
Stoichiometric Ratios of Carbon, Nitrogen and Phosphorus of Shrub Organs Vary with Mycorrhizal Type

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Investigation
2.2. Determination of Carbon, Nitrogen and Phosphorus Concentrations
2.3. Data Analysis
3. Results
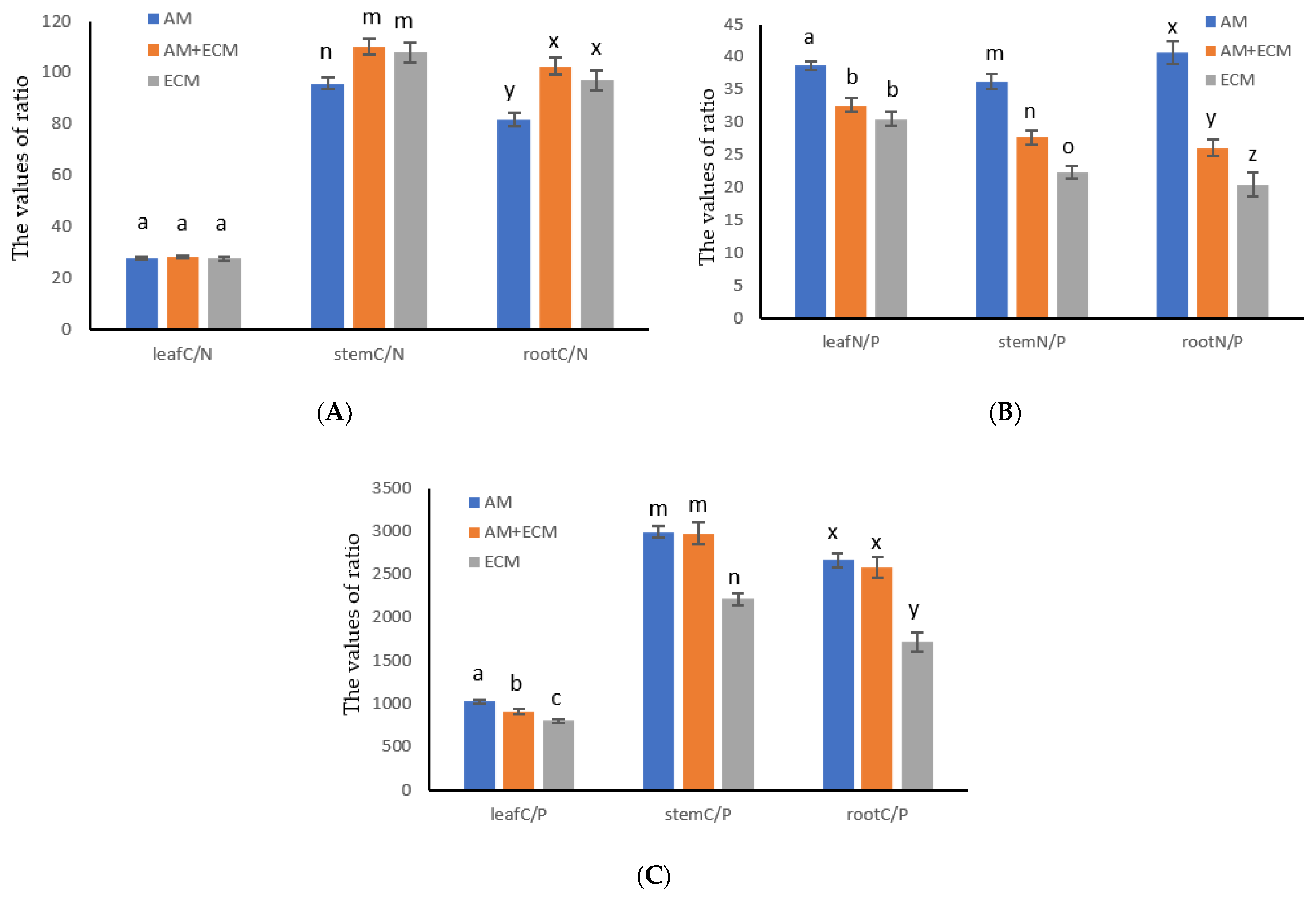
3.1. Elemental Concentration Ratios in Shrub Organs in Different Mycorrhizal Types
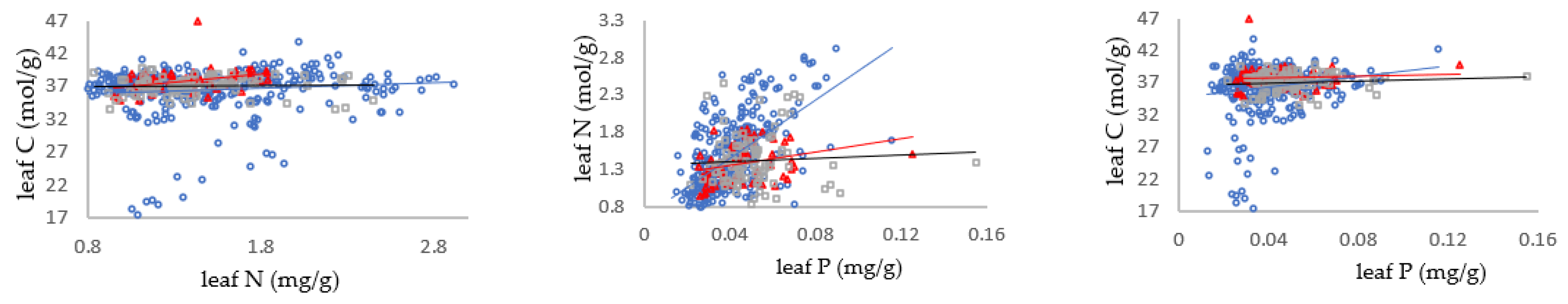
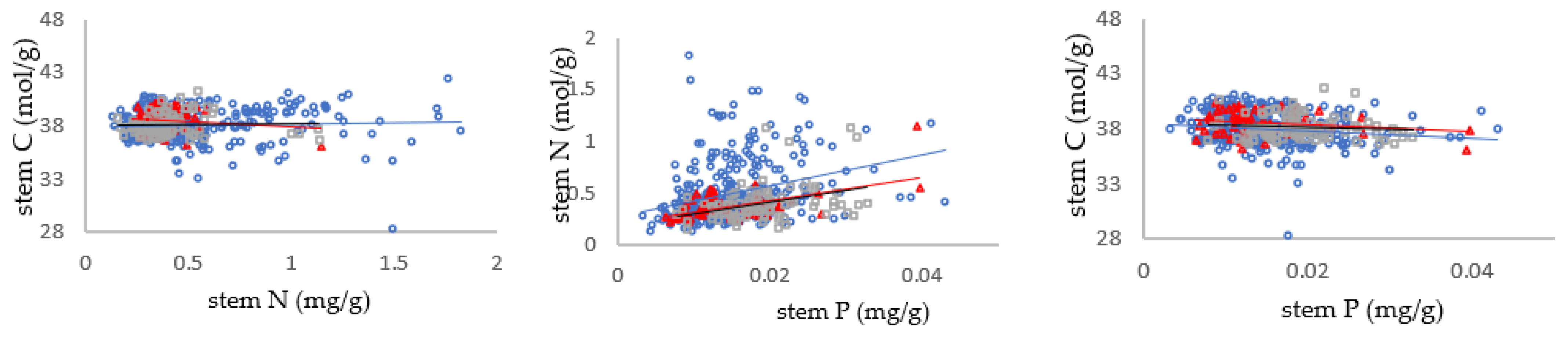
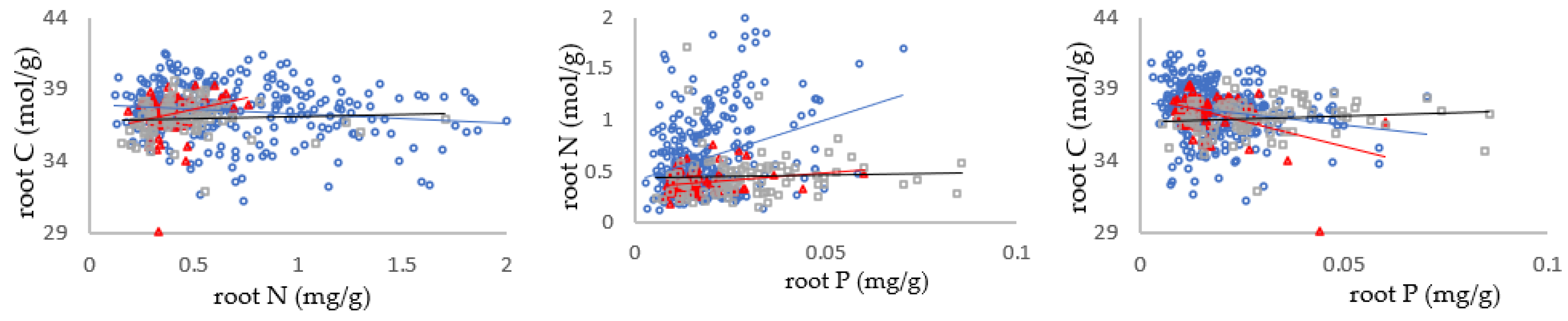
3.2. Relationship between Carbon, Nitrogen and Phosphorus in Different Mycorrhizal Types of Shrub Leaves
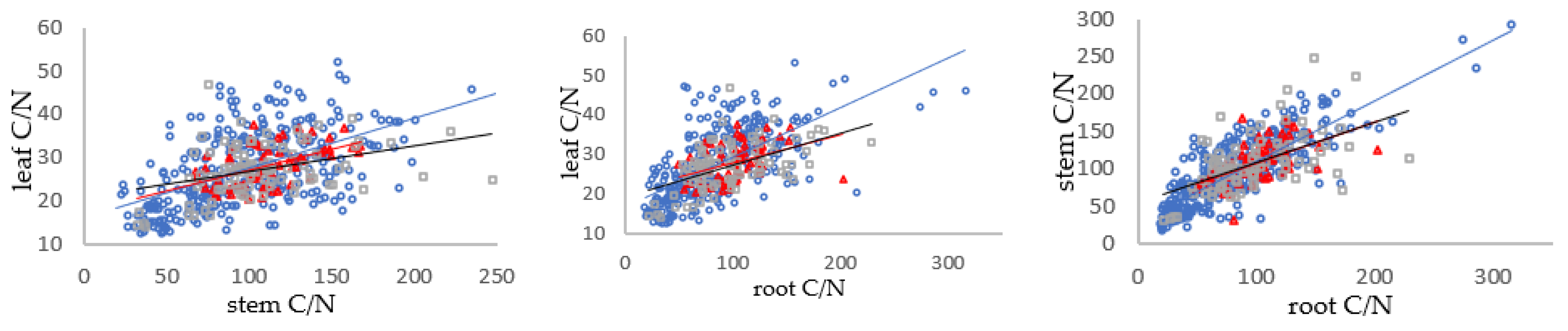
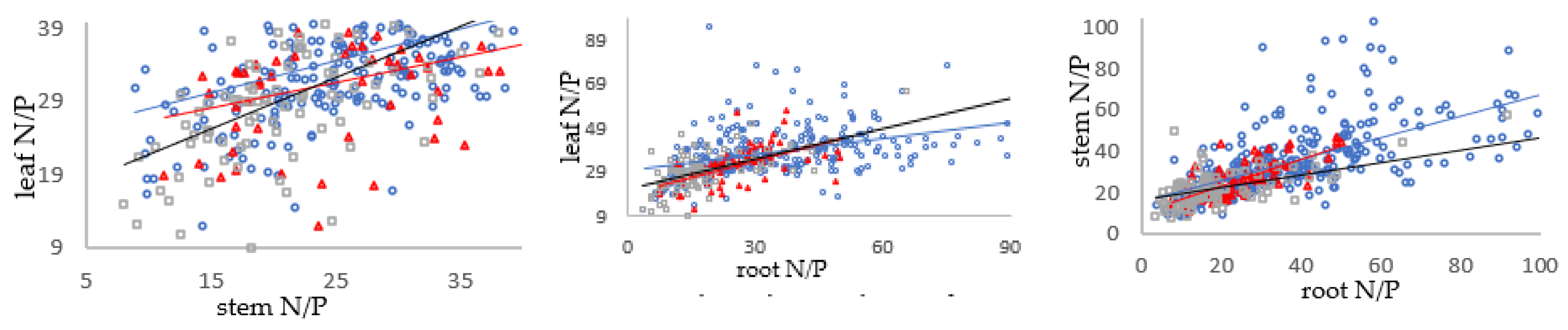
3.3. Relationships between C/N, N/P and C/P in Shrub Organs in Different Mycorrhizal Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, J.R.; Wang, G.L.; Meng, M.; Wang, R.C. Effects of drought stress on the stoichiometric characteristics in different organs of three shrub species. Chin. J. Appl. Ecol. 2021, 32, 73–81. [Google Scholar]
- Huang, D.; Wang, D.M.; Ren, Y. Using leaf nutrient stoichiometry as an indicator of flood tolerance and eutrophication in the riparian zone of the Lijang River. Ecol. Indic. 2019, 98, 821–829. [Google Scholar] [CrossRef]
- Wang, J.L.; Chen, G.F.; Zou, G.Y.; Song, X.F.; Liu, F.X. Comparative on plant stoichiometry response to agricultural non-point source pollution in different types of ecological ditches. Environ. Sci. Pollut. Res. 2019, 26, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Chen, L.Y.; Peng, Y.F.; Ding, J.Z.; Li, F.; Yang, G.B.; Kou, D.; Liu, L.; Fang, K.; Zhang, B.B.; et al. Linking microbial C: N: P stoichiometry to microbial community and abiotic factors along a 3500-km grassland transect on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2016, 25, 1416–1427. [Google Scholar] [CrossRef]
- Yang, W.; Zhou, J.G.; Wang, M.H. Spatial variation of ecological stoichiometry of soil C, N and P in a small hilly watershed in subtropics of China. Acta Petrol. 2015, 52, 1336–1344. [Google Scholar]
- Zeng, Q.C.; Li, X.; Dong, Y.H.; Li, Y.Y.; Chen, M.; An, S.S. Ecological stoichiometry characteristics and physical-chemical properties of soils at different latitudes on the Loess Plateau. J. Nat. Resour. 2015, 30, 870–879. [Google Scholar]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Yang, Y.H.; Luo, Y.Q. Carbon: Nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Ji, C.H.; Liu, H.L.; Cha, Z.Z.; Lin, Q.H.; Feng, G. Spatial-Temporal Variation of N, P, and K Stoichiometry in Cropland of Hainan Island. Agriculture 2021, 12, 39. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, B.R.; An, S.S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. Catena 2018, 166, 328–338. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y. Plant mixture balances terrestrial ecosystem C:N:P stoichiometry. Nat. Commun. 2021, 12, 4562. [Google Scholar] [CrossRef]
- Zeng, Q.C.; Li, X.; Dong, Y.H.; An, S.S.; Darboux, F. Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. Catena 2016, 147, 481–488. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Lv, S.Q.; Song, H.; Wang, M.C.; Zhao, Q.; Huang, H.; Niklas, K.J. Plant type dominates fine-root C: N: P stoichiometry across China: A meta-analysis. J. Biogeogr. 2020, 47, 1019–1029. [Google Scholar] [CrossRef]
- Li, Y.G.; Zhou, X.B.; Zhang, Y.M. Shrub modulates the stoichiometry of moss and soil in desert ecosystems, China. J. Arid Land 2019, 11, 579–594. [Google Scholar] [CrossRef] [Green Version]
- She, S.F.; Hu, Y.F.; Shu, X.Y.; Yan, X.; Li, Z.; Wang, Q.; He, J.; Jia, A.D. Variation of C, N and P stoichiometry in dominant understory plants during stand development in Salix cupularis plantations in alpine grassland in Northwestern Sichuan, China. Acta Pratacult. Sin. 2018, 27, 123–130. [Google Scholar]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Ågren, G.I. Stoichiometry and nutrition of plant growth in natural communities. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Wang, N.; Fu, F.Z.; Wang, B.T.; Wang, R.J. Carbon, nitrogen and phosphorus stoichiometry in Pinus tabulaeformis forest ecosystems in warm temperate Shanxi Province, north China. J. For. Res. 2018, 29, 1665–1673. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controlslitters decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Guo, L.L.; Chen, Y.; Zhang, Z.; Fukushima, T. N: P stoichiometry in a forested runoff during storm events: Comparisons with regions and vegetation types. Sci. World J. 2012, 2021, 257392. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, E103–E122. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.F.; Li, Q.; Lei, Z.F.; Zhang, J.B.; Song, X.Z.; Song, X.Z. Ecological stoichiometry of nitrogen and phosphorus in Moso bamboo (Phyllostachys edulis) during the explosive growth period of new emergent shoots. J. Plant. Res. 2019, 132, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Zhao, N.; Liu, C.C.; Yang, H.; Li, M.L.; Yu, G.R.; Wilcox, K.; Yu, Q.; He, N.P. C:N:P stoichiometry in China’s forests: From organs to ecosystems. Funct. Ecol. 2018, 32, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Tang, Z.Y.; Ji, C.J.; Liu, H.Y.; Ma, W.H.; Mohhamot, A.; Shi, Z.Y.; Sun, W.; Wang, T.; Wang, X.P.; et al. Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across Northern China. Sci. Rep. 2014, 4, 5448. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.F.; Freschet, G.T.; Pan, X.; Cornelissen, J.H.C.; Li, Y.; Dong, M. Coordinated variation in leaf and root traits across multiple spatial scales in Chinese semi-arid and arid ecosystems. New Phytol. 2010, 188, 543–553. [Google Scholar] [CrossRef]
- Chen, M.L.; Cui, J.T.; Deng, L.; Wang, K.B. Distribution patterns and ecological stoichiometry of water, carbon, nitrogen and phosphorus in different organs of two conifer species on the Loess Plateau. J. Environ. Earth 2018, 9, 54–63. [Google Scholar]
- Collins, C.G.; Wright, S.J.; Wurzburger, N. Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees. Oecologia 2016, 180, 1037–1047. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Xu, W.T.; Zhou, G.Y.; Bai, Y.F.; Li, J.X.; Tang, X.L.; Chen, D.M.; Liu, Q.; Ma, W.H.; Xiong, G.M.; et al. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef] [Green Version]
- Minden, V.; Kleyer, M. Internal and external regulation of plant organ stoichiometry. Plant Biol. 2014, 16, 897–907. [Google Scholar] [CrossRef]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, K.P.; Cornelissen, J.H.C.; Wang, Y.J.; Wu, C.B.; He, Y.J.; Ou, J.; Tan, Q.Y.; Xia, T.T.; Kang, L.L.; Guo, Y.; et al. AM Fungi Alleviate Phosphorus Limitation and Enhance Nutrient Competitiveness of Invasive Plants via Mycorrhizal Networks in Karst Areas. Front. Ecol. Evol. 2020, 8, 125. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Chagnon, P.L.; Bradley, R.L. Evidence that soil nutrient stoichiometry controls the competitive abilities of arbuscular mycorrhizal vs. root-borne non-mycorrhizal fungi. Fungal Ecol. 2013, 6, 557–560. [Google Scholar] [CrossRef]
- Han, L. Applied Research of Shrub in Urban Areas-Main Unban Area of Ankang as an Example. Shaanxi J. Agric. Sci. 2020, 66, 80–83. [Google Scholar]
- Averill, C.; Bhatnagar, J.M.; Dietze, M.C.; Pearse, W.D.; Kivlin, S.N. Global imprint of mycorrhizal fungi on whole-plant nutrient economics. Proc. Natl. Acad. Sci. USA 2019, 116, 23163–23168. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.Y.; Li, K.; Zhu, X.Y.; Wang, F.Y. The worldwide leaf economic spectrum traits are closely linked with mycorrhizal traits. Fungal Ecol. 2020, 43, 100877. [Google Scholar] [CrossRef]
- Jones, J.B. Laboratory Guide for Conducting Soil Tests and Plant Analysis; CRC Press: New York, NY, USA, 2001. [Google Scholar]
- Li, D.F.; Yu, S.L.; Wang, G.X.; Fang, W.W. Environmental heterogeneity and mechanism of stoichiometry properties of vegetative organs in dominant shrub communities across the Loess Plateau. Chin. J. Plant Ecol. 2015, 39, 453–465. [Google Scholar]
- Xiong, J.L.; Dong, L.W.; Lu, J.L.; Hu, W.G.; Gong, H.Y.; Xie, S.B.; Zhao, D.M.; Zhang, Y.H.; Wang, X.T.; Deng, Y.; et al. Variation in plant carbon, nitrogen and phosphorus contents across the drylands of China. Funct. Ecol. 2022, 36, 174–186. [Google Scholar] [CrossRef]
- Tao, Y.; Zhou, X.B.; Zhang, Y.M.; Yin, B.F.; Li, Y.G.; Zang, Y.X. Foliar C: N: P stoichiometric traits of herbaceous synusia and the spatial patterns and drivers in a temperate desert in Central Asia. Glob. Ecol. Conserv. 2021, 28, e01620. [Google Scholar] [CrossRef]
- Zhang, K.; He, M.Z.; Li, X.R.; Tan, H.J.; Gao, Y.H.; Li, G.; Han, G.J.; Wu, Y.Y. Foliar carbon, nitrogen and phosphorus stoichiometry of typical desert plants across the Alashan Desert. Acta Ecol. Sin. 2014, 34, 6538–6547. [Google Scholar]
- Zou, Z.G.; Zeng, F.P.; Zeng, Z.X.; Du, H.; Tang, H.; Zhang, H. The Variation in the Stoichiometric Characteristics of the Leaves and Roots of Karst Shrubs. Forests 2021, 12, 852. [Google Scholar] [CrossRef]
- Yang, X.; Chi, X.L.; Ji, C.J.; Liu, H.Y.; Ma, W.H.; Mohhammat, A.; Shi, Z.Y.; Wang, X.P.; Yu, S.L.; Yue, M.; et al. Variations of leaf N and P concentrations in shrubland biomes across northern China: Phylogeny, climate, and soil. Biogeosciences 2016, 13, 4429–4438. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.Y.; He, S.B.; Wang, Z.N.; Yang, H.M. Carbon, nitrogen and phosphorus stoichiometry in differently aged lucerne stands during flowering. Legum. Res. 2016, 39, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, W.C.; Xu, M.P.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Response of forest growth to C: N: P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Liu, X.Y.; Hu, Y.K. C:N:P stoichiometry of leaves and fine roots in typical forest swamps of the GreaterHinggan Mountains, China. Yingyong Shengtai Xuebao 2020, 31, 3385–3394. [Google Scholar]
- Gao, Y.; Yuan, Y.; Li, Q.; Kou, L.; Fu, X.L.; Dai, X.Q.; Wang, H.M. Mycorrhizal type governs foliar and root multi-elemental stoichiometries of trees mainly via root traits. Plant Soil 2021, 460, 229–246. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Lü, X.T.; Hartmann, H.; Keller, A.; Han, X.G.; Trumbore, S.; Phillips, R.P. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales. Glob. Ecol. Biogeogr. 2018, 27, 875–885. [Google Scholar] [CrossRef]
- Vrede, T.; Dobberfuhl, D.R.; Kooijman, S.A.L.M.; Elser, J.J. Fundamental connections among organism C: N: P stoichiometry, macromolecular composition, and growth. Ecology 2004, 85, 1217–1229. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon-nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef]
- Dong, C.G.; Qiao, Y.N.; Cao, Y.; Chen, Y.M.; Wu, X.; Xue, W.Y. Seasonal variations in carbon, nitrogen, and phosphorus stoichiometry of a Robinia pseudoacacia plantation on the Loess Hilly region. China. Forests 2021, 12, 214. [Google Scholar] [CrossRef]
- Bennett, J.A.; Maherali, H.; Reinhart, K.O.; Lekberg, Y.; Hart, M.M.; Klironomos, J. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 2017, 355, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Kohler, A.; Kuo, A.; Nagy, L.G.; Morin, E.; Barry, K.W.; Buscot, F.; Canback, B.; Choi, C.; Cichocki, N.; Clum, A.; et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 2015, 47, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Law, B.E. Regional analysis of drought and heat impacts on forests: Current and future science directions. Glob. Chang. Biol. 2014, 20, 3595–3599. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.P.; Yan, Z.B.; Gheyret, G.; Zhou, G.Y.; Xie, Z.Q.; Tang, Z.Y. The community-level scaling relationship between leaf nitrogen and phosphorus changes with plant growth, climate and nutrient limitation. J. Ecol. 2020, 108, 1276–1286. [Google Scholar] [CrossRef]
- Zhang, J.H.; Li, M.X.; Xu, L.; Zhu, J.X.; Dai, G.H.; He, N.P. C: N: P stoichiometry in terrestrial ecosystems in China. Sci. Total Environ. 2021, 795, 148849. [Google Scholar] [CrossRef]
- Ågren, G.I. The C: N: P stoichiometry of autotrophs–theory and observations. Ecol. Lett. 2004, 7, 185–191. [Google Scholar] [CrossRef]
- Li, H.; Crabbe, M.J.C.; Xu, F.; Wang, W.; Niu, R.; Gao, X.; Zhang, P.; Chen, H.K. Seasonal variations in carbon, nitrogen and phosphorus concentrations and C: N: P stoichiometry in the leaves of differently aged Larix principis-rupprechtii Mayr. plantations. Forests 2017, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Shi, Z.Y.; Zhang, M.H.; Li, Y.; Gao, J.K.; Wang, X.G. Stoichiometry of Carbon, Nitrogen and Phosphorus in Shrub Organs Linked Closely with Mycorrhizal Strategy in Northern China. Front. Plant Sci. 2021, 12, 687347. [Google Scholar] [CrossRef]
- Ma, X.X.; Hong, J.T.; Wang, X.D. C: N: P stoichiometry of perennial herbs’ organs in the alpine steppe of the northern Tibetan Plateau. J. Mt. Sci. 2019, 16, 2039–2047. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H.; Reich, P.B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Nat. Commun. 2011, 2, 2555–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brundrett, M.C. Global diversity and importance of mycorrhizal and non-mycorrhizal plants. Ecol. Stud. 2017, 230, 533–556. [Google Scholar]
- Sardans, J.; Peñuelas, J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol. 2012, 160, 1741–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Z.; Li, D.D.; Jiao, F.; Yao, J.; Du, H.T. The latitudinal patterns of leaf and soil C: N: P stoichiometry in the loess plateau of China. Front. Plant Sci. 2019, 10, 85. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio | Mycorrhiza Type | Latitude | Mycorrhizatype × Latitude | |||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| leaf C/N | 0.28 | 0.75 | 2.33 | 0.13 | 4.70 | 0.01 |
| leaf N/P | 3.35 | 0.04 | 2.12 | 0.15 | 7.62 | <0.001 |
| leaf C/P | 3.24 | 0.04 | 0.12 | 0.73 | 4.54 | 0.01 |
| stem C/N | 1.80 | 0.17 | 49.25 | <0.001 | 10.80 | <0.001 |
| stem N/P | 4.72 | 0.01 | 12.65 | <0.001 | 4.73 | 0.009 |
| stem C/P | 3.35 | 0.04 | 6.96 | 0.009 | 0.66 | 0.52 |
| root C/N | 2.58 | 0.08 | 14.40 | <0.001 | 3.73 | 0.02 |
| root N/P | 3.49 | 0.04 | 9.18 | 0.003 | 3.97 | 0.02 |
| root C/P | 1.38 | 0.26 | 0.11 | 0.74 | 0.45 | 0.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Shi, Z.; Sun, Y.; Wang, X.; Yang, W.; Gao, J.; Wang, X. Stoichiometric Ratios of Carbon, Nitrogen and Phosphorus of Shrub Organs Vary with Mycorrhizal Type. Agriculture 2022, 12, 1061. https://doi.org/10.3390/agriculture12071061
Yang S, Shi Z, Sun Y, Wang X, Yang W, Gao J, Wang X. Stoichiometric Ratios of Carbon, Nitrogen and Phosphorus of Shrub Organs Vary with Mycorrhizal Type. Agriculture. 2022; 12(7):1061. https://doi.org/10.3390/agriculture12071061
Chicago/Turabian StyleYang, Shuang, Zhaoyong Shi, Yichun Sun, Xiaohui Wang, Wenya Yang, Jiakai Gao, and Xugang Wang. 2022. "Stoichiometric Ratios of Carbon, Nitrogen and Phosphorus of Shrub Organs Vary with Mycorrhizal Type" Agriculture 12, no. 7: 1061. https://doi.org/10.3390/agriculture12071061