Phenology and Dwarfing Gene Interaction Effects on the Adaptation of Selected Wheat (Triticum aestivum L.) Advanced Lines across Diverse Water-Limited Environments of Western Australia

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Experiments
2.3. Agronomic Traits
2.4. Genotyping of the Plant Materials
2.5. Statistical Analysis
3. Results
3.1. Allelic Distribution at The Vrn1 and Ppd loci
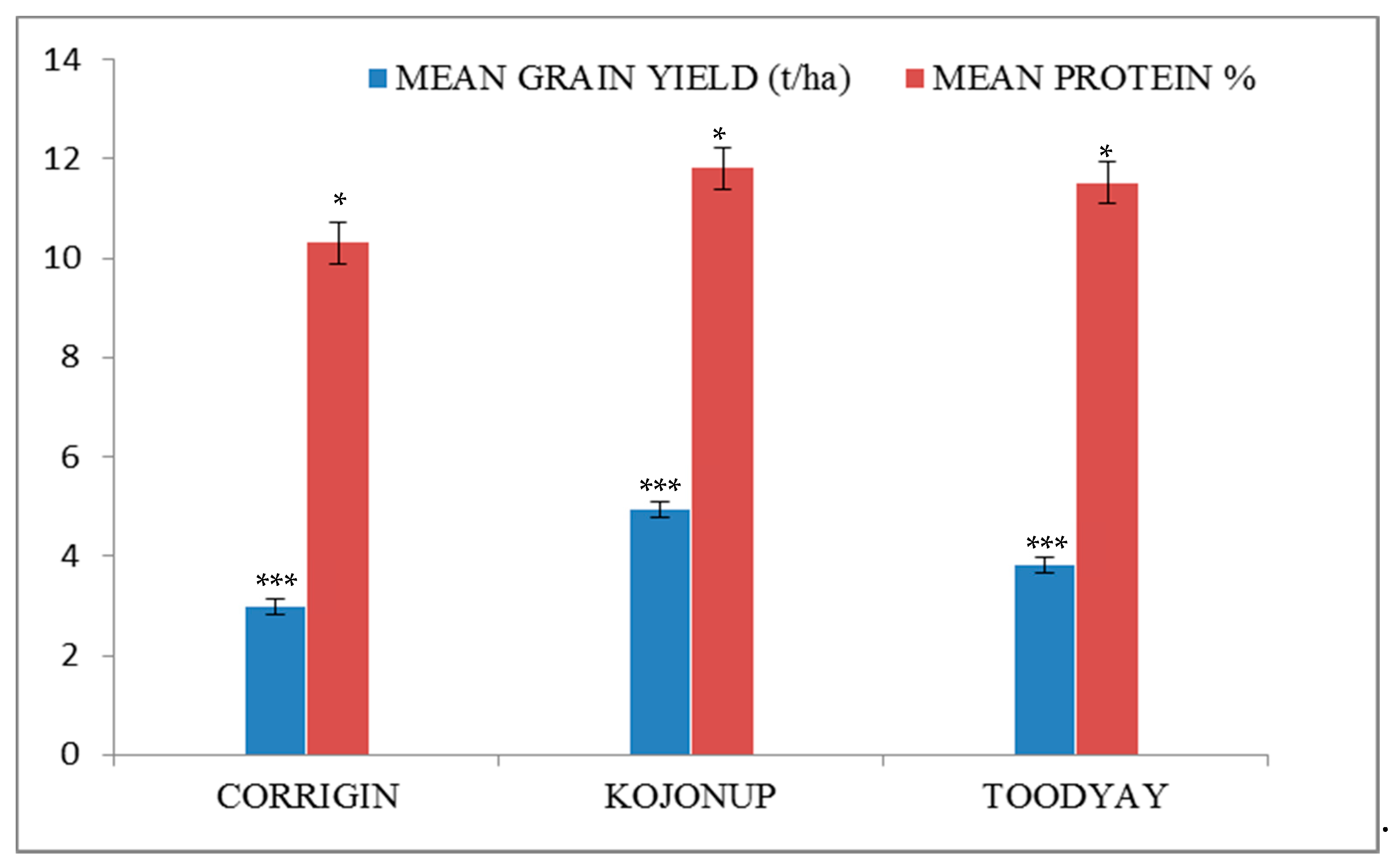
3.2. Environmental Effects on Yield and Protein Content
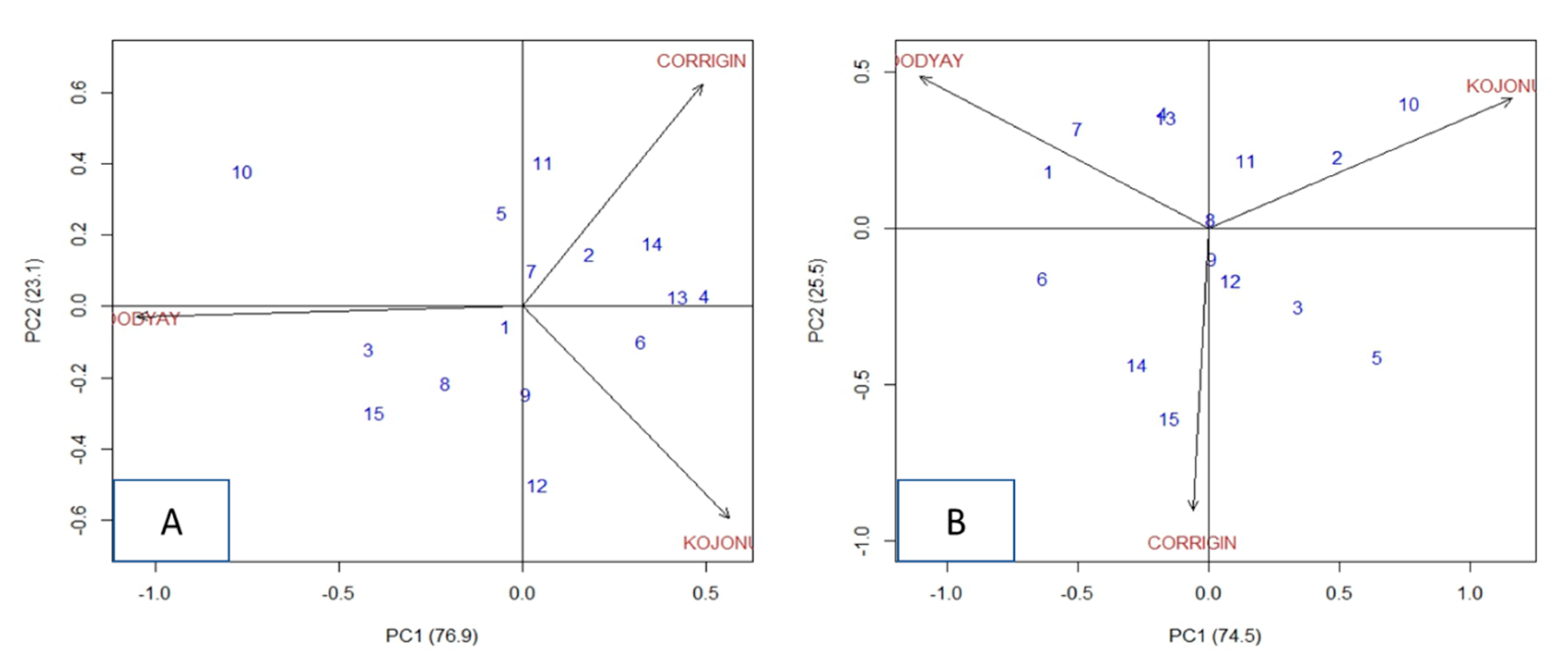
3.3. Environmental and Allelic Combination Effects on Yield and Protein Content
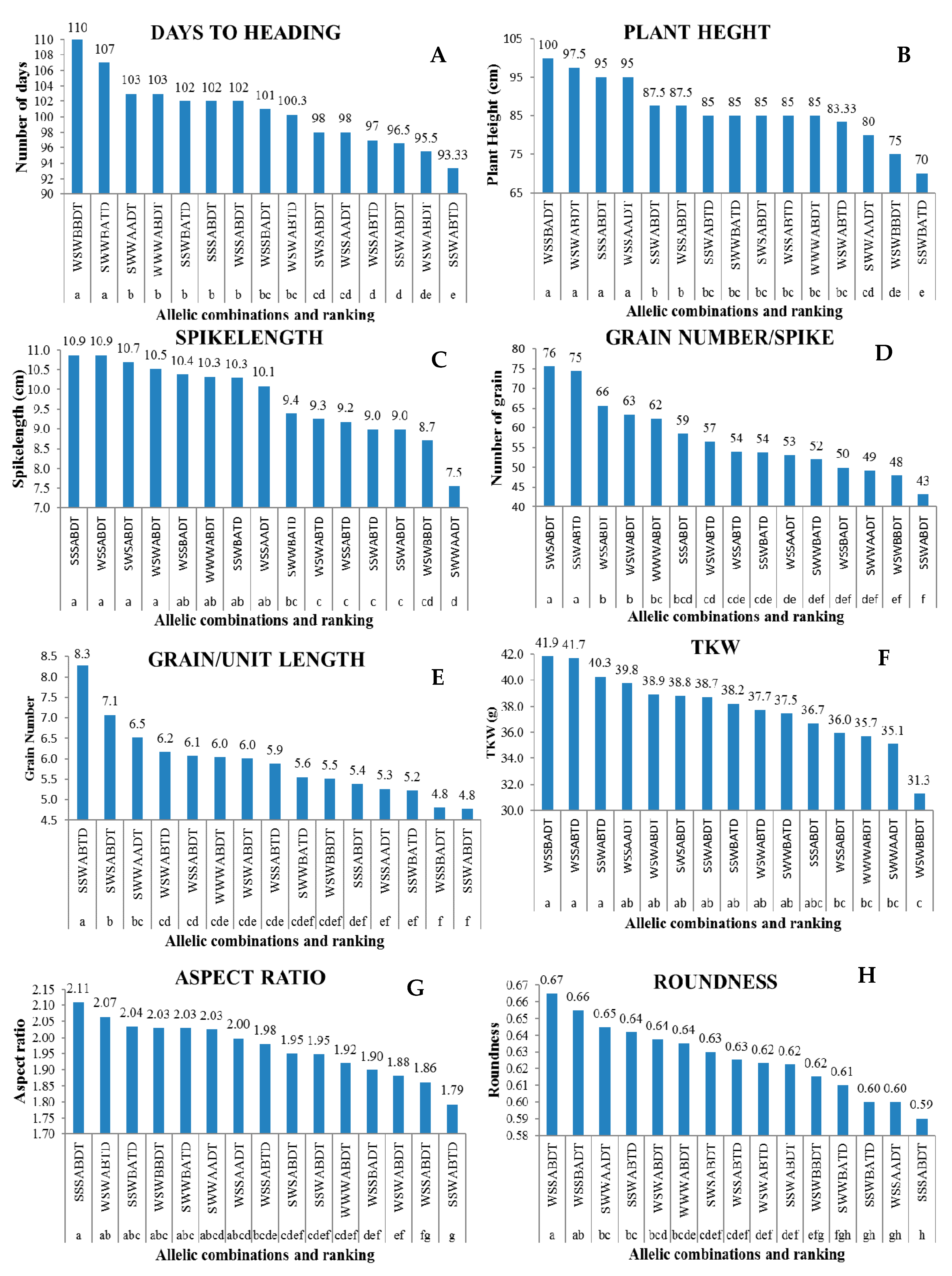
3.4. Allelic Combination Effects on Agronomic Traits
4. Discussion
4.1. Allelic Diversity in the Advanced Lines Used
4.2. Allelic Combination Effects on Agronomic Traits
4.3. Environment and Allelic Combination Effects on Yield and Protein Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- GRDC. 2015. Available online: https://grdc.com.au/__data/assets/pdf_file/0021/240717/GRDC-GrowNotes-Wheat-Western.pdf (accessed on 23 September 2020).
- Turner, N.C.; Molyneux, N.; Yang, S.; Xiong, Y.; Siddique, K.H.M. Climate change in south-west Australia and north-west China: Challenges and opportunities for crop production. Crop. Pasture Sci. 2011, 62, 445–456. [Google Scholar] [CrossRef]
- Pugsley, A.T. The impact of plant physiology on Australian wheat breeding. Euphytica 1983, 32, 743–748. [Google Scholar] [CrossRef]
- Turner, N.C. Sustainable production of crops and pastures under drought in a Mediterranean environment. Ann. Appl. Biol. 2004, 144, 139–147. [Google Scholar] [CrossRef]
- Eagles, H.A.; Cane, K.; Trevaskis, B. Veery wheats carry an allele of Vrn-A1 that has implications for freezing tolerance in winter wheats. Plant Breed. 2011, 130, 413–418. [Google Scholar] [CrossRef]
- Reynolds, M.P. Climate Change and Crop Production; CABI: Wallingford, UK, 2010. [Google Scholar]
- Law, C.N.; Sutka, J.; Worland, A.J. A Genetic study of day-length response in wheat. Heredity 1978, 41, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Pugsley, A. A genetic analysis of the spring-winter habit of growth in wheat. Aust. J. Agric. Res. 1971, 22, 21–31. [Google Scholar] [CrossRef]
- Beales, J.; Turner, A.; Griffiths, S.; Snape, J.W.; Laurie, D.A. A Pseudo-Response Regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 115, 721–733. [Google Scholar] [CrossRef]
- Chen, Y.; Carver, B.F.; Wang, S.; Zhang, F.; Yan, L. Genetic loci associated with stem elongation and winter dormancy release in wheat. Theor. Appl. Genet. 2009, 118, 881–889. [Google Scholar] [CrossRef]
- Díaz, A.; Zikhali, M.; Turner, A.S.; Isaac, P.; Laurie, D.A. Copy Number Variation Affecting the Photoperiod-B1 and Vernalization-A1 Genes Is Associated with Altered Flowering Time in Wheat (Triticum aestivum). PLoS ONE 2012, 7, e33234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulkes, M.; Sylvester-Bradley, R.; Worland, A.; Snape, J. Effects of a photoperiod-response gene Ppd-D1 on yield potential and drought resistance in UK winter wheat. Euphytica 2004, 135, 63–73. [Google Scholar] [CrossRef]
- Guo, Z.; Song, Y.; Zhou, R.; Ren, Z.; Jia, J. Discovery, evaluation and distribution of haplotypes of the wheat Ppd-D1 gene. New Phytol. 2010, 185, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Muterko, A.F.; Kalendar, R.; Cockram, J.; Balashova, I. Discovery, evaluation and distribution of haplotypes and new alleles of the Photoperiod-A1 gene in wheat. Plant Mol. Biol. 2015, 88, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Nishida, H.; Yoshida, T.; Kawakami, K.; Fujita, M.; Long, B.; Akashi, Y.; Laurie, D.; Kato, K. Structural variation in the 5’ upstream region of photoperiod-insensitive alleles Ppd-A1a and Ppd-B1a identified in hexaploid wheat (Triticum aestivum L.), and their effect on heading time. Mol. Breed. 2013, 31, 27–37. [Google Scholar] [CrossRef]
- Worland, A.J. The influence of flowering time genes on environmental adaptability in European wheats. Euphytica 1996, 89, 49–57. [Google Scholar] [CrossRef]
- Yan, L.; Fu, D.; Li, C.; Blechl, A.; Tranquilli, G.; Bonafede, M.; Sanchez, A.; Valarik, M.; Yasuda, S.; Dubcovsky, J. The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc. Natl. Acad. Sci. USA 2006, 103, 19581–19586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Loukoianov, A.; Blechl, A.; Tranquilli, G.; Ramakrishna, W.; SanMiguel, P.; Bennetzen, J.L.; Echenique, V.C.; Dubcovsky, J. The Wheat VRN2 Gene Is a Flowering Repressor Down-Regulated by Vernalization. Science 2004, 303, 1640–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Helguera, M.; Kato, K.; Fukuyama, S.; Sherman, J.; Dubcovsky, J. Allelic variation at the VRN-1 promoter region in polyploid wheat. Theor. Appl. Genet. 2004, 109, 1677–1686. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef] [Green Version]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of Flowering Time. Plant Cell 2002, 14, S111–S130. [Google Scholar] [CrossRef] [Green Version]
- Wellmer, F.; Riechmann, J.L. Gene networks controlling the initiation of flower development. Trends Genet. 2010, 26, 519–527. [Google Scholar] [CrossRef]
- Stelmakh, A.F. Genetic systems regulating flowering response in wheat. Euphytica 1998, 100, 359–369. [Google Scholar] [CrossRef]
- Grogan, S.M.; Brown-Guedira, G.; Haley, S.D.; McMaster, G.S.; Reid, S.D.; Smith, J.; Byrne, P.F. Allelic Variation in Developmental Genes and Effects on Winter Wheat Heading Date in the USA Great Plains. PLoS ONE 2016, 11, e0152852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eagles, H.A.; Cane, K.; Vallance, N. The flow of alleles of important photoperiod and vernalisation genes through Australian wheat. Crop. Pasture Sci. 2009, 60, 646–657. [Google Scholar] [CrossRef]
- Fu, D.; Szűcs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; Von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, M.; Spielmeyer, W.; Gale, K.; Rebetzke, G.; Richards, R. “Perfect” markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat. Theor. Appl. Genet. 2002, 105, 1038–1042. [Google Scholar] [CrossRef]
- Cane, K.; Eagles, H.A.; Laurie, D.A.; Trevaskis, B.; Vallance, N.; Eastwood, R.F.; Gororo, N.N.; Kuchel, H.; Martin, P.J. Ppd-B1 and Ppd-D1 and their effects in southern Australian wheat. Crop. Pasture Sci. 2013, 64, 100–114. [Google Scholar] [CrossRef]
- Smith, A.B.; Cullis, B.R.; Thompson, R. The analysis of crop cultivar breeding and evaluation trials: An overview of current mixed model approaches. J. Agric. Sci. 2005, 143, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Butler, D.; Cullis, B.; Gilmour, A.; Gogel, B.; Thompson, R. ASReml-R reference manual (version 3). In The State of Queensland; Department of Primary Industries and Fisheries: Brisbane, QLD, Australia, 2009. [Google Scholar]
- Purchase, J.L.; Hatting, H.; van Deventer, C.S. Genotype × environment interaction of winter wheat (Triticum aestivum L.) in South Africa: II. Stability analysis of yield performance. S. Afr. J. Plant Soil 2000, 17, 101–107. [Google Scholar] [CrossRef]
- Eberhart, S.A.; Russell, W.A. Stability Parameters for Comparing Varieties 1. Crop. Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef] [Green Version]
- R Foundation for Statistical Computing. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 13 July 2017).
- Shaw, L.M.; Turner, A.S.; Laurie, D.A. The impact of photoperiod insensitive Ppd-1a mutations on the photoperiod pathway across the three genomes of hexaploid wheat (Triticum aestivum). Plant J. 2012, 71, 71–84. [Google Scholar] [CrossRef]
- Bentley, A.R.; Turner, A.S.; Gosman, N.; Leigh, F.J.; Maccaferri, M.; Dreisigacker, S.; Greenland, A.; Laurie, D.A. Frequency of photoperiod-insensitive Ppd-A1a alleles in tetraploid, hexaploid and synthetic hexaploid wheat germplasm. Plant Breed. 2011, 130, 10–15. [Google Scholar] [CrossRef]
- Edwards, I.; Murdoch University, Murdoch, WA, Australia. Personal communication, 2020.
- Allard, V.; Veisz, O.; Kõszegi, B.; Rousset, M.; Le Gouis, J.; Martre, P. The quantitative response of wheat vernalization to environmental variables indicates that vernalization is not a response to cold temperature. J. Exp. Bot. 2011, 63, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Eagles, H.A.; Cane, K.; Kuchel, H.; Hollamby, G.J.; Vallance, N.; Eastwood, R.F.; Gororo, N.N.; Martin, P.J. Photoperiod and vernalization gene effects in southern Australian wheat. Crop. Pasture Sci. 2010, 61, 721–730. [Google Scholar] [CrossRef]
- Dowla, M.A.N.N.U.; Edwards, I.; O’Hara, G.; Islam, S.; Ma, W. Developing Wheat for Improved Yield and Adaptation Under a Changing Climate: Optimization of a Few Key Genes. Engineering 2018, 4, 514–522. [Google Scholar] [CrossRef]
- Eagles, H.A.; Cane, K.; Trevaskis, B.; Vallance, N.; Eastwood, R.F.; Gororo, N.N.; Kuchel, H.; Martin, P.J. Ppd1, Vrn1, ALMT1 and Rht genes and their effects on grain yield in lower rainfall environments in southern Australia. Crop. Pasture Sci. 2014, 65, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, M.; Navabi, A.; Salmon, D.F.; Yang, R.-C.; Murdoch, B.M.; Moore, S.S.; Spaner, D. Genetic analysis of flowering and maturity time in high latitude spring wheat. Euphytica 2007, 154, 207–218. [Google Scholar] [CrossRef]
- Worland, A.J.; Börner, A.; Korzun, V.; Li, W.; Petrović, S.; Sayers, E.J. The influence of photoperiod genes on the adaptability of European winter wheats. Euphytica 1998, 100, 385–394. [Google Scholar] [CrossRef]
- Dubcovsky, J.; Lijavetzky, D.; Appendino, L.; Tranquilli, G. Comparative RFLP mapping of Triticum monococcum genes controlling vernalization requirement. Theor. Appl. Genet. 1998, 97, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.; Dunford, R.P.; Coupland, G.; Laurie, D.A. The Evolution of CONSTANS-Like Gene Families in Barley, Rice, and Arabidopsis1. Plant Physiol. 2003, 131, 1855–1867. [Google Scholar] [CrossRef] [Green Version]
- Snape, J.W.; Sarma, R.; Quarrie, S.; Fish, L.; Galiba, G.; Sutka, J. Mapping genes for flowering time and frost tolerance in cereals using precise genetic stocks. Euphytica 2001, 120, 309–315. [Google Scholar] [CrossRef]
- Wilhelm, E.P.; Boulton, M.I.; Al-Kaff, N.; Balfourier, F.; Bordes, J.; Greenland, A.J.; Powell, W.; Mackay, I. Rht-1 and Ppd-D1 associations with height, GA sensitivity, and days to heading in a worldwide bread wheat collection. Theor. Appl. Genet. 2013, 126, 2233–2243. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Huang, Z.; Zhu, Q.; Wang, L. Remobilization of Carbon Reserves Is Improved by Controlled Soil-Drying during Grain Filling of Wheat. Crop. Sci. 2000, 40, 1645–1655. [Google Scholar] [CrossRef]
- Richards, R. The effect of dwarfing genes in spring wheat in dry environments. II. Growth, water use and water-use efficiency. Aust. J. Agric. Res. 1992, 43, 529–539. [Google Scholar] [CrossRef]
- Fischer, R.A. Number of kernels in wheat crops and the influence of solar radiation and temperature. J. Agric. Sci. 1985, 105, 447–461. [Google Scholar] [CrossRef]
- Fischer, R.A. Wheat physiology: A review of recent developments. Crop. Pasture Sci. 2011, 62, 95–114. [Google Scholar] [CrossRef] [Green Version]
- Foulkes, M.J.; Slafer, G.A.; Davies, W.J.; Berry, P.M.; Sylvester-Bradley, R.; Martre, P.; Calderini, D.F.; Griffiths, S.; Reynolds, M.P. Raising yield potential of wheat. III. Optimizing partitioning to grain while maintaining lodging resistance. J. Exp. Bot. 2011, 62, 469–486. [Google Scholar] [CrossRef] [Green Version]
- Abbate, P.; Pontaroli, A.C.; Lázaro, L.; Gutheim, F. A method of screening for spike fertility in wheat. J. Agric. Sci. 2013, 151, 322–330. [Google Scholar] [CrossRef]
- Alwala, S.; Kwolek, T.; McPherson, M.; Pellow, J.; Meyer, D. A comprehensive comparison between Eberhart and Russell joint regression and GGE biplot analyses to identify stable and high yielding maize hybrids. Field Crop. Res. 2010, 119, 225–230. [Google Scholar] [CrossRef]
- Grausgruber, H.; Oberforster, M.; Werteker, M.; Ruckenbauer, P.; Vollmann, J. Stability of quality traits in Austrian-grown winter wheats. Field Crop. Res. 2000, 66, 257–267. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SL. NO. | ALLELIC COMBINATION i | Line Name | Vernalization Loci Suffix a and c = “S” and v = “W” | Photoperiod Loci Suffix a and c = “A” and b = “B” | Reduced Height Loci Suffix a = “T” and b = “D” | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| VRN A1 | VRN B1 | VRN D1 | PPD-A1 | PPD-D1 | PPD-B1 | Rht-1 | Rht-2 | |||
| 1 | SSS-AB-DT | CMT-4 | Vrn-A1a | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a |
| 2 | SSW-AB-DT | QLD-4 | Vrn-A1a | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a |
| Bonnie Rock | Vrn-A1a | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a | ||
| 3 | SSW-AB-TD | SP-2 | Vrn-A1a | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b |
| VIC-1 | Vrn-A1a | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b | ||
| VIC-2 | Vrn-A1a | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b | ||
| VIC-3 | Vrn-A1c | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b | ||
| 4 | SSW-BA-TD | UK-2 | Vrn-A1a | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1b | Ppd-B1a | Rht-B1a | Rht-D1b |
| 5 | SWS-AB-DT | SP-1 | Vrn-A1a | Vrn-B1v | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a |
| 6 | SWW-AA-DT | UK-4 | Vrn-A1a | Vrn-B1v | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1c | Rht-B1b | Rht-D1a |
| 7 | SWW-BA-TD | UK-1 | Vrn-A1a | Vrn-B1v | Vrn-D1v | Ppd-A1b | Ppd-D1b | Ppd-B1a | Rht-B1a | Rht-D1b |
| 8 | WSS-AA-DT | SP-4 | Vrn-A1v | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1a | Rht-B1b | Rht-D1a |
| CMT-3 | Vrn-A1v | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1a | Rht-B1b | Rht-D1a | ||
| 9 | WSS-AB-DT | CMT-2 | Vrn-A1v | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a |
| QLD-3 | Vrn-A1v | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a | ||
| 10 | WSS-AB-TD | Mace | Vrn-A1v | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b |
| 11 | WSS-BA-DT | CMT-1 | Vrn-A1v | Vrn-B1a | Vrn-D1a | Ppd-A1b | Ppd-D1b | Ppd-B1a | Rht-B1b | Rht-D1a |
| 12 | WSW-AB-DT | QLD-2 | Vrn-A1v | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a |
| SP-3 | Vrn-A1v | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1a | ||
| 13 | WSW-AB-TD | Wyalkatchem | Vrn-A1v | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b |
| Magenta | Vrn-A1v | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1a | Rht-D1b | ||
| 14 | WSW-BB-DT | UK-3 | Vrn-A1v | Vrn-B1a | Vrn-D1v | Ppd-A1b | Ppd-D1b | Ppd-B1b | Rht-B1b | Rht-D1a |
| 15 | WWW-AB-DT | QLD-1 | Vrn-A1v | Vrn-B1v | Vrn-D1v | Ppd-A1b | Ppd-D1a | Ppd-B1b | Rht-B1b | Rht-D1 i |
| Pool | Descriptions | Genetic Background |
|---|---|---|
| Spanish/French (SP) | Biparental cross | Califa Sur, Rinconda, Farak, Arrturnik, Fidel, Soissons, Recital, Monopol |
| Winter x Spring (UK) | Three-way | Winter: Einstein, WW66, Heperion (from Europe) Spring: Correl, Carinya, VP1081, Sunzell, Sunstate (Australia) |
| Synthetic Hexaploid (VIC) | Multi-parental | Pavon, 30271, TM56, Janz, Annuello, Aegilops squarrosa |
| CIMMYT (CMT) | Multi-parental and wide crosses | Kiritati, Waxwing, Onix, Tacupeto, Pastor, CRBD-3, Stork, T. diccoides |
| Queensland (QLD) | Recurrent selection | Seri, Batavia, Kukri, Sunstate, Janz, Hartog |
| Sl No. | ALLELIC COMBINATION | Number of Lines | YIELD (ton/Ha) | PROTEIN (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Corrigin | Kojonup | Toodyay | Corrigin | Kojonup | Toodyay | |||
| 1 | SSSABDT | 1 (CMT-4) | 2.84 | 4.87 | 3.78 | 10.53 | 11.57 | 12.72 |
| 2 | SSWABDT | 2 (QLD-4 and Bonnie Rock | 2.98 | 4.75 | 3.42 | 10.32 | 12.77 | 11.36 |
| 3 | SSWABTD | 4 (SP-2, VIC-1, VIC-2 and VIC-3) | 3.19 | 5.27 | 4.74 | 10.24 | 11.81 | 10.74 |
| 4 | SSWBATD | 1 (UK-2) | 3.37 | 5.32 | 3.41 | 9.87 | 11.70 | 11.79 |
| 5 | SWSABDT | 1 (SP-1) | 3.00 | 4.60 | 3.73 | 11.57 | 13.27 | 11.42 |
| 6 | SWSAADT | 1 (UK-4) | 3.04 | 5.15 | 3.43 | 10.20 | 10.70 | 11.90 |
| 7 | SWWBATD | 1 (UK-1) | 2.95 | 4.77 | 3.66 | 9.80 | 11.17 | 11.83 |
| 8 | WSSAADT | 2 (SP-4 and CMT-3) | 2.70 | 4.93 | 4 | 10.42 | 11.98 | 11.65 |
| 9 | WSSABDT | 2 (CMT-2 and QLD-3) | 2.90 | 5.19 | 3.89 | 10.53 | 11.92 | 11.60 |
| 10 | WSSABTD | 1 (Mace) | 3.65 | 5.04 | 5.39 | 9.50 | 12.53 | 10.48 |
| 11 | WSSBADT | 1 (CMT-1) | 3.15 | 4.56 | 3.59 | 9.93 | 11.93 | 11.33 |
| 12 | WSWABDT | 2 (QLD-2 and SP-3) | 2.74 | 5.37 | 3.87 | 10.50 | 11.87 | 11.33 |
| 13 | WSWABTD | 2 (Wyalkatchem and Magenta) | 3.20 | 5.15 | 3.35 | 10.26 | 12.08 | 12.25 |
| 14 | WSWBBDT | 1 (UK-3) | 2.88 | 4.62 | 3.03 | 10.37 | 10.90 | 11.26 |
| 15 | WWWABDT | 1 (QLD-1) | 2.33 | 4.66 | 4 | 10.70 | 11.13 | 11.09 |
| Standard Error of Difference (SED)± | 0.2929 | 0.6729 | ||||||
| Allelic Combination | Yield (ton/Ha) | Protein | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sl No | ASV (Rank) | S2di | bi | Mean | GEI | ASV (Rank) | S2di | bi | Means | GEI | |||||
| CORRIGIN | KOJONUP | TOODYAY | CORRIGIN | KOJONUP | TOODYAY | ||||||||||
| 1 | SSSABDT | 0.101 (1) | 0.080 | 1.033 | 3.83 | −0.062 | 0.011 | 0.050 | 1.054 (12) | 0.773 | 1.038 | 11.611 | −0.150 | −0.622 | 0.772 |
| 2 | SSWABDT | 0.362 (5) | 0.022 | 0.922 | 3.72 | 0.190 | 0.006 | −0.196 | 0.877 (10) | 0.303 | 1.371 | 11.506 | −0.261 | 0.683 | −0.423 |
| 3 | SSWABTD | 0.773 (12) | 0.219 | 1.031 | 4.40 | −0.288 | −0.157 | 0.445 | 0.642 (8) | 0.134 | 0.828 | 10.944 | 0.228 | 0.289 | −0.517 |
| 4 | SSWBATD | 0.905 (14) | 0.324 | 1.040 | 4.03 | 0.264 | 0.260 | −0.524 | 0.476 (6) | 0.100 | 1.344 | 11.156 | −0.361 | −0.033 | 0.394 |
| 5 | SWSABDT | 0.285 (4) | 0.083 | 0.812 | 3.78 | 0.155 | −0.207 | 0.053 | 1.182 (14) | 1.021 | 0.736 | 12.122 | 0.373 | 0.567 | −0.940 |
| 6 | SWSAADT | 0.598 (9) | 0.078 | 1.105 | 3.87 | 0.092 | 0.246 | −0.338 | 1.094 (13) | 0.703 | 0.671 | 10.933 | 0.195 | −0.811 | 0.616 |
| 7 | SWWBATD | 0.112 (2) | 0.083 | 0.934 | 3.79 | 0.083 | −0.052 | −0.031 | 0.913 (11) | 0.514 | 1.211 | 11.022 | −0.294 | −0.433 | 0.727 |
| 8 | WSSAADT | 0.437 (7) | 0.002 | 1.121 | 3.88 | −0.250 | 0.024 | 0.227 | 0.037 (1) | 0.230 | 1.037 | 11.378 | −0.033 | 0.028 | 0.005 |
| 9 | WSSABDT | 0.247 (3) | 0.084 | 1.169 | 3.99 | −0.165 | 0.168 | −0.003 | 0.099 (2) | 0.228 | 0.903 | 11.367 | 0.095 | −0.028 | −0.067 |
| 10 | WSSABTD | 1.449 (15) | 0.790 | 0.648 | 4.69 | −0.114 | −0.683 | 0.796 | 1.368 (15) | 1.019 | 1.622 | 10.878 | −0.450 | 1.078 | −0.628 |
| 11 | WSSBADT | 0.418 (6) | 0.070 | 0.730 | 3.77 | 0.308 | −0.236 | −0.072 | 0.327 (4) | 0.199 | 1.266 | 11.089 | −0.227 | 0.267 | −0.040 |
| 12 | WSWABDT | 0.505 (8) | 0.085 | 1.347 | 3.99 | −0.328 | 0.352 | −0.024 | 0.221 (3) | 0.200 | 0.847 | 11.267 | 0.162 | 0.022 | −0.184 |
| 13 | WSWABTD | 0.766 (11) | 0.209 | 1.031 | 3.90 | 0.226 | 0.217 | −0.444 | 0.454 (5) | 0.113 | 1.330 | 11.530 | −0.346 | −0.030 | 0.375 |
| 14 | WSWBBDT | 0.668 (10) | 0.140 | 0.922 | 3.51 | 0.295 | 0.083 | −0.378 | 0.641 (7) | 0.115 | 0.473 | 10.844 | 0.450 | −0.522 | 0.072 |
| 15 | WWWABDT | 0.798 (13) | 0.226 | 1.154 | 3.66 | −0.405 | −0.032 | 0.437 | 0.659 (9) | 0.220 | 0.323 | 11.011 | 0.617 | −0.455 | −0.162 |
| Mean | 3.92 | 11.24 | |||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dowla, M.A.N.N.U.; Islam, S.; Stefanova, K.; Hara, G.O.; Ma, W.; Edwards, I. Phenology and Dwarfing Gene Interaction Effects on the Adaptation of Selected Wheat (Triticum aestivum L.) Advanced Lines across Diverse Water-Limited Environments of Western Australia. Agriculture 2020, 10, 470. https://doi.org/10.3390/agriculture10100470
Dowla MANNU, Islam S, Stefanova K, Hara GO, Ma W, Edwards I. Phenology and Dwarfing Gene Interaction Effects on the Adaptation of Selected Wheat (Triticum aestivum L.) Advanced Lines across Diverse Water-Limited Environments of Western Australia. Agriculture. 2020; 10(10):470. https://doi.org/10.3390/agriculture10100470
Chicago/Turabian StyleDowla, Mirza A.N.N.U., Shahidul Islam, Katia Stefanova, Graham O’ Hara, Wujun Ma, and Ian Edwards. 2020. "Phenology and Dwarfing Gene Interaction Effects on the Adaptation of Selected Wheat (Triticum aestivum L.) Advanced Lines across Diverse Water-Limited Environments of Western Australia" Agriculture 10, no. 10: 470. https://doi.org/10.3390/agriculture10100470