
Assessment of the Interferon-Lambda-3 Polymorphism in the Antibody Response to COVID-19 in Older Adults Seropositive for CMV

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. SARS-CoV-2 Diagnosis
2.3. Blood Samples Collection
2.4. COVID-19 Vaccination
2.5. Determination of Specific IgG for SARS-CoV-2
2.6. Virus Neutralization Test (VNT)
2.7. Determination of Specific IgM and IgG for CMV
2.8. Genotyping for the Polymorphism in the IFN-Lambda (il28b Gene)
2.9. Statistical Analysis
3. Results
3.1. Sample Characterization
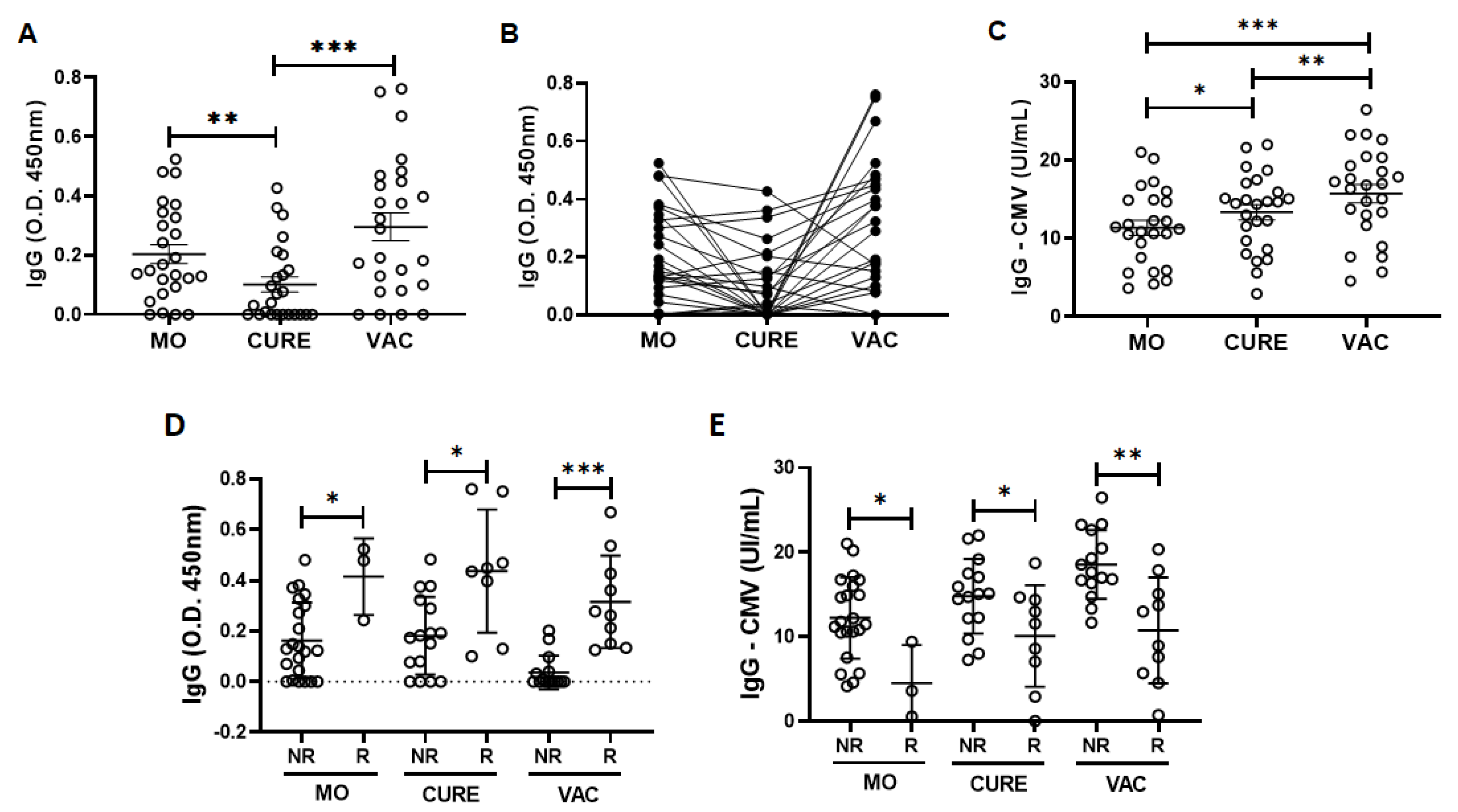
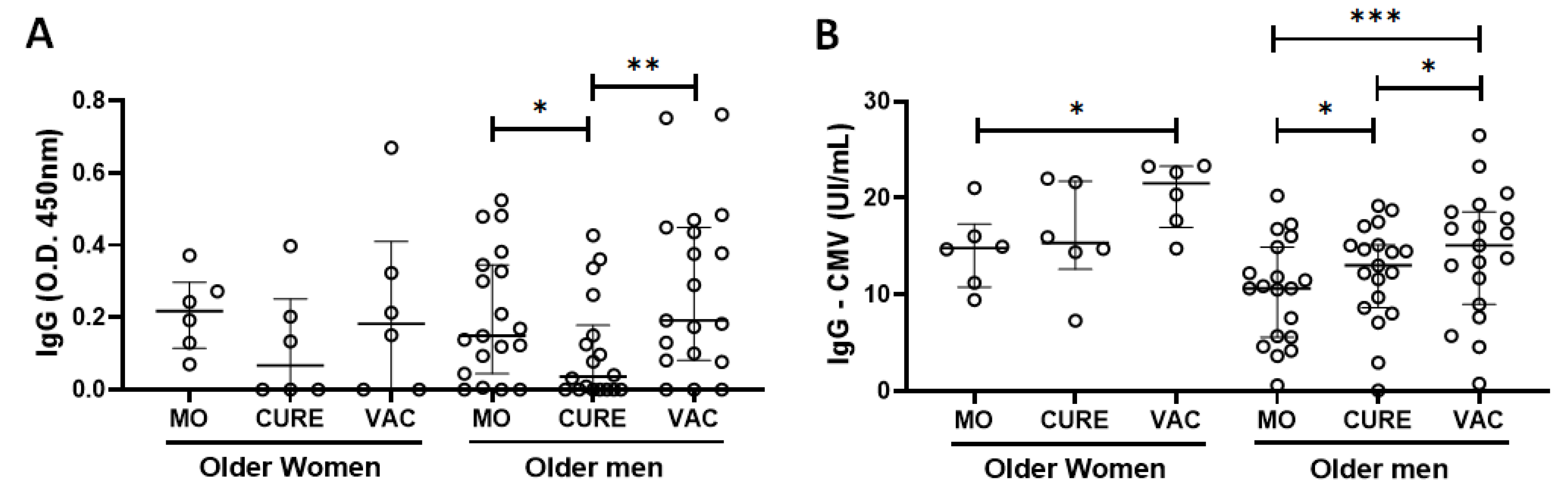
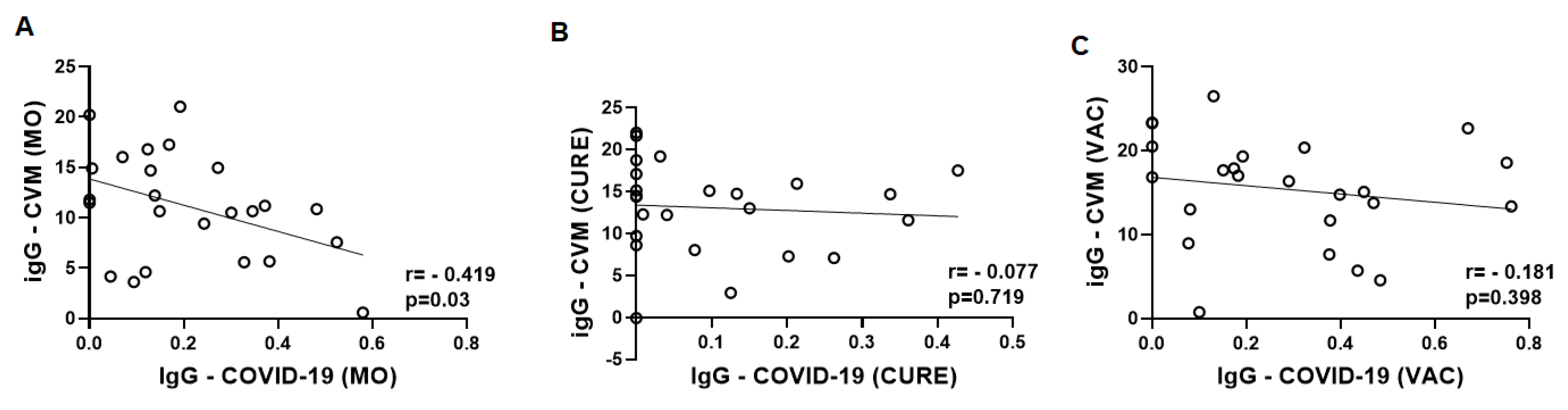
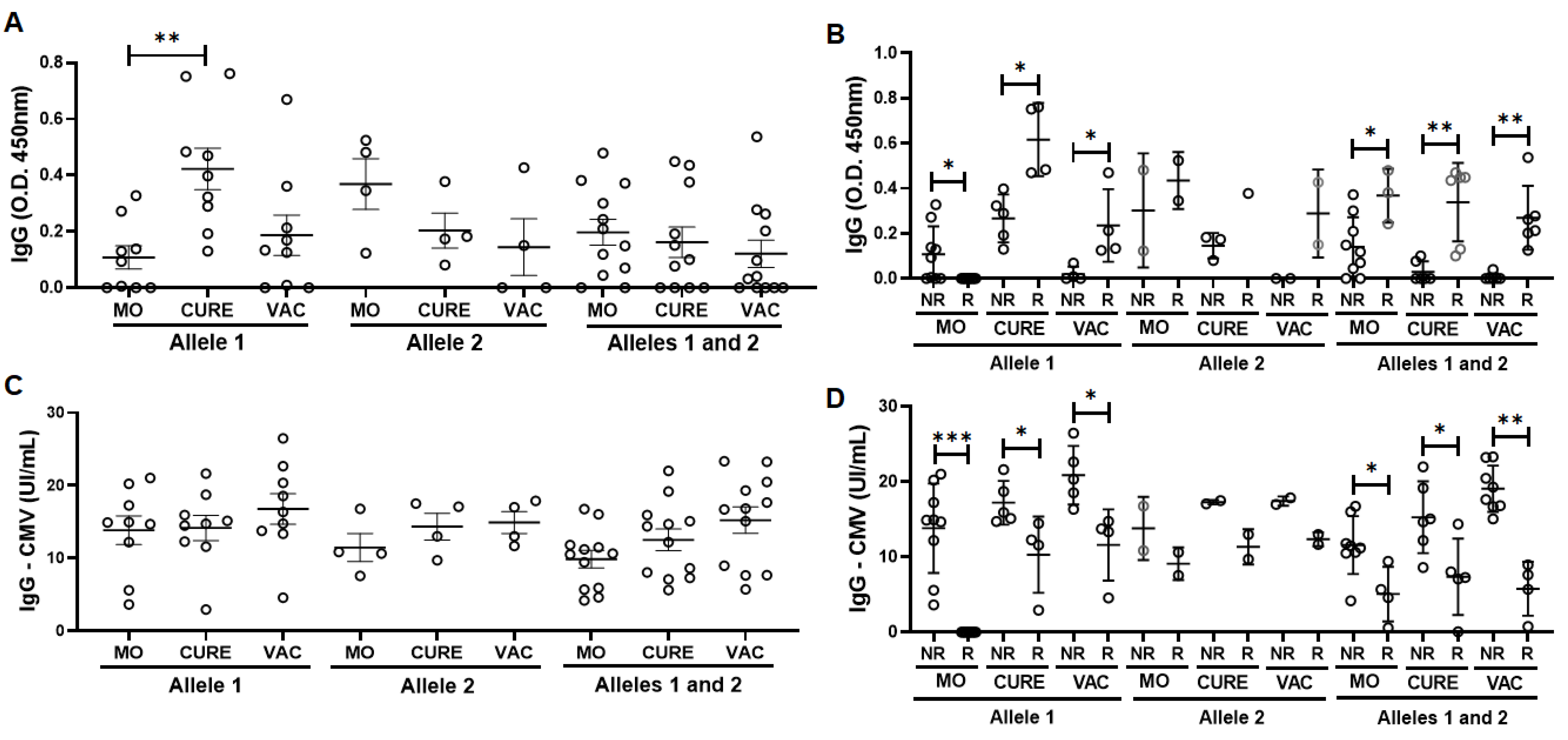
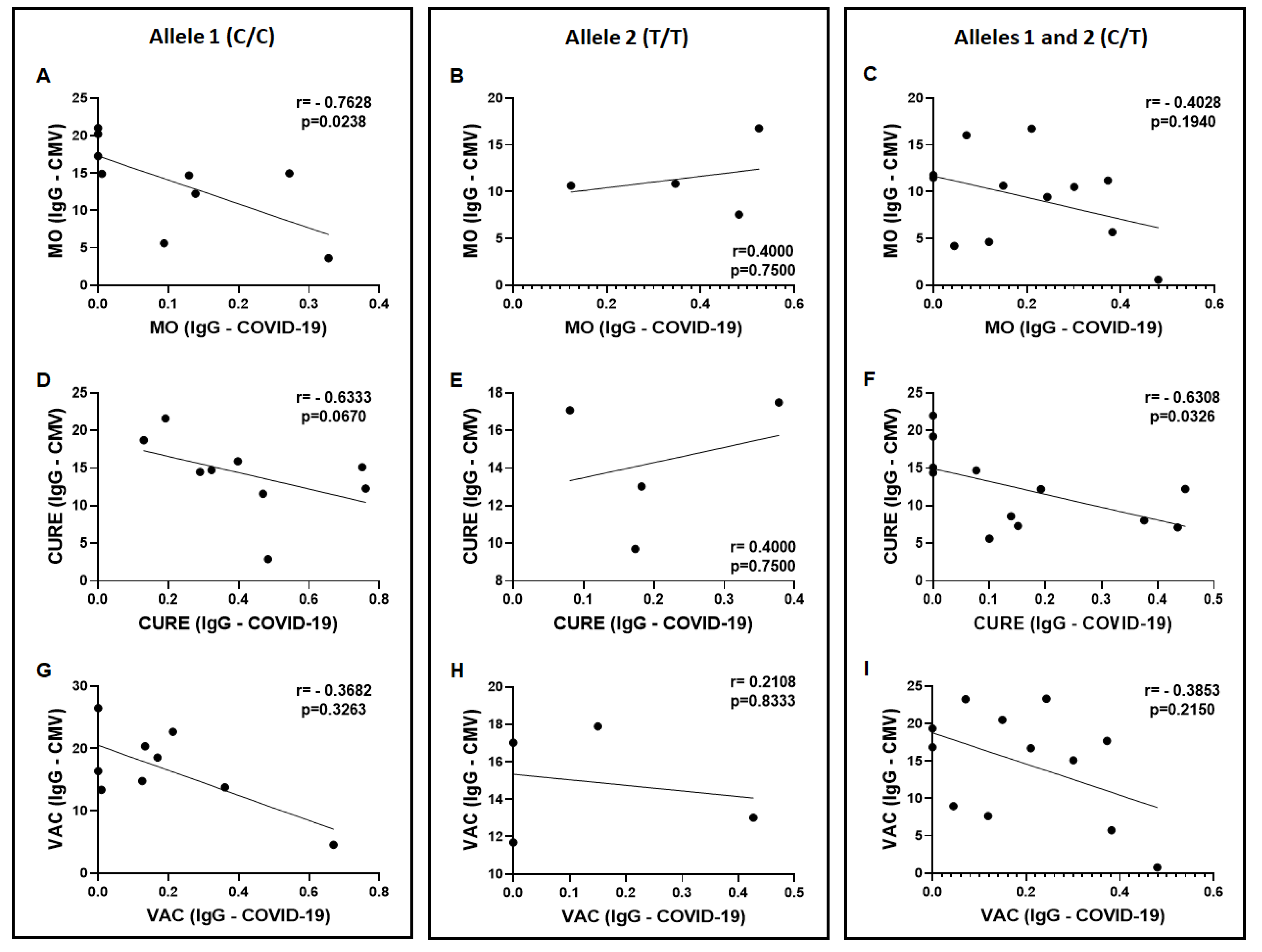
3.2. Specific IgG for the CMV and SARS-CoV-2 Antigens
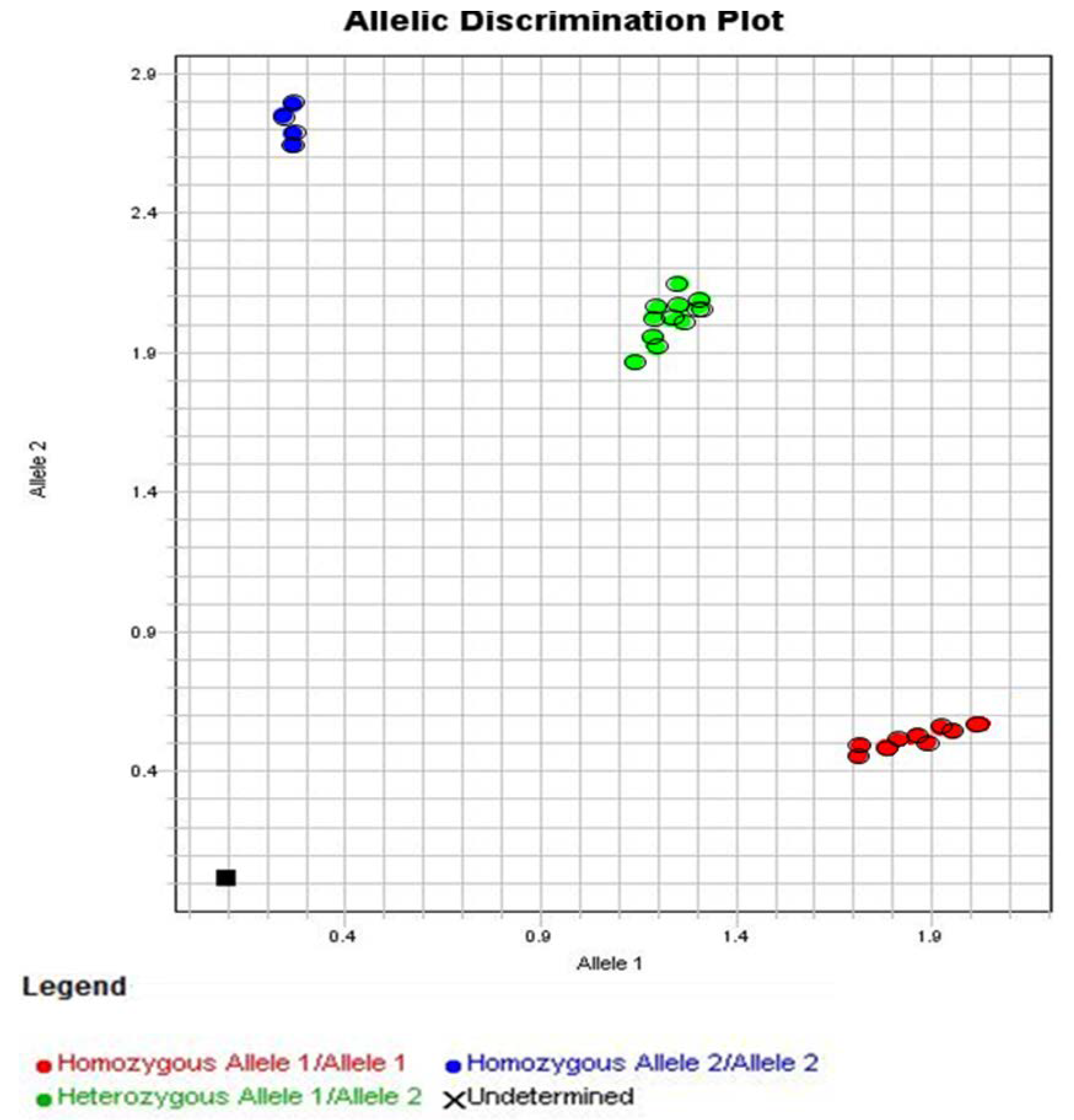
3.3. Allelic Discrimination of the Genotyping for the Polymorphism in IFN-Lambda (il28b Gene)
3.4. Specific IgG for the CMV and SARS-CoV-2 Antigens in the Volunteers Grouped Based on the Allelic Discrimination
4. Discussion
5. Limitations of the Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Brüggen, M.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune response to SARS-CoV-2 and mechanisms of immunopathological changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef] [PubMed]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Hu, N.; Lou, J.; Chen, K.; Kang, X.; Xiang, Z.; Chen, H.; Wang, D.; Liu, N.; Liu, D.; et al. Characteristics of COVID-19 infection in Beijing. J. Infect. 2020, 80, 401–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhavoronkov, A. Geroprotective and senoremediative strategies to reduce the comorbidity, infection rates, severity, and lethality in gerophilic and gerolavic infections. Aging 2020, 12, 6492–6510. [Google Scholar] [CrossRef] [PubMed]
- Alexeyev, M.F. Is there more to aging than mitochondrial DNA and reactive oxygen species? FEBS J. 2009, 276, 5768–5787. [Google Scholar] [CrossRef] [Green Version]
- Rokni, M.; Ghasemi, V.; Tavakoli, Z. Immune responses and pathogenesis ofSARS-CoV-2 during an outbreak in Iran: Comparison withSARSandMERS. Rev. Med. Virol. 2020, 30, e2107. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef]
- Dadras, O.; Afsahi, A.M.; Pashaei, Z.; Mojdeganlou, H.; Karimi, A.; Habibi, P.; Barzegary, A.; Fakhfouri, A.; Mirzapour, P.; Janfaza, N.; et al. The relationship between COVID-19 viral load and disease severity: A systematic review. Immun. Inflamm. Dis. 2021, 10, e580. [Google Scholar] [CrossRef]
- Chong, Y.; Ikematsu, H.; Yamaji, K.; Nishimura, M.; Nabeshima, S.; Kashiwagi, S.; Hayashi, J. CD27+ (memory) B cell decrease and apoptosis-resistant CD27- (naive) B cell increase in aged humans: Implications for age-related peripheral B cell developmental disturbances. Int. Immunol. 2005, 17, 383–390. [Google Scholar] [CrossRef]
- Chen, S.; Guan, F.; Candotti, F.; Benlagha, K.; Camara, N.O.S.; Herrada, A.A.; James, L.K.; Lei, J.; Miller, H.; Kubo, M.; et al. The role of B cells in COVID-19 infection and vaccination. Front. Immunol. 2022, 13, 988536. [Google Scholar] [CrossRef]
- Kovaiou, R.D.; Herndler-Brandstetter, D.; Grubeck-Loebenstein, B. Age-related changes in immunity: Implications for vaccination in the elderly. Expert Rev. Mol. Med. 2007, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Furtado, G.E.; Letieri, R.V.; Caldo-Silva, A.; Sardão, V.A.; Teixeira, A.M.; de Barros, M.P.; Vieira, R.P.; Bachi, A.L.L. Sustaining efficient immune functions with regular physical exercise in the COVID-19 era and beyond. Eur. J. Clin. Investig. 2021, 51, e13485. [Google Scholar] [CrossRef] [PubMed]
- Bachi, A.L.; Suguri, V.M.; Ramos, L.R.; Mariano, M.; Vaisberg, M.; Lopes, J.D.; Bachi, A.L.; Suguri, V.M.; Ramos, L.R.; Mariano, M.; et al. Increased production of autoantibodies and specific antibodies in response to influenza virus vaccination in physically active older individuals. Results Immunol. 2013, 3, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, K.; Wei, Y.; Szwajcer, A.; Rabbani, R.; Zarychanski, R.; Abou-Setta, A.M.; Mahmud, S.M. Efficacy and safety of high-dose influenza vaccine in elderly adults: A systematic review and meta-analysis. Vaccine 2017, 35, 2775–2780. [Google Scholar] [CrossRef]
- Weinberger, B. Vaccines and Vaccination against SARS-CoV-2: Considerations for the Older Population. Vaccines 2021, 9, 1435. [Google Scholar] [CrossRef]
- McDonald, I.; Murray, S.M.; Reynolds, C.J.; Altmann, D.M.; Boyton, R.J. Comparative systematic review and meta-analysis of reactogenicity, immunogenicity and efficacy of vaccines against SARS-CoV-2. NPJ Vaccines 2021, 6, 74. [Google Scholar] [CrossRef]
- Franceschi, C.; Zaikin, A.; Gordleeva, S.; Ivanchenko, M.; Bonifazi, F.; Storci, G.; Bonafè, M. Inflammaging 2018: An update and a model. Semin. Immunol. 2018, 40, 1–5. [Google Scholar] [CrossRef]
- Accardi, G.; Caruso, C. Immune-inflammatory responses in the elderly: An update. Immun. Ageing 2018, 15, 11. [Google Scholar] [CrossRef]
- Heath, J.J.; Grant, M.D. The Immune Response Against Human Cytomegalovirus Links Cellular to Systemic Senescence. Cells 2020, 9, 766. [Google Scholar] [CrossRef] [Green Version]
- Jergović, M.; Contreras, N.A.; Nikolich-Žugich, J. Impact of CMV upon immune aging: Facts and fiction. Med. Microbiol. Immunol. 2019, 208, 263–269. [Google Scholar] [CrossRef]
- Schmaltz, H.N.; Fried, L.P.; Xue, Q.-L.; Walston, J.; Leng, S.X.; Semba, R.D. Chronic Cytomegalovirus Infection and Inflammation Are Associated with Prevalent Frailty in Community-Dwelling Older Women. J. Am. Geriatr. Soc. 2005, 53, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G. Immunosenenescence: Role of cytomegalovirus. Exp. Gerontol. 2014, 54, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Derhovanessian, E.; Larbi, A.; Strindhall, J.; Wikby, A. Cytomegalovirus and human immunosenescence. Rev. Med. Virol. 2008, 19, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, J.; Parsons, R.; Botelho, F.; Millar, J.; McNeil, S.; Fulop, T.; McElhaney, J.; Andrew, M.K.; Walter, S.D.; Devereaux, P.J.; et al. Immune Biomarkers Predictive of Respiratory Viral Infection in Elderly Nursing Home Residents. PLoS ONE 2014, 9, e108481. [Google Scholar] [CrossRef] [PubMed]
- Fensterl, V.; Sen, G.C. Interferons and viral infections. Biofactors 2009, 35, 14–20. [Google Scholar] [CrossRef]
- Park, A.; Iwasaki, A. Type I and Type III Interferons—Induction, Signaling, Evasion, and Application to Combat COVID-19. Cell Host Microbe 2020, 27, 870–878. [Google Scholar] [CrossRef]
- Linari, S.; Aghemo, A.; Bartolozzi, D.; Santagostino, E.; Rumi, M.; Fognani, E.; Fasulo, M.R.; Gragnani, L.; Bruno, R.; Morfini, M.; et al. Interferon lambda 3 rs12979860 polymorphism in patients with haemophilia and HCV infection: A predictor of spontaneous viral clearance and sustained virological response. Thromb. Haemost. 2014, 111, 1067–1076. [Google Scholar] [CrossRef]
- Bellanti, F.; Vendemiale, G.; Altomare, E.; Serviddio, G. The Impact of Interferon Lambda 3 Gene Polymorphism on Natural Course and Treatment of Hepatitis C. J. Immunol. Res. 2012, 2012, 849373. [Google Scholar] [CrossRef] [Green Version]
- Laidlaw, S.M.; Dustin, L.B. Interferon Lambda: Opportunities, Risks, and Uncertainties in the Fight Against HCV. Front. Immunol. 2014, 5, 545. [Google Scholar] [CrossRef] [Green Version]
- Quarleri, J.; Delpino, M.V. Type I and III IFN-mediated antiviral actions counteracted by SARS-CoV-2 proteins and host inherited factors. Cytokine Growth Factor Rev. 2021, 58, 55–65. [Google Scholar] [CrossRef]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A mouse-adapted SARS-CoV-2 model for the evaluation of COVID-19 medical countermeasures. Nature 2020, 586, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; An, P.; Winkler, C.A. Host Genetics of Cytomegalovirus Pathogenesis. Front. Genet. 2019, 10, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, P. “The ancient and the new”: Is there an interaction between cytomegalovirus and SARS-CoV-2 infection? Immun. Ageing 2020, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Kadambari, S.; Klenerman, P.; Pollard, A.J. Why the elderly appear to be more severely affected by COVID-19: The potential role of immunosenescence and CMV. Rev. Med. Virol. 2020, 30, e2144. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.D.M.B.D.; Amaral, J.B.D.; França, C.N.; Monteiro, F.R.; Alvares-Saraiva, A.M.; Kalil, S.; Durigon, E.L.; Oliveira, D.B.L.; Rodrigues, S.S.; Heller, D.; et al. Distinct Immunological Profiles Help in the Maintenance of Salivary Secretory IgA Production in Mild Symptoms COVID-19 Patients. Front. Immunol. 2022, 13, 890887. [Google Scholar] [CrossRef]
- Araujo, D.B.; Machado, R.R.G.; Amgarten, D.E.; Malta, F.D.M.; De Araujo, G.G.; Monteiro, C.O.; Candido, E.D.; Soares, C.P.; De Menezes, F.G.; Pires, A.C.C.; et al. SARS-CoV-2 isolation from the first reported patients in Brazil and establishment of a coordinated task network. Memórias Do Inst. Oswaldo Cruz 2020, 115, e200342. [Google Scholar] [CrossRef]
- Oliveira, J.R.; Ruiz, C.M.R.; Machado, R.R.G.; Magawa, J.Y.; Daher, I.P.; Urbanski, A.H.; Schmitz, G.J.H.; Arcuri, H.A.; Ferreira, M.A.; Sasahara, G.L.; et al. Immunodominant antibody responses directed to SARS-CoV-2 hotspot mutation sites and risk of immune escape. Front. Immunol. 2023, 13, 1010105. [Google Scholar] [CrossRef]
- Nurtop, E.; Villarroel, P.M.S.; Pastorino, B.; Ninove, L.; Drexler, J.F.; Roca, Y.; Gake, B.; Dubot-Peres, A.; Grard, G.; Peyrefitte, C.; et al. Combination of ELISA screening and seroneutralisation tests to expedite Zika virus seroprevalence studies. Virol. J. 2018, 15, 192. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.-C.; Tiu, C.; Hu, Z.; Chen, V.C.-W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2–spike protein–protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Shen, C.; Wang, Z.; Zhao, F.; Yang, Y.; Li, J.; Yuan, J.; Wang, F.; Li, D.; Yang, M.; Xing, L.; et al. Treatment of 5 Critically Ill Patients With COVID-19 With Convalescent Plasma. JAMA 2020, 323, 1582. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, G.X.; Sasahara, G.L.; Magawa, J.Y.; Nunes, J.P.S.; Bruno, F.R.; Kuramoto, A.C.; Almeida, R.R.; Ferreira, M.A.; Scagion, G.P.; Candido, D.; et al. Reduced T Cell and Antibody Responses to Inactivated Coronavirus Vaccine Among Individuals Above 55 Years Old. Front. Immunol. 2022, 13, 812126. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.M.; da Silva, L.T.; Candido, D.; Zheng, Y.; Tiyo, B.T.; Ferreira, A.E.F.; Corrêa-Silva, S.; Scagion, G.P.; Leal, F.B.; Chalup, V.N.; et al. Salivary, serological, and cellular immune response to the CoronaVac vaccine in health care workers with or without previous COVID-19. Sci. Rep. 2022, 12, 10125. [Google Scholar] [CrossRef] [PubMed]
- Prokunina-Olsson, L.; Muchmore, B.; Tang, W.; Pfeiffer, R.M.; Park, H.; Dickensheets, H.; Hergott, D.; Porter-Gill, P.; Mumy, A.; Kohaar, I.; et al. A variant upstream of IFNL3 (IL28B) creating a new interferon gene IFNL4 is associated with impaired clearance of hepatitis C virus. Nat. Genet. 2013, 45, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Felismino, E.S.; Santos, J.M.B.; Rossi, M.; Santos, C.A.F.; Durigon, E.L.; Oliveira, D.B.L.; Thomazelli, L.M.; Monteiro, F.R.; Sperandio, A.; Apostólico, J.S.; et al. Better Response to Influenza Virus Vaccination in Physically Trained Older Adults Is Associated With Reductions of Cytomegalovirus-Specific Immunoglobulins as Well as Improvements in the Inflammatory and CD8+ T-Cell Profiles. Front. Immunol. 2021, 12, 713763. [Google Scholar] [CrossRef] [PubMed]
- Smetana, J.; Chlibek, R.; Shaw, J.; Splino, M.; Prymula, R. Influenza vaccination in the elderly. Hum. Vaccines Immunother. 2017, 14, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Derhovanessian, E.; Theeten, H.; Hähnel, K.; Van Damme, P.; Cools, N.; Pawelec, G. Cytomegalovirus-associated accumulation of late-differentiated CD4 T-cells correlates with poor humoral response to influenza vaccination. Vaccine 2013, 31, 685–690. [Google Scholar] [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Landin, A.M.; Blomberg, B.B. Cytomegalovirus (CMV) seropositivity decreases B cell responses to the influenza vaccine. Vaccine 2015, 33, 1433–1439. [Google Scholar] [CrossRef] [Green Version]
- Trzonkowski, P.; Myśliwska, J.; Szmit, E.; Wickiewicz, J.; Łukaszuk, K.; Brydak, L.B.; Machała, M.; Myśliwski, A. Association between cytomegalovirus infection, enhanced proinflammatory response and low level of anti-hemagglutinins during the anti-influenza vaccination—An impact of immunosenescence. Vaccine 2003, 21, 3826–3836. [Google Scholar] [CrossRef]
- Derhovanessian, E.; Maier, A.B.; Hähnel, K.; McElhaney, J.E.; Slagboom, E.P.; Pawelec, G. Latent Infection with Cytomegalovirus Is Associated with Poor Memory CD4 Responses to Influenza A Core Proteins in the Elderly. J. Immunol. 2014, 193, 3624–3631. [Google Scholar] [CrossRef] [Green Version]
- Furman, D.; Jojic, V.; Sharma, S.; Shen-Orr, S.S.; Angel, C.J.L.; Onengut-Gumuscu, S.; Kidd, B.A.; Maecker, H.T.; Concannon, P.; Dekker, C.L.; et al. Cytomegalovirus infection enhances the immune response to influenza. Sci. Transl. Med. 2015, 7, 281ra43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, C.M.; White, M.J.; Bottomley, C.; Lusa, C.; Galan, A.R.; Turner, S.E.G.; Goodier, M.; Riley, E.M. Impaired NK Cell Responses to Pertussis and H1N1 Influenza Vaccine Antigens in Human Cytomegalovirus-Infected Individuals. J. Immunol. 2015, 194, 4657–4667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, K.; Fulop, T.; Tedder, G.; Gentleman, B.; Garneau, H.; Meneilly, G.S.; Kleppinger, A.; Pawelec, G.; McElhaney, J.E. Cytomegalovirus Seropositivity Predicts a Decline in the T Cell But Not the Antibody Response to Influenza in Vaccinated Older Adults Independent of Type 2 Diabetes Status. Journals Gerontol. Ser. A 2016, 72, 1163–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elzen, W.P.D.; Vossen, A.; Cools, H.J.; Westendorp, R.G.; Kroes, A.; Gussekloo, J. Cytomegalovirus infection and responsiveness to influenza vaccination in elderly residents of long-term care facilities. Vaccine 2011, 29, 4869–4874. [Google Scholar] [CrossRef]
- Merani, S.; Kuchel, G.A.; Kleppinger, A.; McElhaney, J.E. Influenza vaccine-mediated protection in older adults: Impact of influenza infection, cytomegalovirus serostatus and vaccine dosage. Exp. Gerontol. 2017, 107, 116–125. [Google Scholar] [CrossRef]
- Simonnet, A.; Engelmann, I.; Moreau, A.-S.; Garcia, B.; Six, S.; El Kalioubie, A.; Robriquet, L.; Hober, D.; Jourdain, M. High incidence of Epstein–Barr virus, cytomegalovirus, and human-herpes virus-6 reactivations in critically ill patients with COVID-19. Infect. Dis. Now 2021, 51, 296–299. [Google Scholar] [CrossRef]
- Naendrup, J.-H.; Borrega, J.G.; Eichenauer, D.A.; Shimabukuro-Vornhagen, A.; Kochanek, M.; Böll, B. Reactivation of EBV and CMV in Severe COVID-19—Epiphenomena or Trigger of Hyperinflammation in Need of Treatment? A Large Case Series of Critically ill Patients. J. Intensive Care Med. 2021, 37, 1152–1158. [Google Scholar] [CrossRef]
- Saade, A.; Moratelli, G.; Azoulay, E.; Darmon, M. Herpesvirus reactivation during severe COVID-19 and high rate of immune defect. Infect. Dis. Now 2021, 51, 676–679. [Google Scholar] [CrossRef]
- Agwa, S.; Kamel, M.; Elghazaly, H.; Elsamee, A.A.; Hafez, H.; Girgis, S.; Elarab, H.E.; Ebeid, F.; Sayed, S.; Sherif, L.; et al. Association between Interferon-Lambda-3 rs12979860, TLL1 rs17047200 and DDR1 rs4618569 Variant Polymorphisms with the Course and Outcome of SARS-CoV-2 Patients. Genes 2021, 12, 830. [Google Scholar] [CrossRef]
- Müller, L.; Di Benedetto, S. How Immunosenescence and Inflammaging May Contribute to Hyperinflammatory Syndrome in COVID-19. Int. J. Mol. Sci. 2021, 22, 12539. [Google Scholar] [CrossRef]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current State of the Science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.R.; Thomas, D.L.; Jackson, S.S.; Prokunina-Olsson, L.; Donnelly, R.P.; Hartmann, R. Weak Induction of Interferon Expression by Severe Acute Respiratory Syndrome Coronavirus 2 Supports Clinical Trials of Interferon-λ to Treat Early Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Klinkhammer, J.; Schnepf, D.; Ye, L.; Schwaderlapp, M.; Gad, H.H.; Hartmann, R.; Garcin, D.; Mahlakõiv, T.; Staeheli, P. IFN-λ prevents influenza virus spread from the upper airways to the lungs and limits virus transmission. Elife 2018, 7, e33354. [Google Scholar] [CrossRef] [PubMed]
- Sommereyns, C.; Paul, S.; Staeheli, P.; Michiels, T. IFN-Lambda (IFN-λ) Is Expressed in a Tissue-Dependent Fashion and Primarily Acts on Epithelial Cells In Vivo. PLOS Pathog. 2008, 4, e1000017. [Google Scholar] [CrossRef] [PubMed]
- Roohi, A.; Soroosh, P. May interferon λ be a novel therapeutic approach against COVID-19? Med. Hypotheses 2020, 146, 110351. [Google Scholar] [CrossRef]
- Pestka, S.; Krause, C.D.; Walter, M.R. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef]
- Witte, K.; Witte, E.; Sabat, R.; Wolk, K. IL-28A, IL-28B, and IL-29: Promising cytokines with type I interferon-like properties. Cytokine Growth Factor Rev. 2010, 21, 237–251. [Google Scholar] [CrossRef]
- Bravo, D.; Solano, C.; Giménez, E.; Remigia, M.J.; Corrales, I.; Amat, P.; Navarro, D. Effect of the IL28B Rs12979860 C/T polymorphism on the incidence and features of active cytomegalovirus infection in allogeneic stem cell transplant patients. J. Med. Virol. 2013, 86, 838–844. [Google Scholar] [CrossRef]
- Saponi-Cortes, J.M.R.; Rivas, M.D.; Calle-Alonso, F.; Sanchez, J.F.; Costo, A.; Martin, C.; Zamorano, J. IFNL4 genetic variant can predispose to COVID-19. Sci. Rep. 2021, 11, 21185. [Google Scholar] [CrossRef]
- Tu, H.; Bao, J. IFNL4, ACE1, PKR, IFNG, MBL2 genetic polymorphisms and severe COVID-19: A protocol for systematic review and meta-analysis. Medicine 2022, 101, e29405. [Google Scholar] [CrossRef]
- Egli, A.; Levin, A.; Santer, D.; Joyce, M.; O’Shea, D.; Thomas, B.; Lisboa, L.; Barakat, K.; Bhat, R.; Fischer, K.P.; et al. Immunomodulatory Function of Interleukin 28B During Primary Infection With Cytomegalovirus. J. Infect. Dis. 2014, 210, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Chmelova, K.; Frankova, S.; Jirsa, M.; Neroldova, M.; Sticova, E.; Merta, D.; Senkerikova, R.; Trunecka, P.; Spicak, J.; Sperl, J. IL28B rs12979860 T allele protects against CMV disease in liver transplant recipients in the post-prophylaxis and late period. Transpl. Infect. Dis. 2019, 21, e13124. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Volunteers | p-Value | ||
|---|---|---|---|---|
| Total | Women | Men | ||
| (n = 25) | (n = 06) | (n = 19) | ||
| Age (years) | 75.8 ± 8.5 | 75.4 ± 8.4 | 77.0 ± 9.7 | 0.483 |
| Height (m) | 1.68 ± 0.09 | 1.58 ± 0.07 * | 1.71 ± 0.08 | 0.003 |
| Weight (kg) | 75.2 ± 10.9 | 63.3 ± 6.8 * | 79.1 ± 9.1 | 0.001 |
| BMI (kg/m2) | 26.5 ± 2.9 | 25.9 ± 2.5 | 26.9 ± 3.1 | 0.199 |
| Clinical characteristics (n) | ||||
| Cough | 5 | 1 | 4 | >0.05 |
| Fever | 3 | 0 | 3 | >0.05 |
| Sore throat | 1 | 0 | 1 | >0.05 |
| Coryza | 2 | 0 | 2 | >0.05 |
| Oxygen Desaturation | 2 | 1 | 1 | >0.05 |
| Respiratory distress | 2 | 1 | 1 | >0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nardy, A.; Camargo, C.T.S.; Oliveira, Y.F.C.d.; Silva, F.C.d.; Almeida, M.S.d.; Monteiro, F.R.; Silva, B.R.; Amaral, J.B.d.; Oliveira, D.B.L.; Durigon, E.L.; et al. Assessment of the Interferon-Lambda-3 Polymorphism in the Antibody Response to COVID-19 in Older Adults Seropositive for CMV. Vaccines 2023, 11, 480. https://doi.org/10.3390/vaccines11020480
Nardy A, Camargo CTS, Oliveira YFCd, Silva FCd, Almeida MSd, Monteiro FR, Silva BR, Amaral JBd, Oliveira DBL, Durigon EL, et al. Assessment of the Interferon-Lambda-3 Polymorphism in the Antibody Response to COVID-19 in Older Adults Seropositive for CMV. Vaccines. 2023; 11(2):480. https://doi.org/10.3390/vaccines11020480
Chicago/Turabian StyleNardy, Ariane, Camila Tussato Soares Camargo, Yasmim Faustina Castro de Oliveira, Fernanda Cristina da Silva, Millena Soares de Almeida, Fernanda Rodrigues Monteiro, Brenda Rodrigues Silva, Jônatas Bussador do Amaral, Danielle Bruna Leal Oliveira, Edison Luiz Durigon, and et al. 2023. "Assessment of the Interferon-Lambda-3 Polymorphism in the Antibody Response to COVID-19 in Older Adults Seropositive for CMV" Vaccines 11, no. 2: 480. https://doi.org/10.3390/vaccines11020480