
Effect Mechanism of Solar Radiation on Maize Yield Formation
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling and Measurement
2.3. Statistical Analysis
3. Results
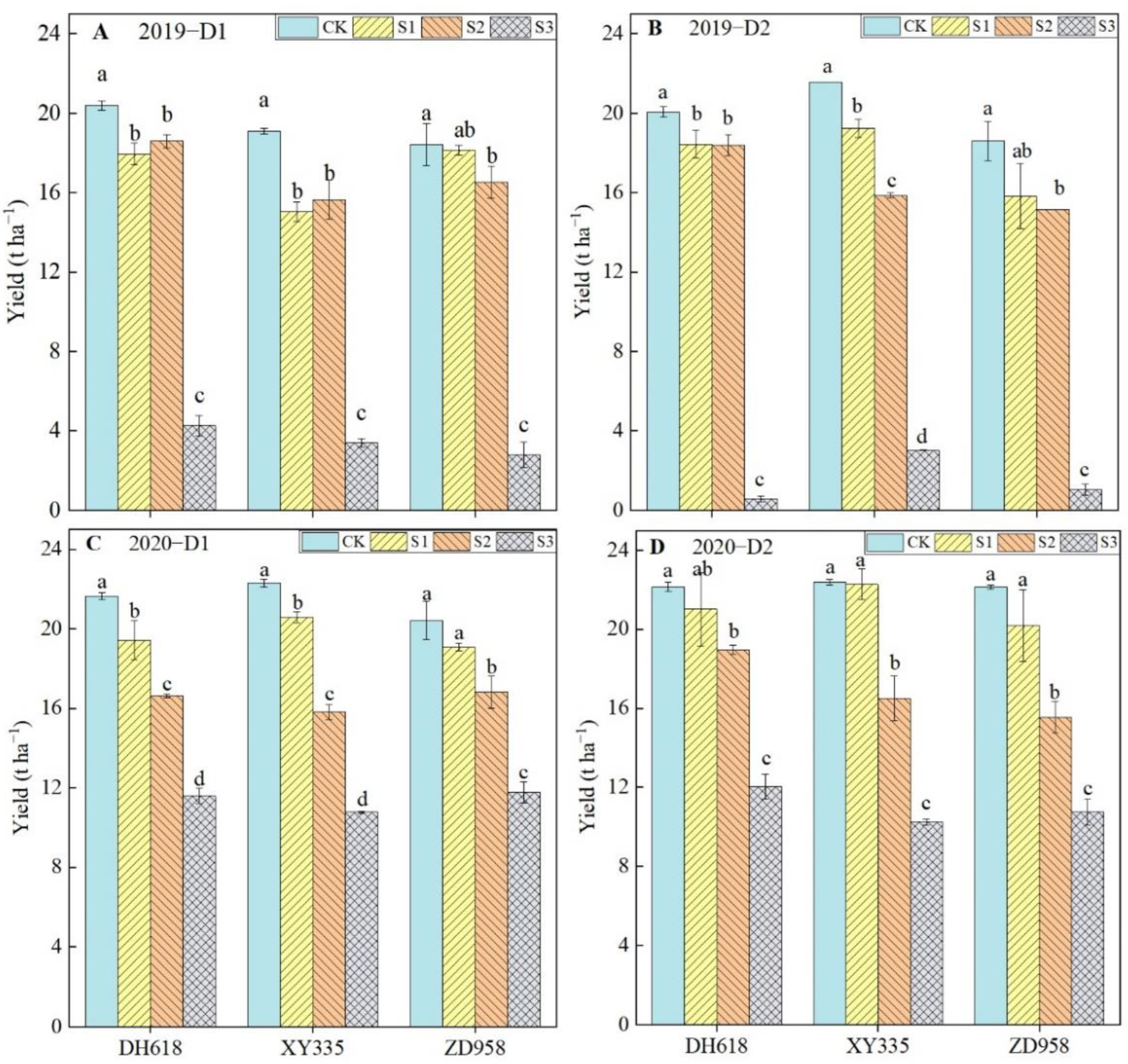
3.1. Effects of Shading on Maize Yields under Different Planting Densities
3.2. Effects of Shading on Maize Ear Number and Barrenness
3.3. Effects of Shading on Maize Ear Traits and the Relationships between PAR and Both the Number of Kernels Per Row and the Grain Abortion Rate
3.4. Quantitative Relationships between Grain Weights and Accumulated Solar Radiation
3.5. Relationships between Ear Characteristics and Maize Yields
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luan, X.Y.; Bommarco, R.; Scaini, A.; Vico, G. Combined heat and drought suppress rainfed maize and soybean yields and modify irrigation benefits in the USA. Environ. Res. Lett. 2021, 16, 064023. [Google Scholar] [CrossRef]
- Chen, X.C.; Chen, F.J.; Chen, Y.L.; Gao, Q.; Yang, X.L.; Yuan, L.X.; Zhang, F.S.; Mi, G.H. Modern maize hybrids in Northeast China exhibit increased yield potential and resource use efficiency despite adverse climate change. Glob. Chang. Biol. 2013, 19, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Liu, Y.E.; Liu, W.M.; Yang, H.; Xie, R.Z.; Wang, K.R.; Ming, B.; Liu, G.Z.; Xue, J.; Wang, Y.H.; et al. Quantifying maize grain yield losses caused by climate change based on extensive field data across China. Resour., Conserv. Recycl. 2021, 174, 105811. [Google Scholar] [CrossRef]
- Zhang, Q.; Zheng, Y.; Tong, D.; Shao, M.; Wang, S.; Zhang, Y.; Xu, X.; Wang, J.; He, H.; Liu, W.; et al. Drivers of improved PM2.5 air quality in China from 2013 to 2017. Proc. Natl. Acad. Sci. USA 2019, 116, 24463–24469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.F.; Liu, B.H.; Yang, H.S.; Chen, X.P. Solar dimming decreased maize yield potential on the North China Plain. Food Energy Secur. 2020, 9, e235. [Google Scholar] [CrossRef]
- Mu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Long-term low radiation decreases leaf photosynthesis, photochemical efficiency and grain yield in winter wheat. J. Agron. Crop Sci. 2010, 196, 38–47. [Google Scholar] [CrossRef]
- Li, X.Y.; Wagner, F.; Peng, W.; Yang, J.N.; Mauzerall, D.L. Reduction of solar photovoltaic resources due to air pollution in China. Proc. Natl. Acad. Sci. USA 2017, 114, 11867–11872. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Z.; Liu, W.M.; Hou, P.; Ming, B.; Yang, Y.S.; Guo, X.X.; Xie, R.Z.; Wang, K.R.; Li, S.K. Reducing maize yield gap by matching plant density and solar radiation. J. Integr. Agric. 2021, 20, 363–370. [Google Scholar] [CrossRef]
- Liu, W.M.; Hou, P.; Liu, G.Z.; Yang, Y.S.; Guo, X.X.; Ming, B.; Xie, R.; Wang, K.R.; Liu, Y.E.; Li, S.K. Contribution of total dry matter and harvest index to maize grain yield-A multisource data analysis. Food Energy Secur. 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Liu, Y.E.; Hou, P.; Huang, G.R.; Zhong, X.L.; Li, H.R.; Zhao, J.R.; Li, S.K.; Mei, X.R. Maize grain yield and water use efficiency in relation to climatic factors and plant population in northern China. J. Integr. Agric. 2021, 20, 3156–3169. [Google Scholar] [CrossRef]
- Jia, S.F.; Li, C.F.; Dong, S.T.; Zhang, J.W. Effects of shading at different stages after anthesis on maize grain weight and quality at cytology level. Agric. Sci. Chin. 2011, 10, 58–69. [Google Scholar] [CrossRef]
- Zhao, J.; Kong, X.; He, K.; Xu, H.; Mu, J. Assessment of the radiation effect of aerosols on maize production in China. Sci. Total Environ. 2020, 720, 137567. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhao, B.; Dong, S.T.; Liu, P.; Ren, B.C.; Zhang, J.W. Response of summer maize photosynthate accumulation and distribution to shading stress assessed by using 13CO2 stable isotope tracer in the field. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, P.; Liu, Y.E.; Liu, W.M.; Liu, G.Z.; Xie, R.Z.; Wang, K.R.; Ming, B.; Wang, Y.H.; Zhao, R.L.; Zhang, W.J.; et al. How to increase maize production without extra nitrogen input. Resour., Conserv. Recycl. 2020, 160, 104913. [Google Scholar] [CrossRef]
- Ranum, P.; Peña-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef]
- Liu, G.Z.; Hou, P.; Xie, R.Z.; Ming, B.; Wang, K.R.; Xu, W.J.; Liu, W.M.; Yang, Y.S.; Li, S.K. Canopy characteristics of high-yield maize with yield potential of 22.5 Mg ha−1. Field Crops Res. 2017, 213, 221–230. [Google Scholar] [CrossRef]
- Gao, J.; Shi, J.; Dong, S.T.; Liu, P.; Zhao, B.; Zhang, J.W. Grain yield and root characteristics of summer maize (Zea mays L.) under shade stress conditions. J. Agron. Crop Sci. 2017, 203, 562–573. [Google Scholar] [CrossRef]
- Naseer, M.A.; Hussain, S.; Nengyan, Z.; Ejaz, I.; Ahmad, S.; Farooq, M.; Xiaolong, R. Shading under drought stress during grain filling attenuates photosynthesis, grain yield and quality of winter wheat in the Loess Plateau of China. J. Agron. Crop Sci. 2021, 208, 255–263. [Google Scholar] [CrossRef]
- Yang, Y.S.; Xu, W.J.; Hou, P.; Liu, G.Z.; Liu, W.M.; Wang, Y.H.; Zhao, R.L.; Ming, B.; Xie, R.Z.; Wang, K.R.; et al. Improving maize grain yield by matching maize growth and solar radiation. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.Y.; Camberato, J.J.; Jin, L.B.; Zhang, J.W. Effects of shading on spike differentiation and grain yield formation of summer maize in the field. Int. J. Biometeorol. 2015, 59, 1189–1200. [Google Scholar] [CrossRef]
- Andayani, N.N.; Riadi, M.; Rafiuddin; Kalqutny, S.H.; Efendi, R.; Azrai, M. Evaluation of yield and agronomic components of three-way cross maize hybrids under low-light environment. Earth Environ. Sci. 2020, 484, 012016. [Google Scholar] [CrossRef]
- Li, C.H.; Luan, L.M.; Yin, F.; Wang, Q.; Zhao, Y.L. Effects of light stress at different stages on the growth and yield of different maize genotypes (Zea mays L.). Acta Ecol. Sin. 2005, 25, 824–830. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, X.X.; Liu, G.Z.; Liu, W.M.; Xue, J.; Ming, B.; Xie, R.Z.; Wang, K.R.; Hou, P.; Li, S.K. Solar radiation effects on dry matter accumulations and transfer in maize. Front. Plant Sci. 2021, 12, 727134. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.L.; Sun, X.L.; Wang, X.; Yan, F.B.; Lu, W.P. Effect of shading during grain filling on the physicochemical properties of fresh waxy maize. J. Integr. Agric. 2013, 12, 1560–1567. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Shi, Y.L.; Xu, R.C.; Lu, D.L.; Lu, W.P. Effects of shading after pollination on kernel filling and physicochemical quality traits of waxy maize. Crop J. 2016, 4, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.Z.; Liu, J.Y.; Cai, Z.Y.; Wang, H.Q.; Fu, J.F.; Zhang, H.T.; Zhang, Y.D.; Zhu, S.D.; Wu, W.H.; Yan, H.X.; et al. Shade stress on maize seedlings biomass production and photosynthetic traits. Cienc. Rural 2022, 52, e20201022. [Google Scholar] [CrossRef]
- Gao, J.; Shi, J.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Effect of different light intensities on root characteristics and grain yield of summer maize (Zea mays L.). Sci. Agric. Sin. 2017, 50, 2104–2113. [Google Scholar]
- Shi, K.; Gu, X.T.; Lu, W.P.; Lu, D.L. Effects of weak-light stress during grain filling on the physicochemical properties of normal maize starch. Carbohydr. Polym. 2018, 202, 47–55. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, M.; Dong, S.T.; Chen, C.Y.; Sun, R. “Three combination structure” quantitative expression and high yield analysis in crops. Acta Agron. Sin. 2007, 33, 1674–1681. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, X.; Lin, X.; Sassenrath, G.F.; Dai, S.; Lv, S.; Chen, X.; Chen, F.; Mi, G. Radiation interception and use efficiency contributes to higher yields of newer maize hybrids in northeast China. Agron. J. 2015, 107, 1473–1480. [Google Scholar] [CrossRef]
- Earley, E.B.; Miller, R.J.; Reichert, G.L. Effect of shade on maize production under field conditions. Crop Sci. 1966, 6, 1–7. [Google Scholar] [CrossRef]
- Zhou, W.X.; Dong, P.F.; Wang, X.P.; Li, C.H. Effects of low-light stress on kernel setting and metabolism of carbon and nitrogen in different maize (Zea mays L.) genotypes. Acta Agron. Sin. 2013, 39, 1826–1834. [Google Scholar] [CrossRef]
- Zhang, J.W.; Dong, S.T.; Wang, K.J.; Hu, C.H.; Liu, P. Effects of shading on the growth, development and grain yield of summer maize. Chin. J. Appl. Ecol. 2006, 17, 657–662. [Google Scholar]
- Zhong, X.M.; Shi, Z.S. Reserach progress on corn shading stress. J. Maize Sci. 2012, 20, 138–141. [Google Scholar] [CrossRef]
- Zhong, X.M.; Shi, Z.S.; Li, F.H.; Huang, H.J.; Li, H.Y. Effects of light intensity on ear differentiation of barren-stalk defective maize. Adv. Mater. Res. 2013, 726–731, 4864–4869. [Google Scholar] [CrossRef]
- Mbewe, D.M.N.; Hunter, R.B. The effect of shade stress on the performance of corn for silage versus grain. Can. J. Plant Sci. 1986, 66, 53–60. [Google Scholar] [CrossRef]
- Zhao, J.R.; Chen, G.P. Effects of shading treatment at different stages of plant development on grain production of corn (Zea Mays L.) and observations of tip kernel abortion. Sci. Agric. Sin. 1990, 23, 28–34. [Google Scholar] [CrossRef]
- Zhang, J.W.; Wu, H.X.; Dong, S.T.; Wang, K.J.; Hu, C.H.; Liu, P. Effects of shading on yield and quality of summer maize. J. Maize Sci. 2009, 17, 124–129. [Google Scholar] [CrossRef]
- Chen, C.Y.; Wang, R.H.; Zhao, J.R.; Xu, T.J.; Wang, Y.D.; Liu, X.Z.; Liu, C.G.; Pei, Z.C.; Cheng, G.L.; Chen, G.P. Effects of shading on grain-filling properties and yield of maize at different growth stages. Acta Agron. Sin. 2014, 40, 1650–1657. [Google Scholar] [CrossRef]
- Meng, J.J.; Dong, S.T.; Shi, D.Y.; Zhang, H.Y. Relationship of ear differentiation with kernel development and barrenness in maize (Zea mays L.). Acta Agron. Sin. 2013, 39, 912–918. [Google Scholar] [CrossRef]
- He, H.Y.; Hu, Q.; Li, R.; Pan, X.B.; Huang, B.X.; He, Q.J. Regional gap in maize production, climate and resource utilization in China. Field Crops Res. 2020, 254, 107830. [Google Scholar] [CrossRef]
- Xu, K.; Zhu, X.F.; Liu, Y.; Guo, R.; Cheng, L.Y. Effects of drought on maize yield under climate change in China. Trans. Chin. Soc. Agric. Eng. 2020, 36, 149–158. [Google Scholar] [CrossRef]
- Xu, W.J.; Liu, C.W.; Wang, K.R.; Xie, R.Z.; Ming, B.; Wang, Y.H.; Zhang, G.Q.; Liu, G.Z.; Zhao, R.L.; Fan, P.P.; et al. Adjusting maize plant density to different climatic conditions across a large longitudinal distance in China. Field Crops Res. 2017, 212, 126–134. [Google Scholar] [CrossRef]
- Liu, G.Z.; Hou, P.; Xie, R.Z.; Ming, B.; Wang, K.R.; Liu, W.M.; Yang, Y.S.; Xu, W.J.; Chen, J.L.; Li, S.K. Nitrogen uptake and response to radiation distribution in the canopy of high-yield maize. Crop Sci. 2019, 59, 1236–1247. [Google Scholar] [CrossRef]
- Ma, D.L.; Li, S.K.; Zhai, L.C.; Yu, X.F.; Xie, R.Z.; Gao, J.L. Response of maize barrenness to density and nitrogen increases in Chinese cultivars released from the 1950s to 2010s. Field Crops Res. 2020, 250, 107766. [Google Scholar] [CrossRef]
- Wang, X.H.; Zhang, L.; Liu, S.L.; Jun, C.Y.; Wei, W.W.; Liu, C.G.; Wang, Y.J.; Bian, S.F.; Wang, L.C. Grain filling characteristics of maize hybrids differing in maturities. Sci. Agric. Sin. 2014, 47, 3557–3565. [Google Scholar] [CrossRef]
- Guo, X.X.; Yang, Y.S.; Liu, H.F.; Liu, G.Z.; Liu, W.M.; Wang, Y.H.; Zhao, R.L.; Ming, B.; Xie, R.Z.; Wang, K.R.; et al. Effects of solar radiation on root and shoot growth of maize and the quantitative relationship between them. Crop Sci. 2020, 61, 1414–1425. [Google Scholar] [CrossRef]
- Zhai, L.; Wang, Z.B.; Song, S.J.; Zhang, L.H.; Zhang, Z.B.; Jia, X.L. Tillage practices affects the grain filling of inferior kernel of summer maize by regulating soil water content and photosynthetic capacity. Agric. Water Manag. 2021, 245, 106600. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, X.N.; Shao, J.F.; Liu, Z.Y.; Li, Z.X. Effects of light intensity at full growing stage on the growth and yield of different maize varieties. J. Jilin Agric. Univ. 2008, 30, 769–773. [Google Scholar] [CrossRef]
- Estrada-Campuzano, G.; Miralles, D.J.; Slafer, G.A. Yield determination in triticale as affected by radiation in different development phases. Eur. J. Agron. 2008, 28, 597–605. [Google Scholar] [CrossRef]
- Peltonen-Sainio, P.; Kangas, A.; Salo, Y.; Jauhiainen, L. Grain number dominates grain weight in temperate cereal yield determination: Evidence based on 30 years of multi-location trials. Field Crops Res. 2007, 100, 179–188. [Google Scholar] [CrossRef]
- Sadras, V.O. Evolutionary aspects of the trade-off between seed size and number in crops. Field Crops Res. 2007, 100, 125–138. [Google Scholar] [CrossRef]
- Fischer, R.A. The importance of grain or kernel number in wheat: A reply to Sinclair and Jamieson. Field Crops Res. 2008, 105, 15–21. [Google Scholar] [CrossRef]
- Hou, P.; Cui, Z.L.; Bu, L.D.; Yang, H.S.; Zhang, F.S.; Li, S.K. Evaluation of a modified Hybrid-Maize model incorporating a newly developed module of plastic film mulching. Crop Sci. 2014, 54, 2796–2804. [Google Scholar] [CrossRef]
- Yan, G.Z.; Li, W.Y. Research on application of information technology and mathematical modeling in precision agriculture. In Proceedings of the International Conference on Industrial and Information Systems, IEEE, Haikou, China, 24–25 April 2009; pp. 228–231. [Google Scholar]
- Wang, Z.X.; Gao, X.S.; Xu, J.F.; Liu, Y.J.; Luo, Y.N. A study on the grain abortion of maize (Zea mays L.). Sci. Agric. Sin. 1986, 6, 36–40. [Google Scholar]
- Li, X.L.; Zhao, M.; Li, C.F.; Ge, J.Z.; Hou, H.P.; Li, Q.; Hou, L.B. Effect of sowing-date and planting density on dry matter accumulation dynamic and establishment of its simulated model in maize. Acta Agron. Sin. 2011, 36, 2143–2153. [Google Scholar] [CrossRef]
- Pandey, A.; Dou, F.; Morgan, C.L.S.; Guo, J.; Deng, J.; Schwab, P. Modeling organically fertilized flooded rice systems and its long-term effects on grain yield and methane emissions. Sci. Total Environ. 2021, 755, 142578. [Google Scholar] [CrossRef]
- Liu, G.Z.; Yang, H.S.; Xie, R.Z.; Yang, Y.S.; Liu, W.M.; Guo, X.X.; Xue, J.; Ming, B.; Wang, K.K.; Hou, P.; et al. Genetic gains in maize yield and related traits for high-yielding cultivars released during 1980s to 2010s in China. Field Crops Res. 2021, 270, 108223. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | |||||
|---|---|---|---|---|---|---|
| Treatment | Ear Density (104 ha−1) | Decrease Rate (%) | Earless Plant Rate (%) | Ear Density (104 ha−1) | Decrease Rate (%) | Earless Plant Rate (%) |
| DH−D1−CK | 7.7 a | 0.0 b | 7.8 a | 0.8 a | ||
| DH−D1−S1 | 7.6 a | 0.8 | 1.6 b | 6.8 b | 12.4 | 0.0 a |
| DH−D1−S2 | 7.4 a | 3.9 | 1.6 b | 7.5 ab | 4.7 | 0.7 a |
| DH−D1−S3 | 4.7 b | 38.6 | 29.2 a | 6.7 c | 14.0 | 3.2 a |
| DH−D2−CK | 11.2 a | 0.0 b | 11.7 a | 4.3 b | ||
| DH−D2−S1 | 10.9 a | 2.7 | 3.4 b | 11.0 a | 6.5 | 2.0 ab |
| DH−D2−S2 | 11.0 a | 2.2 | 8.1 b | 11.3 a | 3.9 | 1.0 ab |
| DH−D2−S3 | 2.0 b | 82.0 | 73.6 a | 10.2 b | 12.7 | 6.1 a |
| XY−D1−CK | 7.6 a | 0.0 b | 7.7 a | 0.8 a | ||
| XY−D1−S1 | 7.4 a | 3.6 | 2.4 b | 7.5 a | 2.4 | 1.6 a |
| XY−D1−S2 | 7.1 a | 7.1 | 4.3 b | 7.5 a | 3.1 | 0.8 a |
| XY−D1−S3 | 4.4 b | 42.1 | 17.2 a | 7.0 b | 8.7 | 2.3 a |
| XY−D2−CK | 10.0 a | 0.0 b | 11.5 a | 0.4 b | ||
| XY−D2−S1 | 10.0 a | 0.0 | 0.0 b | 11.6 a | −0.8 | 1.0 b |
| XY−D2−S2 | 10.0 a | 0.0 | 0.0 b | 10.9 b | 5.5 | 4.4 b |
| XY−D2−S3 | 3.4 b | 66.1 | 46.7 a | 10.1 c | 12.9 | 12.2 a |
| ZD−D1−CK | 8.8 a | 0.0 b | 8.1 a | 0.8 a | ||
| ZD−D1−S1 | 8.5 a | 2.8 | 0.0 b | 7.9 a | 3.0 | 0.8 a |
| ZD−D1−S2 | 8.0 a | 9.0 | 0.8 b | 7.8 a | 3.7 | 0.8 a |
| ZD−D1−S3 | 4.5 b | 48.3 | 14.1 a | 7.4 b | 9.0 | 0.0 a |
| ZD−D2−CK | 11.3 a | 0.0 b | 12.3 a | 0.5 b | ||
| ZD−D2−S1 | 10.6 a | 5.6 | 1.1 b | 12.1 ab | 1.5 | 0.0 b |
| ZD−D2−S2 | 11.1 a | 1.6 | 2.8 b | 11.8 ab | 3.7 | 1.5 ab |
| ZD−D2−S3 | 2.8 b | 82.0 | 22.1 a | 11.5 b | 5.9 | 4.3 a |
| 2019 | 2020 | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | No. of Rows Per Ear | No. of Kernels Per Row | Bald Tip Length (cm) | Kernel Abortion Rate (%) | No. of Rows Per Ear | No. of Kernels Per Row | Bald Tip Length (cm) | Kernel Abortion Rate (%) |
| DH−D1−CK | 16.0 a | 35.8 a | 0.6 c | 9.2 b | 15.7 a | 38.5 a | 0.7 c | 29.1 c |
| DH−D1−S1 | 16.0 a | 35.4 a | 2.0 bc | 5.4 b | 15.7 a | 37.2 ab | 1.4 bc | 34.4 b |
| DH−D1−S2 | 15.0 a | 35.0 a | 1.5 b | 9.9 b | 15.3 a | 34.5 b | 2.0 b | 44.4 a |
| DH−D1−S3 | 15.2 a | 16.8 b | 3.6 a | 52.3 a | 15.7 a | 27.8 c | 5.6 a | 46.8 a |
| DH−D2−CK | 15.0 ab | 31.2 a | 2.1 c | 27.2 c | 15.3 a | 33.7 a | 2.9 b | 39.1 b |
| DH−D2−S1 | 15.5 a | 25.5 b | 3.0 ab | 25.9 c | 16.0 a | 31.8 a | 3.6 b | 40.8 b |
| DH−D2−S2 | 14.4 ab | 26.0 b | 3.1 a | 33.2 b | 16.3 a | 24.8 b | 4.3 ab | 54.1 a |
| DH−D2−S3 | 12.6 b | 14.4 c | 2.4 bc | 60.6 a | 16.4 a | 18.2 c | 5.9 a | 57.5 a |
| XY−D1−CK | 16.0 a | 38.3 a | 1.4 b | 19.2 c | 16.0 a | 40.3 a | 2.0 b | 26.4 b |
| XY−D1−S1 | 16.8 a | 39.0 a | 1.1 b | 22.2 c | 17.7 a | 35.5 b | 2.3 b | 32.9 ab |
| XY−D1−S2 | 16.8 a | 34.0 b | 1.7 ab | 32.0 b | 17.7 a | 29.0 c | 4.2 a | 37.2 a |
| XY−D1−S3 | 16.4 a | 18.4 c | 2.7 a | 60.2 a | 16.8 a | 25.2 d | 5.0 a | 38.8 a |
| XY−D2−CK | 16.0 a | 36.6 a | 2.2 b | 19.5 c | 17.3 a | 33.6 a | 4.4 bc | 44.1 b |
| XY−D2−S1 | 16.0 a | 32.4 b | 2.7 ab | 41.4 b | 15 a | 30.0 a | 4.3 c | 44.3 b |
| XY−D2−S2 | 15.2 ab | 29.4 b | 2.4 ab | 43.5 b | 16 a | 23.8 b | 5.2 ab | 56.0 a |
| XY−D2−S3 | 14.8 b | 17.2 c | 3.5 a | 79.9 a | 17 a | 16.5 c | 5.5 a | 58.8 a |
| ZD−D1−CK | 17.2 a | 34.0 a | 0.9 a | 12.7 a | 15.0 a | 37.2 a | 1.8 b | 30.7 b |
| ZD−D1−S1 | 15.6 ab | 34.8 a | 1.3 a | 14.5 a | 16.3 a | 38.0 a | 2.0 b | 32.8 b |
| ZD−D1−S2 | 17.2 a | 32.8 a | 0.8 a | 19.5 a | 15.7 a | 31.7 a | 2.8 b | 43.0 a |
| ZD−D1−S3 | 15.0 b | 26.0 b | 2.0 a | 43.9 a | 15.3 a | 24.8 b | 4.1 a | 44.0 a |
| ZD−D2−CK | 16.0 a | 33.8 a | 1.8 a | 26.3 b | 16.0 a | 31.8 a | 2.4 b | 21.6 b |
| ZD−D2−S1 | 14.8 a | 30.0 a | 1.8 a | 41.1 a | 15.3 a | 29.3 ab | 2.3 b | 33.1 a |
| ZD−D2−S2 | 14.8 a | 29.8 a | 1.2 a | 42.6 a | 15.0 a | 26.8 b | 3.3 ab | 39.5 a |
| ZD−D2−S3 | 15.0 a | 16.7 b | 1.6 a | 72.0 a | 15.7 a | 26.7 b | 4.1 a | 36.0 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Liu, G.; Guo, X.; Liu, W.; Xue, J.; Ming, B.; Xie, R.; Wang, K.; Li, S.; Hou, P. Effect Mechanism of Solar Radiation on Maize Yield Formation. Agriculture 2022, 12, 2170. https://doi.org/10.3390/agriculture12122170
Yang Y, Liu G, Guo X, Liu W, Xue J, Ming B, Xie R, Wang K, Li S, Hou P. Effect Mechanism of Solar Radiation on Maize Yield Formation. Agriculture. 2022; 12(12):2170. https://doi.org/10.3390/agriculture12122170
Chicago/Turabian StyleYang, Yunshan, Guangzhou Liu, Xiaoxia Guo, Wanmao Liu, Jun Xue, Bo Ming, Ruizhi Xie, Keru Wang, Shaokun Li, and Peng Hou. 2022. "Effect Mechanism of Solar Radiation on Maize Yield Formation" Agriculture 12, no. 12: 2170. https://doi.org/10.3390/agriculture12122170