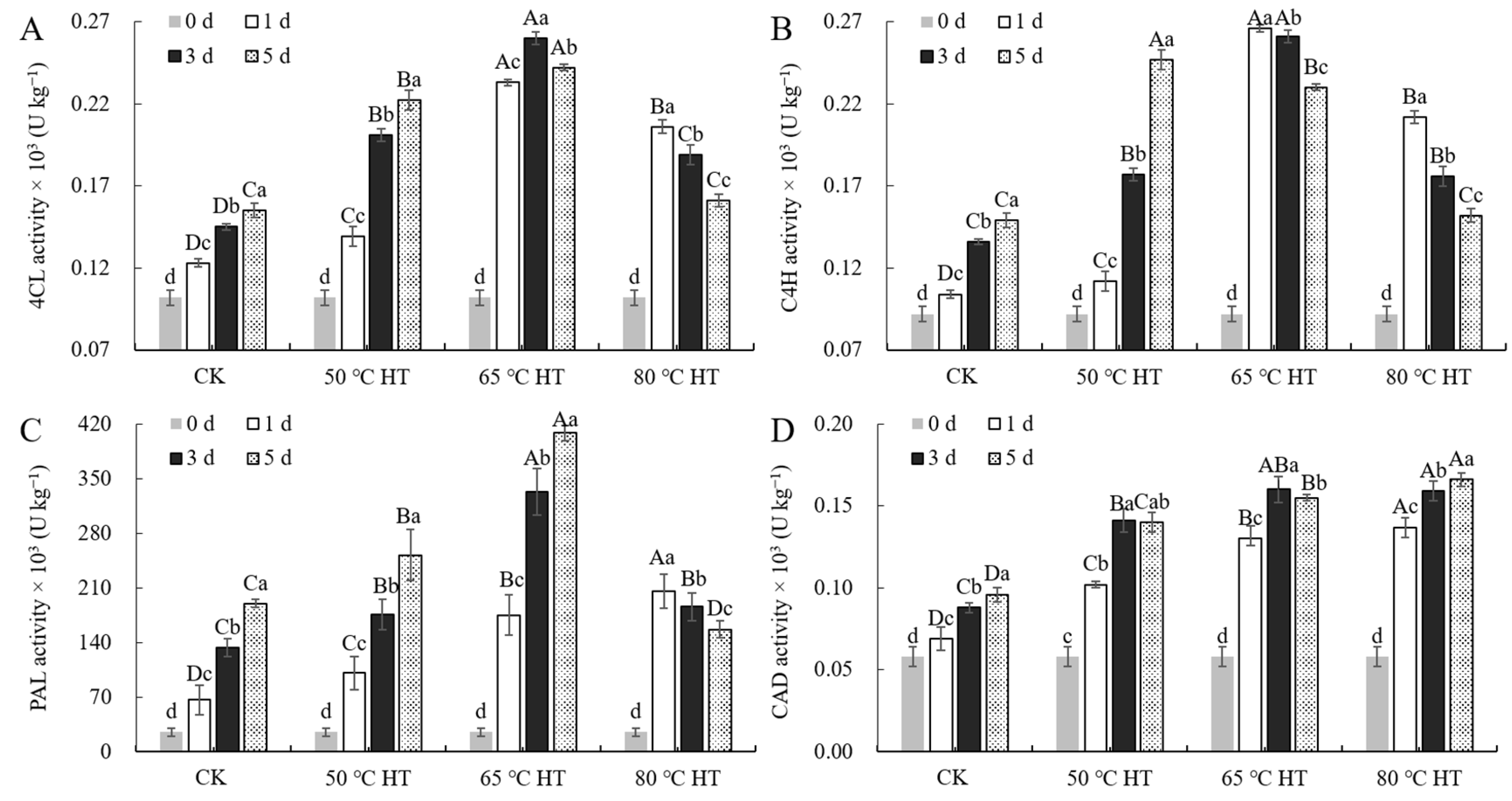
2.3. Effects of RHT Treatment on Phenylpropanoid Metabolic Enzyme Activity during Sweet Potato Callus Healing
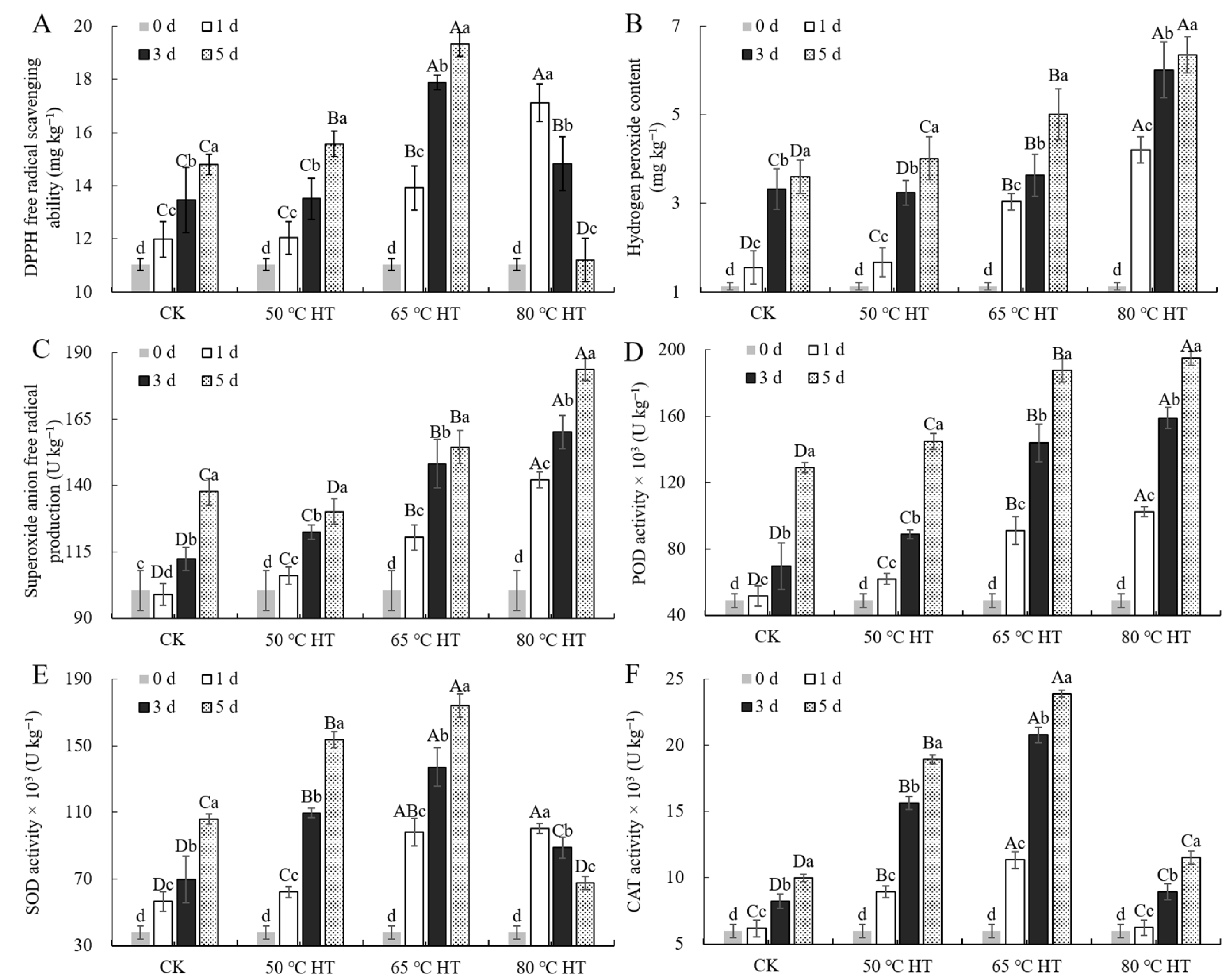
2.4. Effects of RHT Treatment on ROS Production and Scavenging in Sweet Potato Callus
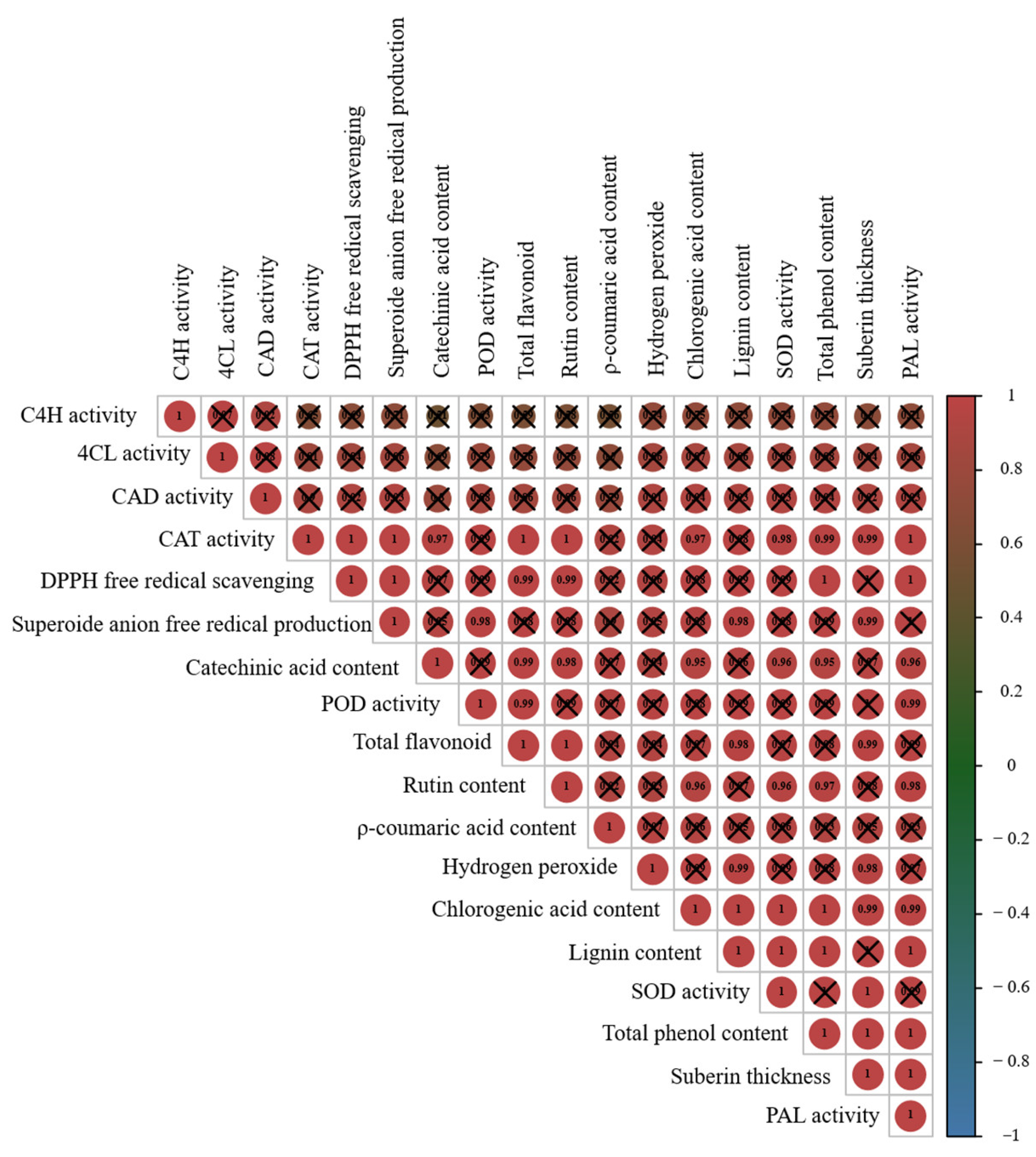
2.5. Pearson’s Correlation Analysis of Factors Affecting Sweet Potato Callus
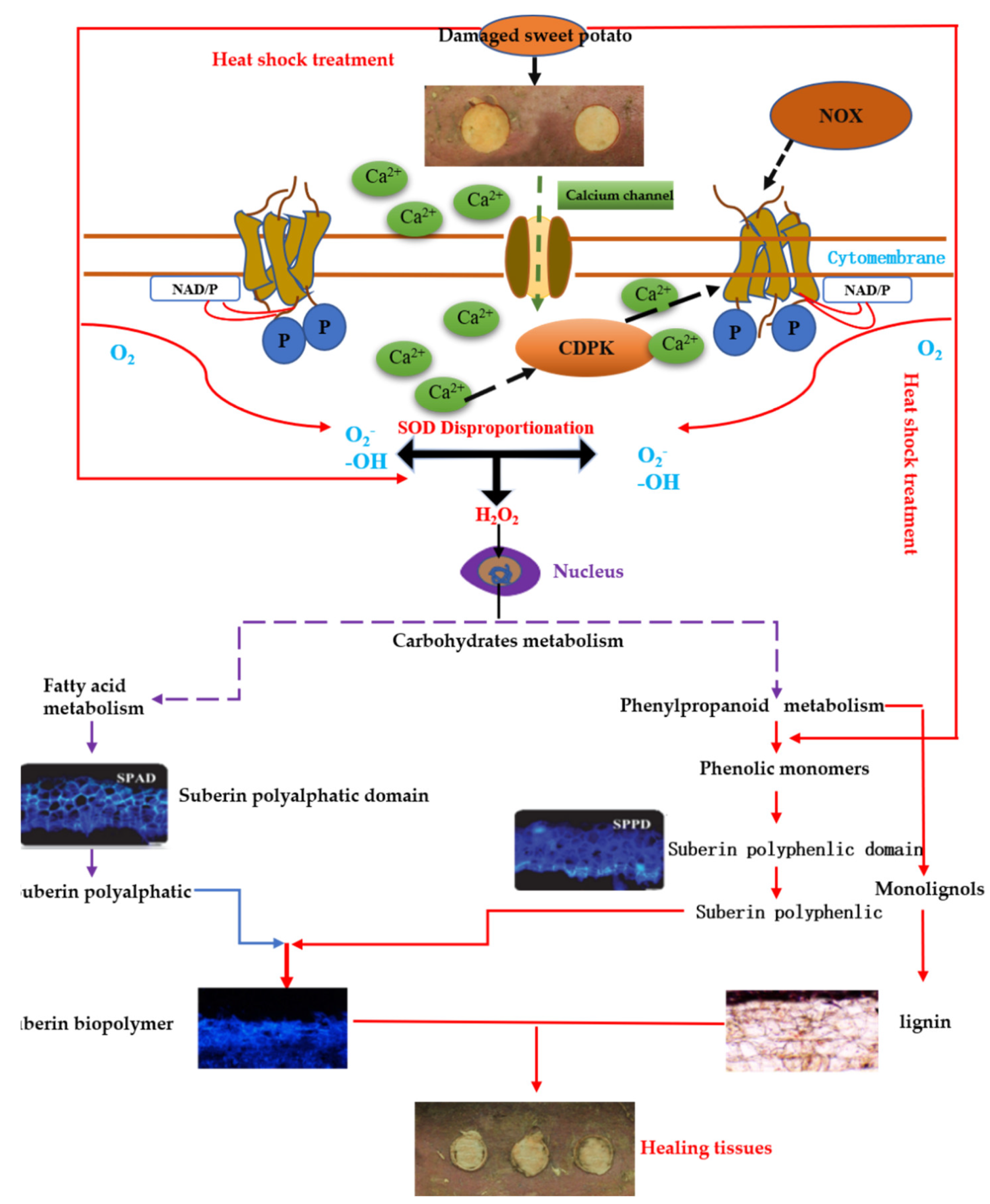
3. Discussion
4. Materials and Methods
4.1. Sweet Potato Acquisition and Sample Preparation
4.2. Artificial Injury and Heat Shock Treatment of Sweet Potato Samples
4.3. Lignin and Suberin Accumulation in Wounded Tissue
4.4. Determination of Lignin Content in Wound Callus of Sweet Potato
4.5. Determination of Phenylpropanoid Metabolism-Related Enzyme Activities in Sweet Potato Callus
4.6. Determination of Antioxidant Capacity and ROS Metabolism-Related Enzymes of Sweet Potato Callus
4.7. Determination of Phenols, Hydrogen Peroxide, and Superoxide Anion in Sweet Potato Callus
4.8. Data Statistics and Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Pazos, J.V.; Rosero, A.; Martínez, R.; Pérez, J.; Morelo, J.; Araujo, H.; Burbano-Erazo, E. Influence of morpho-physiological traits on root yield in sweet potato (Ipomoea batatas Lam.) genotypes and its adaptation in a sub-humid environment. Sci. Hortic. 2020, 275, 109703. [Google Scholar] [CrossRef]
- Ji, C.Y.; Kim, H.S.; Lee, C.-J.; Kim, S.-E.; Lee, H.-U.; Nam, S.-S.; Li, Q.; Ma, D.-F.; Kwak, S.-S. Comparative transcriptome profiling of tuberous roots of two sweetpotato lines with contrasting low temperature tolerance during storage. Gene 2019, 727, 144244. [Google Scholar] [CrossRef] [PubMed]
- Parmar, A.; Kirchner, S.M.; Sturm, B.; Hensel, O. Pre-harvest Curing: Effects on Skin Adhesion, Chemical Composition and Shelf-life of Sweetpotato Roots under Tropical Conditions. East Afr. Agric. For. J. 2017, 82, 130–143. [Google Scholar] [CrossRef]
- Xin, Q.; Sun, J.; Feng, X.; Zhao, Z.; Liu, B.; Jiang, L.; Hao, G. Heat Shock Treatment Promotion Wound Curing and Metabolic Mechanism of Sweet Potato. Food Sci. 2022. (In Chinese with Abstract). [Google Scholar] [CrossRef]
- Xue, W.; Liu, N.; Zhang, T.; Li, J.; Chen, P.; Yang, Y.; Chen, S. Substance metabolism, IAA and CTK signaling pathways regulating the origin of embryogenic callus during dedifferentiation and redifferentiation of cucumber cotyledon nodes. Sci. Hortic. 2021, 293, 110680. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, B.; Ma, L.; Zheng, X.; Gong, D.; Xue, H.; Bi, Y.; Wang, Y.; Zhang, Z.; Prusky, D. Benzo-(1, 2, 3)-thiadiazole-7-carbothioic acid s-methyl ester (BTH) promotes tuber wound healing of potato by elevation of phenylpropanoid metabolism. Postharvest Biol. Technol. 2019, 3, 125–132. [Google Scholar] [CrossRef]
- Dastmalchi, K.; Rodriguez, M.P.; Lin, J.; Yoo, B.; Stark, R.E. Temporal resistance of potato tubers: Antibacterial assays and metabolite profiling of wound-healing tissue extracts from contrasting cultivars. Phytochemistry 2018, 159, 75–89. [Google Scholar] [CrossRef]
- Woolfson, K.N.; Bjelica, A.; Haggitt, M.L.; Kachura, A.; Zhang, Y.N. Differential induction of polar and non-polar metabolism during wound-induced suberization in potato (Solanum tuberosum L.) tubers. Plant J. 2018, 93, 931–942. [Google Scholar] [CrossRef]
- Kumar, G.N.M.; Lulai, E.C.; Suttle, J.C.; Knowles, N.R. Age-induced loss of wound-healing ability in potato tubers is partly regulated by ABA. Planta 2010, 232, 1433–1445. [Google Scholar] [CrossRef]
- Feng, X.; Hansen, J.D.; Biasi, B.; Tang, J.; Mitcham, E.J. Use of hot water treatment to control codling moths in harvested California ‘Bing’ sweet cherries. Postharvest Biol. Technol. 2004, 31, 41–49. [Google Scholar] [CrossRef]
- Jakubowski, T. Use of UV-C radiation for reducing storage losses of potato tubers. Bangladesh J. Bot. 2018, 47, 533–537. [Google Scholar] [CrossRef]
- Lulai, E.C.; Suttle, J.C.; Pederson, S.M. Regulatory involvement of abscisic acid in potato tuber wound healing. J. Exp. Bot. 2008, 59, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Lobato, M.C.; Daleo, G.R.; Andreu, A.B.; Olivieri, F.P. Cell Wall Reinforcement in the Potato Tuber Periderm After Crop Treatment with Potassium Phosphite. Potato Res. 2017, 61, 19–29. [Google Scholar] [CrossRef]
- Goffner, D.; Joffroy, I.; Grima-Pettenati, J.; Halpin, C.; Knight, M.E.; Schuch, W.; Boudet, A.M. Purification and characterization of isoforms of cinnamyl alcohol dehydrogenase from Eucalyptus xylem. Planta 1992, 188, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Mwanga, R.; Andrade, M.; Carey, E.; Low, J.; Yencho, C.; Grüneberg, W. Sweetpotato (Ipomoea batatas L.). In Genetic Improvement of Tropical Crops; Springer: Cham, Switzerland, 2017; pp. 181–218. [Google Scholar] [CrossRef]
- St. Amand, P.C.; Randle, W.M. Ethylene production as a possible indicator of wound healing in roots of several sweet potato. cutivars. Euphytica 1991, 53, 97–102. [Google Scholar] [CrossRef]
- Ge, X.; Zhu, Y.; Li, Z.; Bi, Y.; Yang, J.; Zhang, J.; Prusky, D. Preharvest multiple fungicide stroby sprays promote wound healing of harvested potato tubers by activating phenylpropanoid metabolism. Postharvest Biol. Technol. 2021, 171, 111328. [Google Scholar] [CrossRef]
- Wang, B.; Jiang, H.; Bi, Y.; He, X.; Wang, Y.; Li, Y.; Zheng, X.; Prusky, D. Preharvest multiple sprays with sodium nitroprusside promote wound healing of harvested muskmelons by activation of phenylpropanoid metabolism. Postharvest Biol. Technol. 2019, 158, 110988. [Google Scholar] [CrossRef]
- Zhou, F.H.; Jiang, A.L.; Feng, K.; Gu, S.T.; Xu, D.Y.; Hu, W.Z. Effect of methyl jasmonate on wound healing and resistance in fresh-cut potato cubes. Postharvest Biol. Technol. 2019, 157, 110958. [Google Scholar] [CrossRef]
- Fugate, K.K.; Ribeiro, W.S.; Lulai, E.C.; Deckard, E.L.; Finger, F.L. Cold Temperature Delays Wound Healing in Postharvest Sugarbeet Roots. Front Plant Sci. 2016, 7, 499. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.Y.; Jiang, H.; Bi, Y.; Wang, B.; Wang, T.L.; Li, Y.C.; Gong, D.; Wei, Y.N.; Li, Z.C.; Prusky, D. Comparison of wound healing abilities of four major cultivars of potato tubers in China. Postharvest Biol. Technol. 2020, 164, 111167. [Google Scholar] [CrossRef]
- Irungu, F.G.; Tanga, C.M.; Ndiritu, F.G.; Mathenge, S.G.; Kiruki, F.G.; Mahungu, S.M. Enhancement of potato (Solanum tuberosum L.) postharvest quality by use of magnetic fields—A case of shangi potato variety. Appl. Food Res. 2022, 2, 100191. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, X.; Jiang, H.; Zhao, S.; Silvy, E.M.; Yang, R.; Han, Y.; Bi, Y.; Prusky, D. Kloeckera apiculata promotes the healing of potato tubers by rapidly colonizing and inducing phenylpropanoid metabolism and reactive oxygen species metabolism. Postharvest Biol. Technol. 2022, 192, 112033. [Google Scholar] [CrossRef]
- Meng, G.; Fan, W.; Rasmussen, S.K. Characterisation of the class III peroxidase gene family in carrot taproots and its role in anthocyanin and lignin accumulation. Plant Physiol. Biochem. 2021, 167, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Jiang, F.; Zhou, R.; Wen, J.; Cui, S.; Wang, W.; Wu, Z. Respiratory burst oxidase homologue-dependent H2O2 is essential during heat stress memory in heat sensitive tomato. Sci. Hortic. 2019, 258, 108777. [Google Scholar] [CrossRef]
- Vreugdenhil, D.; Bradshaw, J.E.; Gebhardt, C.; Govers, F.; MacKerron, D.K.L.; Taylor, M.; Ross, H.A. Potato Biology and Biotechnology: Advances and Perspectives; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Liu, B.; Lyu, X.; Wang, C.; Sun, J.; Jiang, W. Process optimization of high temperature short-time hot air treatment to promote sweet potato callus. Trans. Chin. Soc. Agric. Eng. 2020, 36, 313–322, (In Chinese with Abstract). [Google Scholar] [CrossRef]
- Liang, W.; Zhu, Y.; Chai, X.; Kong, R.; Li, B.; Li, Y.; Bi, Y.; Prusky, D. p-Coumaric Acid Promoted Wound Healing of Potato Tubers by Accelerating the Deposition of Suberin Poly Phenolic and Lignin at Wound Sites. Sci. Agric. Sin. 2021, 54, 4434–4445. [Google Scholar]
- Razem, F.A.; Bernards, M.A. Reactive oxygen species production in association with suberization: Evidence for an NADPH-dependent oxidase. J. Exp. Bot. 2003, 54, 935–941. [Google Scholar] [CrossRef]
- Chai, X.; Kong, R.; Zheng, X.; Zhu, Y.; Liang, W.; Zhao, S.; Li, B.; Bi, Y.; Prusky, D. Chlorine dioxide accelerates the wound healing process of potato tubers by eliciting phenylpropanoid metabolism. Food Sci. 2022, 1–11. Available online: http://kns.cnki.net/kcms/detail/11.2206.TS.20220321.1634.034.html (accessed on 22 March 2022). (In Chinese with Abstract).
- Van Oirschot, Q.E.A.; Rees, D.; Aked, J.; Kihurani, A. Sweetpotato cultivars differ in efficiency of wound healing. Postharvest Biol. Technol. 2006, 42, 65–74. [Google Scholar] [CrossRef]
- Lulai, E.C. Skin-Set, Wound Healing, and Related Defects. In Potato Biology and Biotechnology; Elsevier: Amsterdam, The Netherlands, 2007; Chapter 22; pp. 471–500. [Google Scholar]
- Zhou, F.; Xu, D.; Liu, C.; Chen, C.; Tian, M.; Jiang, A. Ascorbic acid treatment inhibits wound healing of fresh-cut potato strips by controlling phenylpropanoid metabolism. Postharvest Biol. Technol. 2021, 181, 111644. [Google Scholar] [CrossRef]
- Vanholme, R.; De Meester, B.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotechnol. 2019, 56, 230–239. [Google Scholar] [CrossRef]
- dos Santos, A.B.; Bottcher, A.; Vicentini, R.; Mayer, J.L.S.; Kiyota, E.; Landell, M.A.; Creste, S.; Mazzafera, P. Lignin biosynthesis in sugarcane is affected by low temperature. Environ. Exp. Bot. 2015, 120, 31–42. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Soler, M.; Serra, O.; Fluch, S.; Molinas, M.; Figueras, M. A potato skin SSH library yields new candidate genes for suberin biosynthesis and periderm formation. Planta 2011, 233, 933–945. [Google Scholar] [CrossRef]
- Sarni, F.; Grand, C.; Boudet, A.M. Purification and properties of cinnamoyl-CoA reductase and cinnamyl alcohol dehydrogenase from poplar stems (Populus × euramericana). Eur. J. Biochem. 1984, 139, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Han, Y.; Han, Z.; Ackah, S.; Li, Z.; Bi, Y.; Yang, Q.; Prusky, D. Hot water dipping stimulated wound healing of potato tubers. Postharvest Biol. Technol. 2020, 167, 111245. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, H.; Fan, X.; Jiao, W.; Cao, J.; Jiang, W. Near freezing point temperature storage inhibits chilling injury and enhances the shelf life quality of apricots following long-time cold storage. J. Food Process. Preserv. 2019, 43, e13958. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xin, Q.; Liu, B.; Sun, J.; Fan, X.; Li, X.; Jiang, L.; Hao, G.; Pei, H.; Zhou, X. Heat Shock Treatment Promoted Callus Formation on Postharvest Sweet Potato by Adjusting Active Oxygen and Phenylpropanoid Metabolism. Agriculture 2022, 12, 1351. https://doi.org/10.3390/agriculture12091351
Xin Q, Liu B, Sun J, Fan X, Li X, Jiang L, Hao G, Pei H, Zhou X. Heat Shock Treatment Promoted Callus Formation on Postharvest Sweet Potato by Adjusting Active Oxygen and Phenylpropanoid Metabolism. Agriculture. 2022; 12(9):1351. https://doi.org/10.3390/agriculture12091351
Chicago/Turabian StyleXin, Qi, Bangdi Liu, Jing Sun, Xinguang Fan, Xiangxin Li, Lihua Jiang, Guangfei Hao, Haisheng Pei, and Xinqun Zhou. 2022. "Heat Shock Treatment Promoted Callus Formation on Postharvest Sweet Potato by Adjusting Active Oxygen and Phenylpropanoid Metabolism" Agriculture 12, no. 9: 1351. https://doi.org/10.3390/agriculture12091351