
Yeast-Based Virus-like Particles as an Emerging Platform for Vaccine Development and Delivery


Abstract
:1. Background of Vaccines
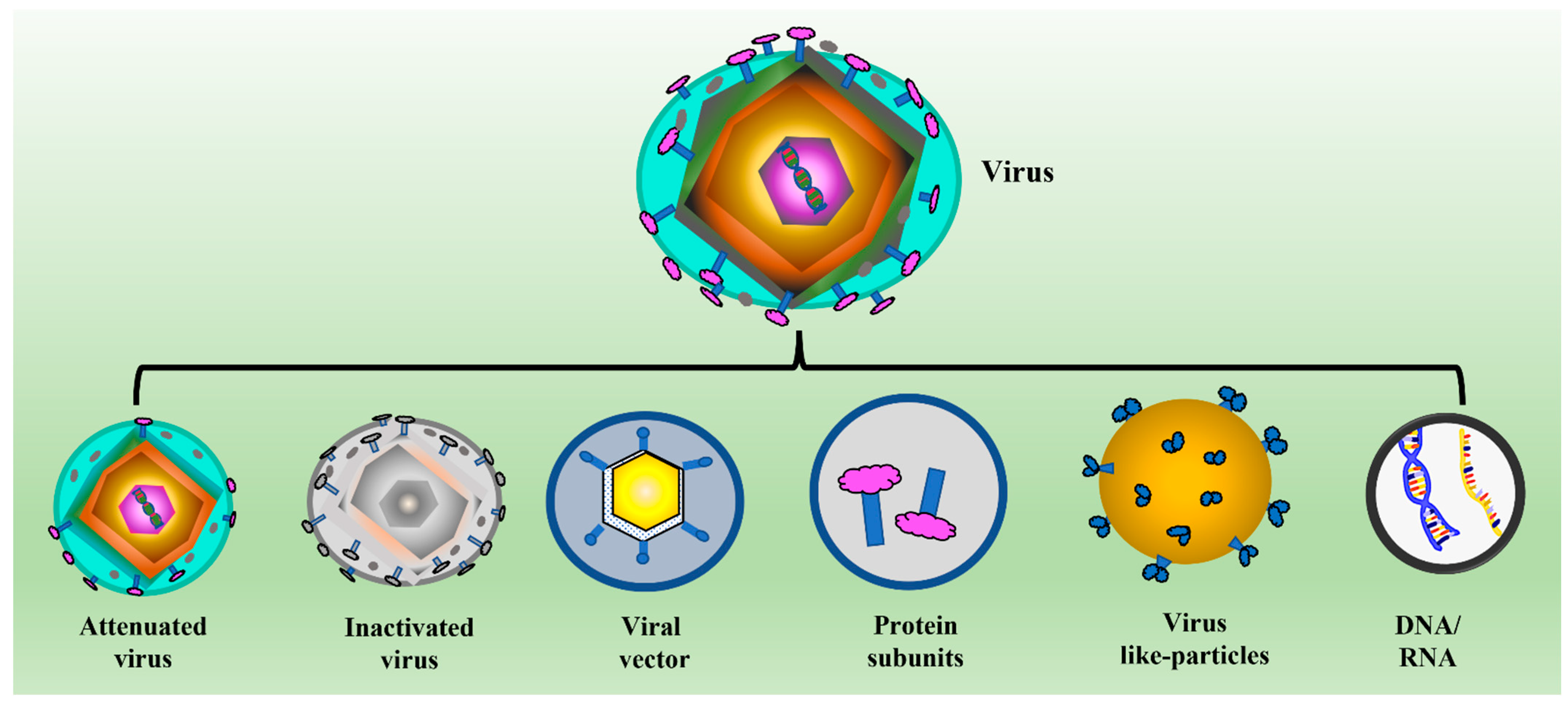
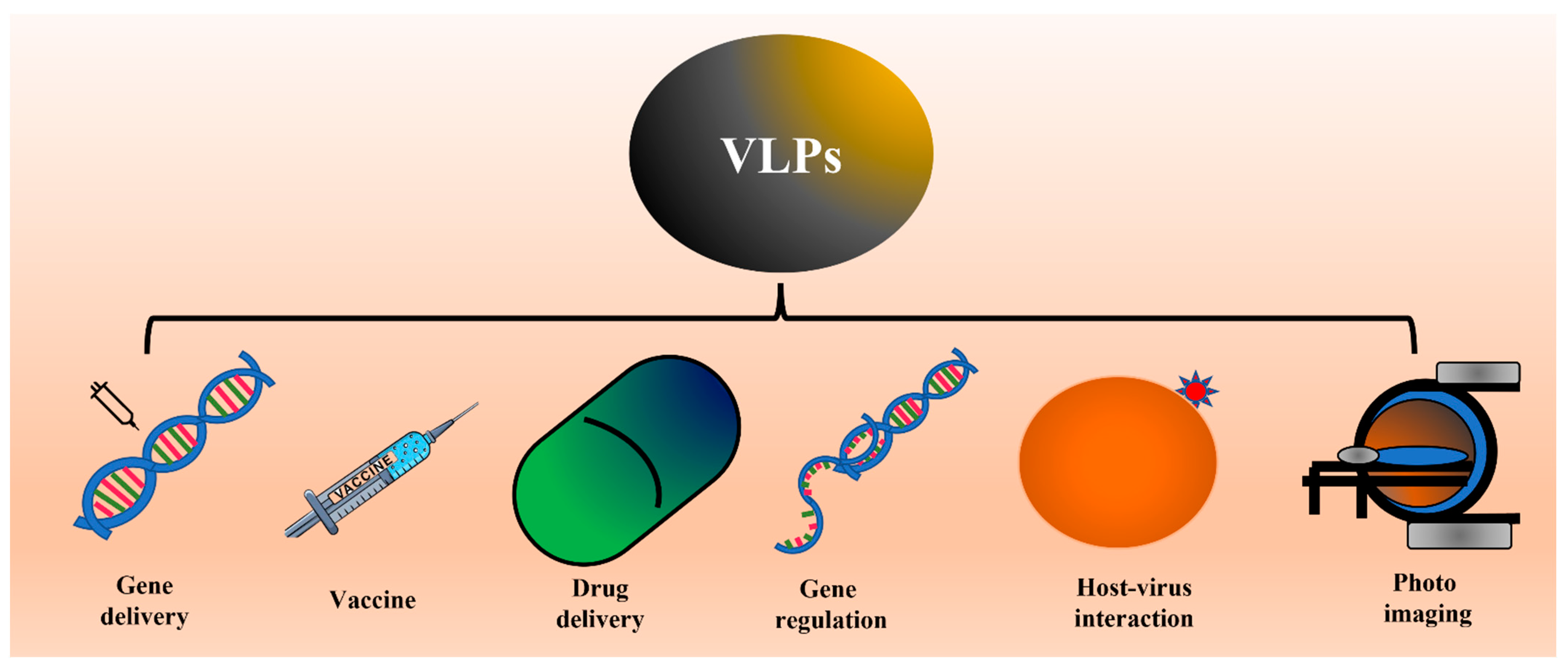
2. Introduction to Virus-Like Particles
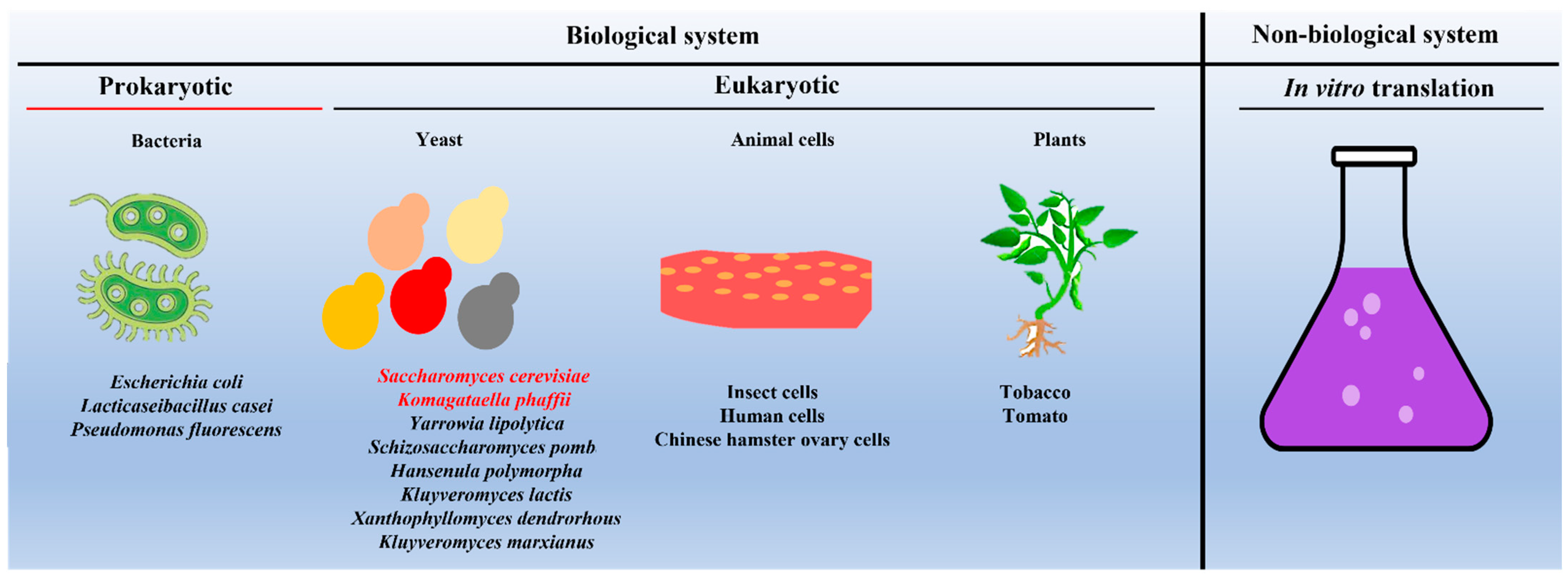
3. Yeast, Host to Produce VLPs on a Commercial Scale

{kind=link}
{kind=link}
{kind=link}
| S. No | Protein Antigen | Virus | Protein Localization | Promoter | References |
|---|---|---|---|---|---|
| 1 | Capsid protein | Red-spotted grouper nervous necrosis virus | IC | Pw42-2 | [123] |
| 2 | ZS and S | Zika virus | IC | AOX1 | [124] |
| 3 | 112-608aa of the ORF2 | Hepatitis E virus | EC | AOX1 | [125] |
| 4 | P1 and CD3 | Poliovirus type I | IC | AOX1 | [126,127] |
| 5 | Chimeric HPV-HIV L1P18 protein | HPV and HIV | IC | GAP | [128] |
| 6 | NY-ESO-1 cancer testis antigen | Norovirus | EC | AOX1 | [129] |
| 7 | Surface antigen | Hepatitis C virus | IC | AOX1 | [130,131] |
| 8 | P1 and 3CD | Enterovirus 71 | EC | AOX1 | [132,133] |
| 9 | E domain III | Dengue Virus | IC | AOX1 | [134,135] |
| 10 | P1 and 3CD | Coxsackievirus A16 | EC | GAP | [136] |
| 11 | VP1 | Norovirus | EC | AOX1 | [137] |
| 12 | E antigen | Dengue virus | IC | AOX1 | [138,139,140,141,142,143] |
| 13 | prME | Japanese encephalitis virus | EC | AOX1 | [144] |
| 14 | E antigen | Dengue virus | EC | GAP | [145] |
| 15 | Matrix protein | Nipah virus | IC | AOX1 | [146] |
| 16 | P1 and 3CD | Enterovirus D68 | IC | AOX1 | [147] |
| 17 | prM and E protein | Tick-borne encephalitis virus | EC | GAP | [148] |
| 18 | P1 and 3CD | Coxsackievirus A10 | IC | AOX1 | [149] |
| 19 | Surface antigen | Hepatitis B virus | IC | AOX1 | [150,151,152,153,154,155,156] |
| 20 | prM and E protein | Dengue virus | IC | GAP | [157] |
| 21 | L1 | HPV16 and 18 | IC | AOX1 | [158,159,160] |
| 22 | L1 | HPV 52 | AOX1 | [161] | |
| 23 | Capsid protein | Cowpea chlorotic mottle virus | EC | AOX1 | [162] |
| 24 | L1 | HPV 58 | IC | AOX1 | [163] |
| 25 | Envelope protein domain III (EDIII), hepatitis B surface antigen | Dengue virus | IC | AOX1 | [164] |
| 26 | VP2 | Infectious bursal disease virus | IC | AOX1 | [165] |
| 27 | P1 and 3CD | Coxsackievirus A16 | IC | AOX1 | [166] |
| 28 | Core protein | Hepatitis B virus | IC | AOX1 | [167,168] |
| 29 | E protein | Dengue virus | IC | AOX1 | [169] |
| 30 | L2 | Grapevine fanleaf virus | EC | AOX1 | [170] |
| 31 | Capsid protein (VP60) | Rabbit hemorrhagic disease virus | IC | AOX1 | [171] |
| 32 | HBc-influenza virus LAH domain | Hepatitis B/Influenza H3N2 virus | IC | AOX1 | [172] |
| 33 | CoreE1E2 Protein | Hepatitis C virus | EC | AOX1 | [173] |
| 34 | P1 and 3CD | Coxsackievirus A6 | IC | AOX1 | [174] |
| 35 | L1, L2 | HPV 16 | IC | AOX1 | [175] |
| 36 | prM/Env | Japanese encephalitis virus | IC | AOX1 | [176] |
| 37 | Den2E-HBsAg | Dengue/Hepatitis B virus | IC | AOX1 | [177] |
| 38 | prM and E protein | Dengue virus | IC | GAP | [178] |
| 39 | Polyprotein | Chikungunya virus | EC | AOX1 | [179] |
| 40 | L1 | HPV 16 | [180] | ||
| 41 | Major capsid protein | Iridovirus | EC | AOX1 | [181] |
| 42 | Surface antigen | Hepatitis B virus | [182] | ||
| 43 | VP1 | Rabbit hemorrhagic disease virus | IC | AOX1 | [183] |
| 44 | L1 | Bovine papillomavirus 1,2,4 | [184] | ||
| 45 | L1 | HPV 16 | IC | AOX1 | [185] |
| 46 | Capsid protein | Norovirus | IC | AOX1 | [186] |
| 47 | L1 | HPV 16 | EC | PGK1 | [187] |
| 48 | Core protein | Hepatitis B virus | [188] | ||
| 49 | VP1 | Calicivirus virus | EC | AOX1 | [189] |
| 50 | Core protein | Hepatitis C virus | IC | AOX1 | [190,191,192,193] |
4. Bottleneck in the Use of a Yeast-Based System on a Commercial Scale
5. In Vivo VLP Assembly and Secretion
6. Aggregation of VLPs, a Matter of Concern
7. Success Story
8. Conclusions and Future Direction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, S.; Guo, C. Rabies control and treatment: From prophylaxis to strategies with curative potential. Viruses 2016, 8, 279. [Google Scholar] [CrossRef] [PubMed]
- Kocourkova, A.; Honegr, J.; Kuca, K.; Danova, J. Vaccine Ingredients: Components that Influence Vaccine Efficacy. Mini-Rev. Med. Chem. 2017, 17, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Scollard, D.M.; Adams, L.B.; Gillis, T.P.; Krahenbuhl, J.L.; Truman, R.W.; Williams, D.L. The continuing challenges of leprosy. Clin. Microbiol. Rev. 2006, 19, 338–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, F.E. Malaria vaccines—Progress and problems. Trends Biotechnol. 1991, 9, 389–394. [Google Scholar] [CrossRef]
- Raffatellu, M.; Chessa, D.; Wilson, R.P.; Dusold, R.; Rubino, S.; Bäumler, A.J. The Vi capsular antigen of Salmonella enterica serotype Typhi reduces Toll-like receptor-dependent interleukin-8 expression in the intestinal mucosa. Infect. Immun. 2005, 73, 3367–3374. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A. Rubella vaccine. In Vaccines, 3rd ed.; Plotkin, S.A., Orenstein, W.A., Eds.; WB Saunders Co.: Philadelphia, PA, USA, 1998; pp. 409–439. [Google Scholar]
- Vogel, F.R.; Sarver, N. Nucleic acid vaccines. Clin. Microbiol. Rev. 1995, 8, 406–410. [Google Scholar] [CrossRef]
- Restifo, N.P.; Ying, H.; Hwang, L.; Leitner, W.W. The promise of nucleic acid vaccines. Gene Ther. 2000, 7, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [Green Version]
- Schlake, T.; Thess, A.; Fotin-Mleczek, M.; Kallen, K.J. Developing mRNA-vaccine technologies. RNA Biol. 2012, 9, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. mRNA-based therapeutics: Powerful and versatile tools to combat diseases. Signal Transduct. Target. Ther. 2022, 7, 166. [Google Scholar] [CrossRef]
- Kaur, S.P.; Gupta, V. COVID-19 Vaccine: A comprehensive status report. Virus Res. 2020, 288, 198114. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. The race for coronavirus vaccines: A graphical guide. Nature 2020, 580, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Srivastava, V.; Baindara, P.; Ahmad, A. Thermostable vaccines: An innovative concept in vaccine development. Expert Rev. Vaccines 2022, 21, 811–824. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin-Drubin, M.E.; Munger, K. Viruses associated with human cancer. Biochim. Biophys. Acta 2008, 1782, 127–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlom, J.; Hodge, J.W.; Palena, C.; Tsang, K.Y.; Jochems, C.; Greiner, J.W.; Farsaci, B.; Madan, R.A.; Heery, C.R.; Gulley, J.L. Therapeutic cancer vaccines. Adv. Cancer Res. 2014, 121, 67–124. [Google Scholar] [PubMed]
- Cuzick, J. Gardasil 9 joins the fight against cervix cancer. Expert Rev. Vaccines 2015, 14, 1047–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouyanfard, S.; Spagnoli, G.; Bulli, L.; Balz, K.; Yang, F.; Odenwald, C.; Seitz, H.; Mariz, F.C.; Bolchi, A.; Ottonello, S.; et al. Minor Capsid Protein L2 Polytope Induces Broad Protection against Oncogenic and Mucosal Human Papillomaviruses. J. Virol. 2018, 92, e01930-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolih, V.; Caldeira, J.; Bolli, E.; Salameh, A.; Conti, L.; Barutello, G.; Riccardo, F.; Magri, J.; Lamolinara, A.; Parra, K.; et al. Development of a VLP-Based Vaccine Displaying an xCT Extracellular Domain for the Treatment of Metastatic Breast Cancer. Cancers 2020, 12, 1492. [Google Scholar] [CrossRef]
- Zhang, S.; Yong, L.K.; Li, D.; Cubas, R.; Chen, C.; Yao, Q. Mesothelin virus-like particle immunization controls pancreatic cancer growth through CD8+ T cell induction and reduction in the frequency of CD4+foxp3+ICOS− regulatory T cells. PLoS ONE 2013, 8, e68303. [Google Scholar] [CrossRef]
- Cubas, R.; Zhang, S.; Li, M.; Chen, C.; Yao, Q. Chimeric Trop2 virus-like particles: A potential immunotherapeutic approach against pancreatic cancer. J. Immunother. 2011, 34, 251–263. [Google Scholar] [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Draper, S.J.; Sack, B.K.; King, C.R.; Nielsen, C.M.; Rayner, J.C.; Higgins, M.K.; Long, C.A.; Seder, R.A. Malaria Vaccines: Recent Advances and New Horizons. Cell Host Microbe 2018, 24, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Kharbikar, B.N. Lyophilized yeast powder for adjuvant free thermostable vaccine delivery. Appl. Microbiol. Biotechnol. 2021, 105, 3131–3143. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R. Investigating the long-term stability of protein immunogen(s) for whole recombinant yeast-based vaccines. FEMS Yeast Res. 2018, 18, foy071. [Google Scholar] [CrossRef] [Green Version]
- Bayer, M.E.; Blumberg, B.S.; Werner, B. Particles associated with Australia antigen in the sera of patients with leukaemia, Down’s Syndrome and Hepatitis. Nature 1968, 218, 1057–1059. [Google Scholar] [CrossRef]
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef]
- Mejía-Méndez, J.L.; Vazquez-Duhalt, R.; Hernández, L.R.; Sánchez-Arreola, E.; Bach, H. Virus-like Particles: Fundamentals and Biomedical Applications. Int. J. Mol. Sci. 2022, 23, 8579. [Google Scholar] [CrossRef]
- Lan, K.; Luo, M.H. Herpesviruses: Epidemiology, pathogenesis, and interventions. Virol. Sin. 2017, 32, 347–348. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Müller, K.M. In Vitro Assembly of Virus-Like Particles and Their Applications. Life 2021, 11, 334. [Google Scholar] [CrossRef]
- Jeong, H.; Seong, B.L. Exploiting virus-like particles as innovative vaccines against emerging viral infections. J. Microbiol. 2017, 55, 220–230. [Google Scholar] [CrossRef]
- Mortola, E.; Roy, P. Efficient assembly and release of SARS coronavirus-like particles by a heterologous expression system. FEBS Lett. 2004, 576, 174–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pushko, P.; Pumpens, P.; Grens, E. Development of virus-like particle technology from small highly symmetric to large complex virus-like particle structures. Intervirology 2013, 56, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Latham, T.; Galarza, J.M. Formation of wild-type and chimeric influenza virus-like particles following simultaneous expression of only four structural proteins. J. Virol. 2001, 75, 6154–6165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gedvilaite, A.; Kucinskaite-Kodze, I.; Lasickiene, R.; Timinskas, A.; Vaitiekaite, A.; Ziogiene, D.; Zvirbliene, A. Evaluation of Trichodysplasia Spinulosa-Associated Polyomavirus Capsid Protein as a New Carrier for Construction of Chimeric Virus-Like Particles Harboring Foreign Epitopes. Viruses 2015, 7, 4204–4229. [Google Scholar] [CrossRef] [Green Version]
- Bai, B.; Hu, Q.; Hu, H.; Zhou, P.; Shi, Z.; Meng, J.; Lu, B.; Huang, Y.; Mao, P.; Wang, H. Virus-like particles of SARS-like coronavirus formed by membrane proteins from different origins demonstrate stimulating activity in human dendritic cells. PLoS ONE 2008, 3, e2685. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.H.; Cai, H.; Steinmetz, N.F. Viral nanoparticles for drug delivery, imaging, immunotherapy, and theranostic applications. Adv. Drug Deliv. Rev. 2020, 156, 214–235. [Google Scholar] [CrossRef]
- Steinmetz, N.F. Viral nanoparticles as platforms for next-generation therapeutics and imaging devices. Nanomedicine 2010, 6, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, M.O.; Gomes, A.C.; Vogel, M.; Bachmann, M.F. Interaction of viral capsid-derived virus-like particles (VLPs) with the innate immune system. Vaccines 2018, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.L.; Peres, C.; Conniot, J.; Matos, A.I.; Moura, L.; Carreira, B.; Sainz, V.; Scomparin, A.; Satchi-Fainaro, R.; Préat, V.; et al. Nanoparticle impact on innate immune cell pattern-recognition receptors and inflammasomes activation. Semin. Immunol. 2017, 34, 3–24. [Google Scholar] [CrossRef]
- Lee, Y.T.; Ko, E.J.; Lee, Y.; Kim, K.H.; Kim, M.C.; Lee, Y.N.; Kang, S.M. Intranasal vaccination with M2e5x virus-like particles induces humoral and cellular immune responses conferring cross-protection against heterosubtypic influenza viruses. PLoS ONE 2018, 13, e0190868. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zheng, X.; Gai, W.; Wong, G.; Wang, H.; Jin, H.; Feng, N.; Zhao, Y.; Zhang, W.; Li, N.; et al. Novel chimeric virus-like particles vaccine displaying MERS-CoV receptor-binding domain induce specific humoral and cellular immune response in mice. Antiviral Res. 2017, 140, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.; Cheinsong-Popov, R.; Callow, D.; Adams, S.; Patou, G.; Hodgkin, K.; Martin, S.; Gotch, F.; Kingsman, A. Immunogenicity of the yeast recombinant p17p24: Ty virus-like particles (p24-VLP) in healthy volunteers. Vaccine 1995, 13, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Yong, C.Y.; Yeap, S.K.; Goh, Z.H.; Ho, K.L.; Omar, A.R.; Tan, W.S. Induction of humoral and cell-mediated immune responses by hepatitis B virus epitope displayed on the virus-like particles of prawn nodavirus. Appl. Environ. Microbiol. 2015, 81, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, L.; Lin, J.; Sun, Y.; Bennouna, S.; Lo, M.; Wu, Q.; Bu, Z.; Pulendran, B.; Compans, R.W.; Yang, C. Ebola virus-like particles produced in insect cells exhibit dendritic cell stimulating activity and induce neutralizing antibodies. Virology 2006, 351, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://www.gavi.org/vaccineswork/what-are-protein-subunit-vaccines-and-how-could-they-be-used-against-covid-19 (accessed on 10 November 2022).
- Sailaja, G.; Skountzou, I.; Quan, F.S.; Compans, R.W.; Kang, S.M. Human immunodeficiency virus-like particles activate multiple types of immune cells. Virology 2007, 362, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Radukic, M.T.; Mller, K.M. Adeno-associated virus capsid protein expression in Escherichia coli and chemically defined capsid assembly. Sci. Rep. 2019, 9, 18631. [Google Scholar] [CrossRef] [Green Version]
- Joe, C.C.; Chatterjee, S.; Lovrecz, G.; Adams, T.E.; Thaysen-Andersen, M.; Walsh, R.; Locarnini, S.A.; Smooker, P.; Netter, H.J. Glycoengineered hepatitis B virus-like particles with enhanced immunogenicity. Vaccine 2020, 38, 3892–3901. [Google Scholar] [CrossRef]
- Shiri, F.; Petersen, K.E.; Romanov, V.; Zou, Q.; Gale, B.K. Characterization and differential retention of Q beta bacteriophage virus-like particles using cyclical electrical field–flow fractionation and asymmetrical flow fieldflow fractionation. Anal. Bioanal. Chem. 2020, 412, 1563–1572. [Google Scholar] [CrossRef]
- Zhai, L.; Yadav, R.; Kunda, N.K.; Anderson, D.; Bruckner, E.; Miller, E.K.; Basu, R.; Muttil, P.; Tumban, E. Oral immunization with bacteriophage MS2-L2 VLPs protects against oral and genital infection with multiple HPV types associated with head & neck cancers and cervical cancer. Antiviral Res. 2019, 166, 56–65. [Google Scholar]
- Glass, P.J.; White, L.J.; Ball, J.M.; Leparc-Goffart, I.; Hardy, M.E.; Estes, M.K. Norwalk virus open reading frame 3 encodes a minor structural protein. J. Virol. 2000, 74, 6581–6591. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, B.; Al-Barwani, F.; Young, V.; Scullion, S.; Ward, V.; Young, S. Virus-Like Particles, a Versatile Subunit Vaccine Platform. In Subunit Vaccine Delivery; Part of the Advances in Delivery Science and Technology Book Series; Springer: New York, NY, USA, 2014; pp. 159–180. [Google Scholar]
- Kim, H.J.; Kim, H.J. Yeast as an expression system for producing virus-like particles: What factors do we need to consider? Lett. Appl. Microbiol. 2017, 64, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kumar, P. Yeast-based vaccines: New perspective in vaccine development and application. FEMS Yeast Res. 2019, 19, foz007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhães, P.O.; Lopes, A.M.; Mazzola, P.G.; Rangel-Yagui, C.; Penna, T.C.; Pessoa, A., Jr. Methods of endotoxin removal from biological preparations: A review. J. Pharm. Pharm. Sci. 2007, 10, 388–404. [Google Scholar] [PubMed]
- Baneyx, F.; Mujacic, M. Recombinant protein folding and misfolding in Escherichia coli. Nat. Biotechnol. 2004, 22, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Dalton, A.C.; Barton, W.A. Over-expression of secreted proteins from mammalian cell lines. Protein Sci. 2014, 23, 517–525. [Google Scholar] [CrossRef]
- Bill, R.M. Recombinant protein subunit vaccine synthesis in microbes: A role for yeast? J. Pharm. Pharmacol. 2015, 67, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Desai, P.N.; Shrivastava, N.; Padh, H. Production of heterologous proteins in plants: Strategies for optimal expression. Biotechnol. Adv. 2010, 28, 427–435. [Google Scholar] [CrossRef]
- Lazutka, J.; Simutis, K.; Matulis, P.; Petraitytė-Burneikienė, R.; Kučinskaitė-Kodzė, I.; Simanavičius, M.; Tamošiunas, P.L. Antigenicity study of the yeast-generated human parvovirus 4 (PARV4) virus-like particles. Virus Res. 2021, 292, 198236. [Google Scholar] [CrossRef]
- Simanavicius, M.; Tamosiunas, P.L.; Petraityte-Burneikiene, R.; Johne, R.; Ulrich, R.G.; Zvirbliene, A.; Kucinskaite-Kodze, I. Generation in yeast and antigenic characterization of hepatitis E virus capsid protein virus-like particles. Appl. Microbiol. Biotechnol. 2018, 102, 185–198. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, L.; Chang, N.; Shi, P.; Gao, T.; Zhang, L.; Huang, J. Preparation of virus-like particles for porcine circovirus type 2 by Yeast Fab Assembly. Virus Genes 2018, 54, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Juozapaitis, M.; Slibinskas, R.; Staniulis, J.; Sakaguchi, T.; Sasnauskas, K. Generation of Sendai virus nucleocapsid-like particles in yeast. Virus Res. 2005, 108, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Tamošiūnas, P.L.; Petraitytė-Burneikienė, R.; Bulavaitė, A.; Marcinkevičiūtė, K.; Simutis, K.; Lasickienė, R.; Firantienė, R.; Ėmužytė, R.; Žvirblienė, A.; Sasnauskas, K. Yeast-generated virus-like particles as antigens for detection of human bocavirus 1-4 specific antibodies in human serum. Appl. Microbiol. Biotechnol. 2016, 100, 4935–4946. [Google Scholar] [CrossRef] [PubMed]
- Burden, C.S.; Jin, J.; Podgornik, A.; Bracewell, D.G. A monolith purification process for virus-like particles from yeast homogenate. J. Chromatogr. B 2012, 880, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kee, G.S.; Jin, J.; Balasundaram, B.; Bracewell, D.G.; Pujar, N.S.; Titchener-Hooker, N.J. Exploiting the intracellular compartmentalization characteristics of the S. cerevisiae host cell for enhancing primary purification of lipid-envelope virus-like particles. Biotechnol. Prog. 2010, 26, 26–33. [Google Scholar] [PubMed]
- Kee, G.S.; Pujar, N.S.; Titchener-Hooker, N.J. Study of detergent-mediated liberation of hepatitis B virus-like particles from S. cerevisiae homogenate: Identifying a framework for the design of future-generation lipoprotein vaccine processes. Biotechnol. Prog. 2008, 24, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Tsunetsugu-Yokota, Y.; Morikawa, Y.; Isogai, M.; Kawana-Tachikawa, A.; Odawara, T.; Nakamura, T.; Grassi, F.; Autran, B.; Iwamoto, A. Yeast-derived human immunodeficiency virus type 1 p55(gag) virus-like particles activate dendritic cells (DCs) and induce perforin expression in Gag-specific CD8+ T cells by cross-presentation of DCs. J. Virol. 2003, 77, 10250–10259. [Google Scholar] [CrossRef] [Green Version]
- Norkiene, M.; Stonyte, J.; Ziogiene, D.; Mazeike, E.; Sasnauskas, K.; Gedvilaite, A. Production of recombinant VP1-derived virus-like particles from novel human polyomaviruses in yeast. BMC Biotechnol. 2015, 15, 68. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Cho, S.Y.; Park, M.H.; Kim, H.J. Comparison of the size distributions and immunogenicity of human papillomavirus type 16 L1 virus-like particles produced in insect and yeast cells. Arch. Pharm. Res. 2018, 41, 544–553. [Google Scholar] [CrossRef]
- Kim, H.J.; Jin, Y.; Kim, H.J. The concentration of carbon source in the medium affects the quality of virus-like particles of human papillomavirus type 16 produced in Saccharomyces cerevisiae. PLoS ONE 2014, 9, e94467. [Google Scholar] [CrossRef] [Green Version]
- Petraityte, R.; Tamosiunas, P.L.; Juozapaitis, M.; Zvirbliene, A.; Sasnauskas, K.; Shiell, B.; Russell, G.; Bingham, J.; Michalski, W.P. Generation of Tioman virus nucleocapsid-like particles in yeast Saccharomyces cerevisiae. Virus Res. 2009, 145, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.L.; Chen, P.J.; Mu, J.J.; Chi, W.K.; Kao, T.L.; Hwang, L.H.; Chen, D.S. Assembly of hepatitis delta virus-like empty particles in yeast. Virology 1997, 236, 374–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nainys, J.; Lasickiene, R.; Petraityte-Burneikiene, R.; Dabrisius, J.; Lelesius, R.; Sereika, V.; Zvirbliene, A.; Sasnauskas, K.; Gedvilaite, A. Generation in yeast of recombinant virus-like particles of porcine circovirus type 2 capsid protein and their use for a serologic assay and development of monoclonal antibodies. BMC Biotechnol. 2014, 14, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Xiao, X.; Zhao, M.; Liu, W.; Pang, L.; Sun, X.; Cen, S.; Yang, B.B.; Huang, Y.; Sheng, W.; et al. EV71 virus-like particles produced by co-expression of capsid proteins in yeast cells elicit humoral protective response against EV71 lethal challenge. BMC Res. Notes 2016, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasnauskas, K.; Bulavaite, A.; Hale, A.; Jin, L.; Knowles, W.A.; Gedvilaite, A.; Dargeviciūte, A.; Bartkeviciūte, D.; Zvirbliene, A.; Staniulis, J.; et al. Generation of recombinant virus-like particles of human and non-human polyomaviruses in yeast Saccharomyces cerevisiae. Intervirology 2002, 45, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Backovic, A.; Cervelli, T.; Salvetti, A.; Zentilin, L.; Giacca, M.; Galli, A. Capsid protein expression and adeno-associated virus like particles assembly in Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulavaitė, A.; Lasickienė, R.; Tamošiūnas, P.L.; Simanavičius, M.; Sasnauskas, K.; Žvirblienė, A. Synthesis of human parainfluenza virus 4 nucleocapsid-like particles in yeast and their use for detection of virus-specific antibodies in human serum. Appl. Microbiol. Biotechnol. 2017, 101, 2991–3004. [Google Scholar] [CrossRef] [PubMed]
- Freivalds, J.; Dislers, A.; Ose, V.; Skrastina, D.; Cielens, I.; Pumpens, P.; Sasnauskas, K.; Kazaks, A. Assembly of bacteriophage Qbeta virus-like particles in yeast Saccharomyces cerevisiae and Pichia pastoris. J. Biotechnol. 2006, 123, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Wi, G.R.; Hwang, J.Y.; Kwon, M.G.; Kim, H.J.; Kang, H.A.; Kim, H.J. Protective immunity against nervous necrosis virus in convict grouper Epinephelus septemfasciatus following vaccination with virus-like particles produced in yeast Saccharomyces cerevisiae. Vet. Microbiol. 2015, 177, 214–218. [Google Scholar] [CrossRef]
- Penkert, R.R.; Young, N.S.; Surman, S.L.; Sealy, R.E.; Rosch, J.; Dormitzer, P.R.; Settembre, E.C.; Chandramouli, S.; Wong, S.; Hankins, J.S.; et al. Saccharomyces cerevisiae-derived virus-like particle parvovirus B19 vaccine elicits binding and neutralizing antibodies in a mouse model for sickle cell disease. Vaccine 2017, 35, 3615–3620. [Google Scholar] [CrossRef]
- Chandramouli, S.; Medina-Selby, A.; Coit, D.; Schaefer, M.; Spencer, T.; Brito, L.A.; Zhang, P.; Otten, G.; Mandl, C.W.; Mason, P.W.; et al. Generation of a parvovirus B19 vaccine candidate. Vaccine 2013, 31, 3872–3878. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, A.; Gedvilaite, A.; Reetz, J.; Rösler, U.; Müller, H.; Johne, R. Serological cross-reactions between four polyomaviruses of birds using virus-like particles expressed in yeast. J. Gen. Virol. 2012, 93, 2658–2667. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, H.Y.; Han, J.F.; Deng, Y.Q.; Zhu, S.Y.; Li, X.F.; Yang, H.Q.; Li, Y.X.; Zhang, Y.; Qin, E.D.; et al. Novel recombinant chimeric virus-like particle is immunogenic and protective against both enterovirus 71 and coxsackievirus A16 in mice. Sci. Rep. 2015, 5, 7878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucarey, S.A.; Noriega, J.; Reyes, P.; Tapia, C.; Sáenz, L.; Zuñiga, A.; Tobar, J.A. The optimized capsid gene of porcine circovirus type 2 expressed in yeast forms virus-like particles and elicits antibody responses in mice fed with recombinant yeast extracts. Vaccine 2009, 27, 5781–5790. [Google Scholar] [CrossRef]
- Rodríguez-Limas, W.A.; Tyo, K.E.; Nielsen, J.; Ramírez, O.T.; Palomares, L.A. Molecular and process design for rotavirus-like particle production in Saccharomyces cerevisiae. Microb. Cell Fact. 2011, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Limas, W.A.; Pastor, A.R.; Esquivel-Soto, E.; Esquivel-Guadarrama, F.; Ramírez, O.T.; Palomares, L.A. Immunogenicity and protective efficacy of yeast extracts containing rotavirus-like particles: A potential veterinary vaccine. Vaccine 2014, 32, 2794–2798. [Google Scholar] [CrossRef]
- Tamošiūnas, P.L.; Simutis, K.; Kodzė, I.; Firantienė, R.; Emužytė, R.; Petraitytė-Burneikienė, R.; Zvirblienė, A.; Sasnauskas, K. Production of human parvovirus 4 VP2 virus-like particles in yeast and their evaluation as an antigen for detection of virus-specific antibodies in human serum. Intervirology 2013, 56, 271–277. [Google Scholar] [CrossRef]
- Bulavaitė, A.; Lasickienė, R.; Vaitiekaitė, A.; Sasnauskas, K.; Žvirblienė, A. Synthesis of human parainfluenza virus 2 nucleocapsid protein in yeast as nucleocapsid-like particles and investigation of its antigenic structure. Appl. Microbiol. Biotechnol. 2016, 100, 4523–4534. [Google Scholar] [CrossRef]
- Juozapaitis, M.; Serva, A.; Kucinskaite, I.; Zvirbliene, A.; Slibinskas, R.; Staniulis, J.; Sasnauskas, K.; Shiell, B.J.; Bowden, T.R.; Michalski, W.P. Generation of menangle virus nucleocapsid-like particles in yeast Saccharomyces cerevisiae. J. Biotechnol. 2007, 130, 441–447. [Google Scholar] [CrossRef]
- Tomo, N.; Goto, T.; Morikawa, Y. Trans-packaging of human immunodeficiency virus type 1 genome into Gag virus-like particles in Saccharomyces cerevisiae. Microb. Cell Fact. 2013, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Tamošiūnas, P.L.; Petraitytė-Burneikienė, R.; Lasickienė, R.; Akatov, A.; Kundrotas, G.; Sereika, V.; Lelešius, R.; Žvirblienė, A.; Sasnauskas, K. Generation of recombinant porcine parvovirus virus-like particles in Saccharomyces cerevisiae and development of virus-specific monoclonal antibodies. J. Immunol. Res. 2014, 2014, 573531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Li, H.Y.; Han, J.F.; Deng, Y.Q.; Li, Y.X.; Zhu, S.Y.; He, Y.L.; Qin, E.D.; Chen, R.; Qin, C.F. Virus-like particles produced in Saccharomyces cerevisiae elicit protective immunity against Coxsackievirus A16 in mice. Appl. Microbiol. Biotechnol. 2013, 97, 10445–10452. [Google Scholar] [CrossRef] [PubMed]
- Zaveckas, M.; Snipaitis, S.; Pesliakas, H.; Nainys, J.; Gedvilaite, A. Purification of recombinant virus-like particles of porcine circovirus type 2 capsid protein using ion-exchange monolith chromatography. J. Chromatogr. B 2015, 991, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Pleckaityte, M.; Bremer, C.M.; Gedvilaite, A.; Kucinskaite-Kodze, I.; Glebe, D.; Zvirbliene, A. Construction of polyomavirus-derived pseudotype virus-like particles displaying a functionally active neutralizing antibody against hepatitis B virus surface antigen. BMC Biotechnol. 2015, 15, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasnauskas, K.; Buzaite, O.; Vogel, F.; Jandrig, B.; Razanskas, R.; Staniulis, J.; Scherneck, S.; Krüger, D.H.; Ulrich, R. Yeast cells allow high-level expression and formation of polyomavirus-like particles. Biol. Chem. 1999, 380, 381–386. [Google Scholar] [CrossRef]
- Jansen, K.U.; Rosolowsky, M.; Schultz, L.D.; Markus, H.Z.; Cook, J.C.; Donnelly, J.J.; Martinez, D.; Ellis, R.W.; Shaw, A.R. Vaccination with yeast-expressed cottontail rabbit papillomavirus (CRPV) virus-like particles protects rabbits from CRPV-induced papilloma formation. Vaccine 1995, 13, 1509–1514. [Google Scholar] [CrossRef]
- Lowe, R.S.; Brown, D.R.; Bryan, J.T.; Cook, J.C.; George, H.A.; Hofmann, K.J.; Hurni, W.M.; Joyce, J.G.; Lehman, E.D.; Markus, H.Z.; et al. Human papillomavirus type 11 (HPV-11) neutralizing antibodies in the serum and genital mucosal secretions of African green monkeys immunized with HPV-11 virus-like particles expressed in yeast. J. Infect. Dis. 1997, 176, 1141–1145. [Google Scholar] [CrossRef]
- Neeper, M.P.; Hofmann, K.J.; Jansen, K.U. Expression of the major capsid protein of human papillomavirus type 11 in Saccharomyces cerevisae. Gene 1996, 180, 1–6. [Google Scholar] [CrossRef]
- Cook, J.C.; Joyce, J.G.; George, H.A.; Schultz, L.D.; Hurni, W.M.; Jansen, K.U.; Hepler, R.W.; Ip, C.; Lowe, R.S.; Keller, P.M.; et al. Purification of virus-like particles of recombinant human papillomavirus type 11 major capsid protein L1 from Saccharomyces cerevisiae. Protein Expr. Purif. 1999, 17, 477–484. [Google Scholar] [CrossRef]
- Jagadish, M.N.; Ward, C.W.; Gough, K.H.; Tulloch, P.A.; Whittaker, L.A.; Shukla, D.D. Expression of potyvirus coat protein in Escherichia coli and yeast and its assembly into virus-like particles. J. Gen. Virol. 1991, 72, 1543–1550. [Google Scholar] [CrossRef]
- Wang, X.W.; Sheng, W.; Zeng, Y. Formation and identification of virus-like particles of poliovirus type I. Chin. J. Exp. Clin. Virol. 2013, 27, 373–375. (In Chinese) [Google Scholar]
- Karpenko, L.I.; Lebedev, L.R.; Ignatyev, G.M.; Agafonov, A.P.; Poryvaeva, V.A.; Pronyaeva, T.R.; Ryabchikova, E.I.; Pokrovsky, A.G.; Ilyichev, A.A. Construction of artificial virus-like particles exposing HIV epitopes, and the study of their immunogenic properties. Vaccine 2003, 21, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Thorsteinsson, M.V.; Johnston, L.B.; DePhillips, P.A.; Zlotnick, A. A quantitative description of in vitro assembly of human papillomavirus 16 virus-like particles. J. Mol. Biol. 2008, 381, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Mach, H.; Volkin, D.B.; Troutman, R.D.; Wang, B.; Luo, Z.; Jansen, K.U.; Shi, L. Disassembly and reassembly of yeast-derived recombinant human papillomavirus virus-like particles (HPV VLPs). J. Pharm. Sci. 2006, 95, 2195–2206. [Google Scholar] [CrossRef]
- Gedvilaite, A.; Zvirbliene, A.; Staniulis, J.; Sasnauskas, K.; Krüger, D.H.; Ulrich, R. Segments of puumala hantavirus nucleocapsid protein inserted into chimeric polyomavirus-derived virus-like particles induce a strong immune response in mice. Viral Immunol. 2004, 17, 51–68. [Google Scholar] [CrossRef]
- Hadiji-Abbes, N.; Martin, M.; Benzina, W.; Karray-Hakim, H.; Gergely, C.; Gargouri, A.; Mokdad-Gargouri, R. Extraction and purification of hepatitis B virus-like M particles from a recombinant Saccharomyces cerevisiae strain using alumina powder. J. Virol. Methods 2013, 187, 132–137. [Google Scholar] [CrossRef]
- Zielonka, A.; Gedvilaite, A.; Ulrich, R.; Lüschow, D.; Sasnauskas, K.; Müller, H.; Johne, R. Generation of virus-like particles consisting of the major capsid protein VP1 of goose hemorrhagic polyomavirus and their application in serological tests. Virus Res. 2006, 120, 128–137. [Google Scholar] [CrossRef]
- Lawatscheck, R.; Aleksaite, E.; Schenk, J.A.; Micheel, B.; Jandrig, B.; Holland, G.; Sasnauskas, K.; Gedvilaite, A.; Ulrich, R.G. Chimeric polyomavirus-derived virus-like particles: The immunogenicity of an inserted peptide applied without adjuvant to mice depends on its insertion site and its flanking linker sequence. Viral Immunol. 2007, 20, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Pumpens, P.; Razanskas, R.; Pushko, P.; Renhof, R.; Gusars, I.; Skrastina, D.; Ose, V.; Borisova, G.; Sominskaya, I.; Petrovskis, I.; et al. Evaluation of HBs, HBc, and frCP virus-like particles for expression of human papillomavirus 16 E7 oncoprotein epitopes. Intervirology 2002, 45, 24–32. [Google Scholar] [CrossRef]
- Choi, Y.R.; Kim, H.J.; Lee, J.Y.; Kang, H.A.; Kim, H.J. Chromatographically purified capsid proteins of red-spotted grouper nervous necrosis virus expressed in Saccharomyces cerevisiae form virus-like particles. Protein Expr. Purif. 2013, 89, 162–168. [Google Scholar] [CrossRef]
- Kwag, H.L.; Kim, H.J.; Chang, D.Y.; Kim, H.J. The production and immunogenicity of human papillomavirus type 58 virus-like particles produced in Saccharomyces cerevisiae. J. Microbiol. 2012, 50, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Hadiji-Abbes, N.; Mihoubi, W.; Martin, M.; Karakasyan-Dia, C.; Frikha, F.; Gergely, C.; Jouenne, T.; Gargouri, A.; Mokdad-Gargouri, R. Characterization of C69R variant HBsAg: Effect on binding to anti-HBs and the structure of virus-like particles. Arch. Virol. 2015, 160, 2427–2433. [Google Scholar] [CrossRef] [PubMed]
- Stolt, A.; Sasnauskas, K.; Koskela, P.; Lehtinen, M.; Dillner, J. Seroepidemiology of the human polyomaviruses. J. Gen. Virol. 2003, 84, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Lowin, T.; Raab, U.; Schroeder, J.; Franssila, R.; Modrow, S. Parvovirus B19 VP2-proteins produced in Saccharomyces cerevisiae: Comparison with VP2-particles produced by baculovirus-derived vectors. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 348–352. [Google Scholar] [CrossRef]
- Joyce, J.G.; Tung, J.S.; Przysiecki, C.T.; Cook, J.C.; Lehman, E.D.; Sands, J.A.; Jansen, K.U.; Keller, P.M. The L1 major capsid protein of human papillomavirus type 11 recombinant virus-like particles interacts with heparin and cell-surface glycosaminoglycans on human keratinocytes. J. Biol. Chem. 1999, 274, 5810–5822. [Google Scholar] [CrossRef] [Green Version]
- Gedvilaite, A.; Frömmel, C.; Sasnauskas, K.; Micheel, B.; Ozel, M.; Behrsing, O.; Staniulis, J.; Jandrig, B.; Scherneck, S.; Ulrich, R. Formation of immunogenic virus-like particles by inserting epitopes into surface-exposed regions of hamster polyomavirus major capsid protein. Virology 2000, 273, 21–35. [Google Scholar] [CrossRef]
- Fu, J.; VanDusen, W.J.; Kolodin, D.G.; O’Keefe, D.O.; Herber, W.K.; George, H.A. Continuous culture study of the expression of hepatitis B surface antigen and its self-assembly into virus-like particles in Saccharomyces cerevisiae. Biotechnol. Bioeng. 1996, 49, 578–586. [Google Scholar] [CrossRef]
- Towne, V.; Zhao, Q.; Brown, M.; Finnefrock, A.C. Pairwise antibody footprinting using surface plasmon resonance technology to characterize human papillomavirus type 16 virus-like particles with direct anti-HPV antibody immobilization. J. Immunol. Methods 2013, 388, 1–7. [Google Scholar] [CrossRef]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [Green Version]
- Barsøe, S.; Toffan, A.; Pascoli, F.; Stratmann, A.; Pretto, T.; Marsella, A.; Er-Rafik, M.; Vendramin, N.; Olesen, N.J.; Sepúlveda, D.; et al. Long-Term Protection and Serologic Response of European Sea Bass Vaccinated with a Betanodavirus Virus-Like Particle Produced in Pichia pastoris. Vaccines 2021, 9, 447. [Google Scholar] [CrossRef]
- Shanmugam, R.K.; Ramasamy, V.; Shukla, R.; Arora, U.; Swaminathan, S.; Khanna, N. Pichia pastoris-expressed Zika virus envelope domain III on a virus-like particle platform: Design, production and immunological evaluation. Pathog. Dis. 2019, 77, ftz026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, J.; Kaul, S.; Srivastava, A.; Kaushik, N.; Ghosh, S.; Sharma, C.; Batra, G.; Banerjee, M.; Shalimar Nayak, B.; Ranjith-Kumar, C.T.; et al. Expression, Purification and Characterization of the Hepatitis E Virus Like-Particles in the Pichia pastoris. Front. Microbiol. 2020, 11, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherry, L.; Grehan, K.; Snowden, J.S.; Knight, M.L.; Adeyemi, O.O.; Rowlands, D.J.; Stonehouse, N.J. Comparative Molecular Biology Approaches for the Production of Poliovirus Virus-Like Particles Using Pichia pastoris. mSphere 2020, 5, e00838-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherry, L.; Grehan, K.; Swanson, J.J.; Bahar, M.W.; Porta, C.; Fry, E.E.; Stuart, D.I.; Rowlands, D.J.; Stonehouse, N.J. Production and Characterisation of Stabilised PV-3 Virus-like Particles Using Pichia pastoris. Viruses 2022, 14, 2159. [Google Scholar] [CrossRef] [PubMed]
- Eto, Y.; Saubi, N.; Ferrer, P.; Joseph-Munné, J. Expression of Chimeric HPV-HIV Protein L1P18 in Pichia pastoris; Purification and Characterization of the Virus-like Particles. Pharmaceutics 2021, 13, 1967. [Google Scholar] [CrossRef]
- Parker, S.A.; Maloy, M.H.; Tome-Amat, J.; Bardliving, C.L.; Batt, C.A.; Lanz, K.J.; Olesberg, J.T.; Arnold, M.A. Optimization of norovirus virus-like particle production in Pichia pastoris using a real-time near-infrared bioprocess monitor. Biotechnol. Prog. 2016, 32, 518–526. [Google Scholar] [CrossRef]
- Pechelyulko, A.; Andreeva-Kovalevskaya, Z.; Dmitriev, D.; Lavrov, V.; Massino, Y.; Nagel, A.; Segal, O.; Sokolova, O.S.; Solonin, A.; Tarakanova, Y.; et al. A simple method to purify recombinant HCV core protein expressed in Pichia pastoris for obtaining virus-like particles and producing monoclonal antibodies. Protein Expr. Purif. 2021, 183, 105864. [Google Scholar] [CrossRef]
- Acosta-Rivero, N.; Aguilar, J.C.; Musacchio, A.; Falcón, V.; Viña, A.; de la Rosa, M.C.; Morales, J. Characterization of the HCV core virus-like particles produced in the methylotrophic yeast Pichia pastoris. Biochem. Biophys. Res. Commun. 2001, 287, 122–125. [Google Scholar] [CrossRef]
- Yang, Z.; Gao, F.; Wang, X.; Shi, L.; Zhou, Z.; Jiang, Y.; Ma, X.; Zhang, C.; Zhou, C.; Zeng, X.; et al. Development and characterization of an enterovirus 71 (EV71) virus-like particles (VLPs) vaccine produced in Pichia pastoris. Hum. Vaccin. Immunother. 2020, 16, 1602–1610. [Google Scholar] [CrossRef]
- Zhang, C.; Ku, Z.; Liu, Q.; Wang, X.; Chen, T.; Ye, X.; Li, D.; Jin, X.; Huang, Z. High-yield production of recombinant virus-like particles of enterovirus 71 in Pichia pastoris and their protective efficacy against oral viral challenge in mice. Vaccine 2015, 33, 2335–2341. [Google Scholar] [CrossRef]
- Shukla, R.; Rajpoot, R.K.; Arora, U.; Poddar, A.; Swaminathan, S.; Khanna, N. Pichia pastoris-Expressed Bivalent Virus-Like Particulate Vaccine Induces Domain III-Focused Bivalent Neutralizing Antibodies without Antibody-Dependent Enhancement in Vivo. Front. Microbiol. 2018, 8, 2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, U.; Tyagi, P.; Swaminathan, S.; Khanna, N. Virus-like particles displaying envelope domain III of dengue virus type 2 induce virus-specific antibody response in mice. Vaccine 2013, 31, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; He, Y.; Lu, J. Virus-Like Particles Produced in Pichia Pastoris Induce Protective Immune Responses against Coxsackievirus A16 in Mice. Med. Sci. Monit. 2016, 22, 3370–3382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomé-Amat, J.; Fleischer, L.; Parker, S.A.; Bardliving, C.L.; Batt, C.A. Secreted production of assembled Norovirus virus-like particles from Pichia pastoris. Microb. Cell Fact. 2014, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poddar, A.; Ramasamy, V.; Shukla, R.; Rajpoot, R.K.; Arora, U.; Jain, S.K.; Swaminathan, S.; Khanna, N. Virus-like particles derived from Pichia pastoris-expressed dengue virus type 1 glycoprotein elicit homotypic virus-neutralizing envelope domain III-directed antibodies. BMC Biotechnol. 2016, 16, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, L.; Mani, S.; Raut, R.; Poddar, A.; Tyagi, P.; Arora, U.; de Silva, A.; Swaminathan, S.; Khanna, N. Pichia pastoris-expressed dengue 3 envelope-based virus-like particles elicit predominantly domain III-focused high titer neutralizing antibodies. Front. Microbiol. 2015, 6, 1005. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.H.; Fu, J.L.; Sugrue, R.J. Characterization of the dengue virus envelope glycoprotein expressed in Pichia pastoris. Methods Mol. Biol. 2007, 379, 163–176. [Google Scholar]
- Mani, S.; Tripathi, L.; Raut, R.; Tyagi, P.; Arora, U.; Barman, T.; Sood, R.; Galav, A.; Wahala, W.; de Silva, A.; et al. Pichia pastoris-expressed dengue 2 envelope forms virus-like particles without pre-membrane protein and induces high titer neutralizing antibodies. PLoS ONE 2013, 8, e64595. [Google Scholar] [CrossRef]
- Rajpoot, R.K.; Shukla, R.; Arora, U.; Swaminathan, S.; Khanna, N. Dengue envelope-based ‘four-in-one’ virus-like particles produced using Pichia pastoris induce enhancement-lacking, domain III-directed tetravalent neutralising antibodies in mice. Sci. Rep. 2018, 8, 8643. [Google Scholar] [CrossRef] [Green Version]
- Khetarpal, N.; Shukla, R.; Rajpoot, R.K.; Poddar, A.; Pal, M.; Swaminathan, S.; Arora, U.; Khanna, N. Recombinant Dengue Virus 4 Envelope Glycoprotein Virus-Like Particles Derived from Pichia pastoris are Capable of Eliciting Homotypic Domain III-Directed Neutralizing Antibodies. Am. J. Trop. Med. Hyg. 2017, 96, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Jiang, Y.; Wang, J.; Fan, H.; Cao, R. Secreted expression of Japanese encephalitis virus prME in Pichia pastoris and immunogenicity evaluation of the virus-like particles in mice. Chin. J. Biotechnol. 2017, 33, 863–874. (In Chinese) [Google Scholar]
- Liu, W.; Jiang, H.; Zhou, J.; Yang, X.; Tang, Y.; Fang, D.; Jiang, L. Recombinant dengue virus-like particles from Pichia pastoris: Efficient production and immunological properties. Virus Genes 2010, 40, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Joseph, N.M.; Ho, K.L.; Tey, B.T.; Tan, C.S.; Shafee, N.; Tan, W.S. Production of the virus-like particles of nipah virus matrix protein in Pichia pastoris as diagnostic reagents. Biotechnol. Prog. 2016, 32, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, X.; Zhang, W.; Dai, W.; Xie, J.; Ye, L.; Wang, H.; Chen, H.; Liu, Q.; Gong, S.; et al. Enterovirus D68 virus-like particles expressed in Pichia pastoris potently induce neutralizing antibody responses and confer protection against lethal viral infection in mice. Emerg. Microbes Infect. 2018, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.M.; Jeong, Y.E.; Wang, E.; Lee, Y.J.; Han, M.G.; Park, C.; Lee, W.J.; Choi, W. Cloning and Expression of Recombinant Tick-Borne Encephalitis Virus-like Particles in Pichia pastoris. Osong Public Health Res. Perspect. 2014, 5, 274–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhang, C.; Liu, Q.; Gong, S.; Geng, L.; Huang, Z. A virus-like particle vaccine protects mice against coxsackievirus A10 lethal infection. Antiviral Res. 2018, 152, 124–130. [Google Scholar] [CrossRef]
- Zahid, M.; Rinas, U. Guidelines for Small-Scale Production and Purification of Hepatitis B Surface Antigen Virus-Like Particles from Recombinant Pichia pastoris. Methods Mol. Biol. 2019, 1923, 309–322. [Google Scholar]
- Lünsdorf, H.; Gurramkonda, C.; Adnan, A.; Khanna, N.; Rinas, U. Virus-like particle production with yeast: Ultrastructural and immunocytochemical insights into Pichia pastoris producing high levels of the hepatitis B surface antigen. Microb. Cell Fact. 2011, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.N.; Sarvari, T.; Bashiri, G.; Khatami, M.; Shojaosadati, S.A. Assessing virus like particles formation and r-HBsAg aggregation during large scale production of recombinant hepatitis B surface antigen from Pichia pastoris. Int. J. Biol. Macromol. 2019, 139, 697–711. [Google Scholar] [CrossRef]
- Gurramkonda, C.; Zahid, M.; Nemani, S.K.; Adnan, A.; Gudi, S.K.; Khanna, N.; Ebensen, T.; Lünsdorf, H.; Guzmán, C.A.; Rinas, U. Purification of hepatitis B surface antigen virus-like particles from recombinant Pichia pastoris and in vivo analysis of their immunogenic properties. J. Chromatogr. B 2013, 940, 104–111. [Google Scholar] [CrossRef]
- Liu, R.; Lin, Q.; Sun, Y.; Lu, X.; Qiu, Y.; Li, Y.; Guo, X. Expression, purification, and characterization of hepatitis B virus surface antigens (HBsAg) in yeast Pichia Pastoris. Appl. Biochem. Biotechnol. 2009, 158, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Gurramkonda, C.; Adnan, A.; Gäbel, T.; Lünsdorf, H.; Ross, A.; Nemani, S.K.; Swaminathan, S.; Khanna, N.; Rinas, U. Simple high-cell density fed-batch technique for high-level recombinant protein production with Pichia pastoris: Application to intracellular production of Hepatitis B surface antigen. Microb. Cell Fact. 2009, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watelet, B.; Quibriac, M.; Rolland, D.; Gervasi, G.; Gauthier, M.; Jolivet, M.; Letourneur, O. Characterization and diagnostic potential of hepatitis B virus nucleocapsid expressed in E. coli and P. pastoris. J. Virol. Methods 2002, 99, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.X.; Jiang, L.F.; Zhou, J.M.; Yin, Y.; Yang, X.M.; Liu, W.Q.; Fang, D.Y. Induction of virus-neutralizing antibodies and T cell responses by dengue virus type 1 virus-like particles prepared from Pichia pastoris. Chin. Med. J. 2012, 125, 1986–1992. [Google Scholar] [PubMed]
- Gupta, G.; Glueck, R.; Rishi, N. Physicochemical characterization and immunological properties of Pichia pastoris based HPV16L1 and 18L1 virus like particles. Biologicals 2017, 46, 11–22. [Google Scholar] [CrossRef]
- Hanumantha, R.N.; Baji, B.P.; Rajendra, L.; Sriraman, R.; Pang, Y.Y.; Schiller, J.T.; Srinivasan, V.A. Expression of codon optimized major capsid protein (L1) of human papillomavirus type 16 and 18 in Pichia pastoris; purification and characterization of the virus-like particles. Vaccine 2011, 29, 7326–7334. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.; Giannino, V.; Rishi, N.; Glueck, R. Immunogenicity of next-generation HPV vaccines in non-human primates: Measles-vectored HPV vaccine versus Pichia pastoris recombinant protein vaccine. Vaccine 2016, 34, 4724–4731. [Google Scholar] [CrossRef]
- Dewi, K.S.; Chairunnisa, S.; Swasthikawati, S.; Yuliawati Agustiyanti, D.F.; Mustopa, A.Z.; Kusharyoto, W.; Ningrum, R.A. Production of codon-optimized Human papillomavirus type 52 L1 virus-like particles in Pichia pastoris BG10 expression system. Prep. Biochem. Biotechnol. 2022, 53, 148–156. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, K.; Liu, A.; Lu, X.; Yang, L.; Zhao, Q. Highly secretory expression of recombinant cowpea chlorotic mottle virus capsid proteins in Pichia pastoris and in-vitro encapsulation of ruthenium nanoparticles for catalysis. Protein Expr. Purif. 2020, 174, 105679. [Google Scholar] [CrossRef]
- Jiang, Z.; Tong, G.; Cai, B.; Xu, Y.; Lou, J. Purification and immunogenicity study of human papillomavirus 58 virus-like particles expressed in Pichia pastoris. Protein Expr. Purif. 2011, 80, 203–210. [Google Scholar] [CrossRef]
- Ramasamy, V.; Arora, U.; Shukla, R.; Poddar, A.; Shanmugam, R.K.; White, L.J.; Mattocks, M.M.; Raut, R.; Perween, A.; Tyagi, P.; et al. A tetravalent virus-like particle vaccine designed to display domain III of dengue envelope proteins induces multi-serotype neutralizing antibodies in mice and macaques which confer protection against antibody dependent enhancement in AG129 mice. PLoS Negl. Trop. Dis. 2018, 12, e0006191. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Pan, Q.; Lu, Z.; Li, K.; Gao, H.; Qi, X.; Gao, Y.; Wang, X. An optimized, highly efficient, self-assembled, subvirus-like particle of infectious bursal disease virus (IBDV). Vaccine 2016, 34, 3508–3514. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, Q.; Ku, Z.; Hu, Y.; Ye, X.; Zhang, Y.; Huang, Z. Coxsackievirus A16-like particles produced in Pichia pastoris elicit high-titer neutralizing antibodies and confer protection against lethal viral challenge in mice. Antiviral Res. 2016, 129, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Stephen, S.L.; Beales, L.; Peyret, H.; Roe, A.; Stonehouse, N.J.; Rowlands, D.J. Recombinant Expression of Tandem-HBc Virus-Like Particles (VLPs). Methods Mol. Biol. 2018, 1776, 97–123. [Google Scholar] [PubMed]
- Freivalds, J.; Dislers, A.; Ose, V.; Pumpens, P.; Tars, K.; Kazaks, A. Highly efficient production of phosphorylated hepatitis B core particles in yeast Pichia pastoris. Protein Expr. Purif. 2011, 75, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Sugrue, R.J.; Fu, J.; Howe, J.; Chan, Y.C. Expression of the dengue virus structural proteins in Pichia pastoris leads to the generation of virus-like particles. J. Gen. Virol. 1997, 78, 1861–1866. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, R.; Shams-Bakhsh, M.; Hassani-Mehraban, A.; Arab, S.S.; Thelen, N.; Thiry, M.; Crommen, J.; Fillet, M.; Jacobs, N.; Brans, A.; et al. Production and characterization of virus-like particles of grapevine fanleaf virus presenting L2 epitope of human papillomavirus minor capsid protein. BMC Biotechnol. 2019, 19, 81. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.W.; Cui, Z.Z.; Wang, Y.K. Expression of capsid gene of Chinese isolate of rabbit hemorrhagic disease virus in Pichia pastoris. Chin. J. Biotechnol. 2005, 21, 135–138. (In Chinese) [Google Scholar]
- Kazaks, A.; Lu, I.N.; Farinelle, S.; Ramirez, A.; Crescente, V.; Blaha, B.; Ogonah, O.; Mukhopadhyay, T.; de Obanos, M.P.; Krimer, A.; et al. Production and purification of chimeric HBc virus-like particles carrying influenza virus LAH domain as vaccine candidates. BMC Biotechnol. 2017, 17, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazlalipour, M.; Keyvani, H.; Monavari, S.H.; Mollaie, H.R. Expression, Purification and Immunogenic Description of a Hepatitis C Virus Recombinant CoreE1E2 Protein Expressed by Yeast Pichia pastoris. Jundishapur J. Microbiol. 2015, 8, e17157. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Shen, C.; Zhang, C.; Zhang, W.; Wang, L.; Lan, K.; Liu, Q.; Huang, Z. Yeast-produced recombinant virus-like particles of coxsackievirus A6 elicited protective antibodies in mice. Antiviral Res. 2016, 132, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Sanchooli, A.; Aghaiypour, K.; Kiasari, B.A.; Samarbaf-Zadeh, A.; Ghadiri, A.; Makvandi, M. VLP Production from Recombinant L1/L2 HPV-16 Protein Expressed in Pichia pastoris. Protein Pept. Lett. 2018, 25, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xiao, A.; Wang, H.; Zhang, X.; Zhang, Y.; Li, Y.; Wei, Y.; Liu, W.; Chen, C. A VLP-Based Vaccine Candidate Protects Mice against Japanese Encephalitis Virus Infection. Vaccines 2022, 10, 197. [Google Scholar] [CrossRef] [PubMed]
- Bisht, H.; Chugh, D.A.; Raje, M.; Swaminathan, S.S.; Khanna, N. Recombinant dengue virus type 2 envelope/hepatitis B surface antigen hybrid protein expressed in Pichia pastoris can function as a bivalent immunogen. J. Biotechnol. 2002, 99, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, J.; Yu, Z.; Fang, D.; Fu, C.; Zhu, X.; He, Z.; Yan, H.; Jiang, L. Tetravalent recombinant dengue virus-like particles as potential vaccine candidates: Immunological properties. BMC Microbiol. 2014, 14, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraswat, S.; Athmaram, T.N.; Parida, M.; Agarwal, A.; Saha, A.; Dash, P.K. Expression and Characterization of Yeast Derived Chikungunya Virus Like Particles (CHIK-VLPs) and Its Evaluation as a Potential Vaccine Candidate. PLoS Negl. Trop. Dis. 2016, 10, e0004782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coimbra, E.C.; Gomes, F.B.; Campos, J.F.; D’arc, M.; Carvalho, J.C.; Mariz, F.C.; Jesus, A.L.; Stocco, R.C.; Beçak, W.; Freitas, A.C. Production of L1 protein from different types of HPV in Pichia pastoris using an integrative vector. Braz. J. Med. Biol. Res. 2011, 44, 1209–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Fan, Y.; LaPatra, S.E.; Ma, J.; Xu, J.; Meng, Y.; Jiang, N.; Zeng, L. Protective immunity of a Pichia pastoris expressed recombinant iridovirus major capsid protein in the Chinese giant salamander, Andrias davidianus. Vaccine 2015, 33, 5662–5669. [Google Scholar] [CrossRef]
- Hosseini, S.N.; Javidanbardan, A.; Alizadeh Salim, B.S.; Khatami, M. Large-scale purification of recombinant hepatitis B surface antigen from Pichia pastoris with non-affinity chromatographic methods as a substitute to immunoaffinity chromatography. Prep. Biochem. Biotechnol. 2018, 48, 683–692. [Google Scholar] [CrossRef]
- Fernández, E.; Toledo, J.R.; Méndez, L.; González, N.; Parra, F.; Martín-Alonso, J.M.; Limonta, M.; Sánchez, K.; Cabrales, A.; Estrada, M.P.; et al. Conformational and thermal stability improvements for the large-scale production of yeast-derived rabbit hemorrhagic disease virus-like particles as multipurpose vaccine. PLoS ONE 2013, 8, e56417. [Google Scholar] [CrossRef] [Green Version]
- Jesus, A.L.; Mariz, F.C.; Souza, H.M.; Cordeiro, M.N.; Coimbra, E.C.; Leitão, M.C.; Nascimento, L.M.; Stocco, R.C.; Beçak, W.; Freitas, A.C. Expression of the bovine papillomavirus type 1, 2 and 4 L1 genes in the yeast Pichia pastoris. Genet. Mol. Res. 2012, 11, 2598–2607. [Google Scholar] [CrossRef] [PubMed]
- Bazan, S.B.; de Alencar Muniz Chaves, A.; Aires, K.A.; Cianciarullo, A.M.; Garcea, R.L.; Ho, P.L. Expression and characterization of HPV-16 L1 capsid protein in Pichia pastoris. Arch. Virol. 2009, 154, 1609–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, M.; Farkas, T.; Jiang, X. Norovirus capsid protein expressed in yeast forms virus-like particles and stimulates systemic and mucosal immunity in mice following an oral administration of raw yeast extracts. J. Med. Virol. 2007, 79, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Mariz, F.C.; Coimbra, E.C.; Jesus, A.L.; Nascimento, L.M.; Torres, F.A.; Freitas, A.C. Development of an IP-Free Biotechnology Platform for Constitutive Production of HPV16 L1 Capsid Protein Using the Pichia pastoris PGK1 Promoter. BioMed Res. Int. 2015, 2015, 594120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, D.; Gauthier, M.; Dugua, J.M.; Fournier, C.; Delpech, L.; Watelet, B.; Letourneur, O.; Arnaud, M.; Jolivet, M. Purification of recombinant HBc antigen expressed in Escherichia coli and Pichia pastoris: Comparison of size-exclusion chromatography and ultracentrifugation. J. Chromatogr. B Biomed. Sci. Appl. 2001, 753, 51–65. [Google Scholar] [CrossRef]
- Fernández, E.; Toledo, J.R.; Mansur, M.; Sánchez, O.; Gil, D.F.; González-González, Y.; Lamazares, E.; Fernández, Y.; Parra, F.; Farnós, O. Secretion and assembly of calicivirus-like particles in high-cell-density yeast fermentations: Strategies based on a recombinant non-specific BPTI-Kunitz-type protease inhibitor. Appl. Microbiol. Biotechnol. 2015, 99, 3875–3886. [Google Scholar] [CrossRef]
- Acosta-Rivero, N.; Rodriguez, A.; Musacchio, A.; Falcón, V.; Suarez, V.M.; Martinez, G.; Guerra, I.; Paz-Lago, D.; Morera, Y.; de la Rosa, M.C.; et al. In vitro assembly into virus-like particles is an intrinsic quality of Pichia pastoris derived HCV core protein. Biochem. Biophys. Res. Commun. 2004, 325, 68–74. [Google Scholar] [CrossRef]
- Acosta-Rivero, N.; Falcón, V.; Alvarez, C.; Musacchio, A.; Chinea, G.; Cristina de la Rosa, M.; Rodriguez, A.; Dueñas-Carrera, S.; Tsutsumi, V.; Shibayama, M.; et al. Structured HCV nucleocapsids composed of P21 core protein assemble primary in the nucleus of Pichia pastoris yeast. Biochem. Biophys. Res. Commun. 2003, 310, 48–53. [Google Scholar] [CrossRef]
- Acosta-Rivero, N.; Alvarez-Obregón, J.C.; Musacchio, A.; Falcón, V.; Dueñas-Carrera, S.; Marante, J.; Menéndez, I.; Morales, J. In vitro self-assembled HCV core virus-like particles induce a strong antibody immune response in sheep. Biochem. Biophys. Res. Commun. 2002, 290, 300–304. [Google Scholar] [CrossRef]
- Majeau, N.; Gagné, V.; Bolduc, M.; Leclerc, D. Signal peptide peptidase promotes the formation of hepatitis C virus non-enveloped particles and is captured on the viral membrane during assembly. J. Gen. Virol. 2005, 86, 3055–3064. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, P.; Du, J.; Li, X.; Lu, W.; Hao, X.; Dong, B.; Yu, Y.; Wang, L. High-level expression and immunogenicity of porcine circovirus type 2b capsid protein without nuclear localization signal expressed in Hansenula polymorpha. Biologicals 2018, 51, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, D.; Barbian, A.; Jenzelewski, V.; Schembecker, G.; Merz, J.; Piontek, M. Bioprocess optimization for purification of chimeric VLP displaying BVDV E2 antigens produced in yeast Hansenula polymorpha. J. Biotechnol. 2019, 306, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, D.; Rolf, T.; Suckow, M.; Kranz, A.; Barbian, A.; Chan, J.A.; Leitsch, J.; Weniger, M.; Jenzelewski, V.; Kouskousis, B.; et al. Establishment of a yeast based VLP platform for antigen presentation. Microb. Cell Fact. 2018, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Li, L.; Jin, Z.; Han, X.; Zhao, P.; Wang, L.; Jiang, C.; Wang, Y.; Wang, W.; Xu, D.; et al. Fermentation, purification and immunogenicity evaluation of hepatitis E virus-like particles expressed in Hansenula polymorpha. Chin. J. Biotechnol. 2017, 33, 653–663. (In Chinese) [Google Scholar]
- Chan, J.A.; Wetzel, D.; Reiling, L.; Miura, K.; Drew, D.R.; Gilson, P.R.; Anderson, D.A.; Richards, J.S.; Long, C.A.; Suckow, M.; et al. Malaria vaccine candidates displayed on novel virus-like particles are immunogenic and induce transmission-blocking activity. PLoS ONE 2019, 14, e0221733. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, D.; Chan, J.A.; Suckow, M.; Barbian, A.; Weniger, M.; Jenzelewski, V.; Reiling, L.; Richards, J.S.; Anderson, D.A.; Kouskousis, B.; et al. Display of malaria transmission-blocking antigens on chimeric duck hepatitis B virus-derived virus-like particles produced in Hansenula polymorpha. PLoS ONE 2019, 14, e0221394. [Google Scholar] [CrossRef] [Green Version]
- McGregor, J.; Hardy, J.M.; Lay, C.S.; Boo, I.; Piontek, M.; Suckow, M.; Coulibaly, F.; Poumbourios, P.; Center, R.J.; Drummer, H.E. Virus-Like Particles Containing the E2 Core Domain of Hepatitis C Virus Generate Broadly Neutralizing Antibodies in Guinea Pigs. J. Virol. 2022, 96, e0167521. [Google Scholar] [CrossRef]
- Duan, J.; Yang, D.; Chen, L.; Yu, Y.; Zhou, J.; Lu, H. Efficient production of porcine circovirus virus-like particles using the nonconventional yeast Kluyveromyces marxianus. Appl. Microbiol. Biotechnol. 2019, 103, 833–842. [Google Scholar] [CrossRef]
- Simon, C.; Klose, T.; Herbst, S.; Han, B.G.; Sinz, A.; Glaeser, R.M.; Stubbs, M.T.; Lilie, H. Disulfide linkage and structure of highly stable yeast-derived virus-like particles of murine polyomavirus. J. Biol. Chem. 2014, 289, 10411–10418. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Chen, L.; Duan, J.; Yu, Y.; Zhou, J.; Lu, H. Investigation of Kluyveromyces marxianus as a novel host for large-scale production of porcine parvovirus virus-like particles. Microb. Cell Fact. 2021, 20, 24. [Google Scholar] [CrossRef]
- Luu, V.T.; Moon, H.Y.; Hwang, J.Y.; Kang, B.K.; Kang, H.A. Development of recombinant Yarrowia lipolytica producing virus-like particles of a fish nervous necrosis virus. J. Microbiol. 2017, 55, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Flores, O.; Alcaíno, J.; Fernandez-Lobato, M.; Cifuentes, V.; Baeza, M. Characterization of virus-like particles and identification of capsid proteins in Xanthophyllomyces dendrorhous. Virus Genes 2015, 50, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Kadri, A.; Wege, C.; Jeske, H. In vivo self-assembly of TMV-like particles in yeast and bacteria for nanotechnological applications. J. Virol. Methods 2013, 189, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Wildt, S.; Gerngross, T.U. The humanization of N-glycosylation pathways in yeast. Nat. Rev. Microbiol. 2005, 3, 119–128. [Google Scholar] [CrossRef]
- Conde, R.; Cueva, R.; Pablo, G.; Polaina, J.; Larriba, G. A search for hyperglycosylation signals in yeast glycoproteins. J. Biol. Chem. 2004, 279, 43789–43798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, W.; Thompson, E.; Badger, C.; Mellquist, J.L.; Garrison, A.R.; Smith, J.M.; Paragas, J.; Hogan, R.J.; Schmaljohn, C. Influences of glycosylation on antigenicity, immunogenicity, and protective efficacy of Ebola virus GP DNA vaccines. J. Virol. 2007, 81, 1821–1837. [Google Scholar] [CrossRef] [Green Version]
- Sakuragi, S.; Goto, T.; Sano, K.; Morikawa, Y. HIV type 1 Gag virus-like particle budding from spheroplasts of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 7956–7961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervera, L.; Gutiérrez-Granados, S.; Martínez, M.; Blanco, J.; Gòdia, F.; Segura, M.M. Generation of HIV-1 Gag VLPs by transient transfection of HEK 293 suspension cell cultures using an optimized animal-derived component free medium. J. Biotechnol. 2013, 166, 152–165. [Google Scholar] [CrossRef]
- Fuenmayor, J.; Gòdia, F.; Cervera, L. Production of virus-like particles for vaccines. N. Biotechnol. 2017, 39, 174–180. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Quan, C.; Luo, J.; Yang, Y.; Yu, M.; Kong, Y.; Ma, G.; Su, Z. Aggregation and antigenicity of virus like particle in salt solution–A case study with hepatitis B surface antigen. Vaccine 2015, 33, 4300–4306. [Google Scholar] [CrossRef]
- Lang, R.; Winter, G.; Vogt, L.; Zurcher, A.; Dorigo, B.; Schimmele, B. Rational design of a stable, freeze-dried virus-like particle-based vaccine formulation. Drug Dev. Ind. Pharm. 2009, 35, 83–97. [Google Scholar] [CrossRef]
- Jezek, J.; Chen, D.; Watson, L.; Crawford, J.; Perkins, S.; Tyagi, A.; Jones-Braun, L. A heat-stable hepatitis B vaccine formulation. Hum. Vaccines 2009, 5, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Kissmann, J.; Ausar, S.F.; Foubert, T.R.; Brock, J.; Switzer, M.H.; Detzi, E.J.; Vedvick, T.S.; Middaugh, C.R. Physical stabilization of Norwalk virus-like particles. J. Pharm. Sci. 2008, 97, 4208–4218. [Google Scholar] [CrossRef] [PubMed]
- Tsoka, S.; Holwill, I.; Hoare, M. Virus-like particle analysis in yeast homogenate using a laser light-scattering assay. Biotechnol. Bioeng. 1999, 63, 290–297. [Google Scholar] [CrossRef]
- Shi, L.; Sanyal, G.; Ni, A.; Luo, Z.; Doshna, S.; Wang, B.; Graham, T.L.; Wang, N.; Volkin, D.B. Stabilization of human papillomavirus virus-like particles by non-ionic surfactants. J. Pharm. Sci. 2005, 94, 1538–1551. [Google Scholar] [CrossRef]
- Ding, Y.; Chuan, Y.P.; He, L.; Middelberg, A.P. Modeling the competition between aggregation and self-assembly during virus-like particle processing. Biotechnol. Bioeng. 2010, 107, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Bi, J.; Zhou, W.; Li, Y.; Wang, Y.; Ma, G.; Su, Z. Improving recovery of recombinant hepatitis B virus surface antigen by ion-exchange chromatographic supports with low ligand density. Process Biochem. 2006, 41, 2320–2326. [Google Scholar] [CrossRef]
- Meingast, C.; Heldt, C.L. Arginine-enveloped virus inactivation and potential mechanisms. Biotechnol. Prog. 2020, 36, e2931. [Google Scholar] [CrossRef]
- Kissmann, J.; Joshi, S.B.; Haynes, J.R.; Dokken, L.; Richardson, C.; Middaugh, C.R. H1N1 influenza virus-like particles: Physical degradation pathways and identification of stabilizers. J. Pharm. Sci. 2011, 100, 634–645. [Google Scholar] [CrossRef]
- Hashemi, K.; Ghahramani Seno, M.M.; Ahmadian, M.R.; Malaekeh-Nikouei, B.; Bassami, M.R.; Dehghani, H.; Afkhami-Goli, A. Optimizing the synthesis and purification of MS2 virus like particles. Sci. Rep. 2021, 11, 19851. [Google Scholar] [CrossRef]
- Galula, J.U.; Chang, G.J.; Chao, D.Y. Production and Purification of Dengue Virus-like Particles from COS-1 Cells. Bio. Protoc. 2019, 9, e3280. [Google Scholar] [CrossRef] [PubMed]
- Pease, L.F., 3rd; Lipin, D.I.; Tsai, D.H.; Zachariah, M.R.; Lua, L.H.; Tarlov, M.J.; Middelberg, A.P. Quantitative characterization of virus-like particles by asymmetrical flow field flow fractionation, electrospray differential mobility analysis, and transmission electron microscopy. Biotechnol Bioeng. 2009, 102, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Lua, L.H.L.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P.J. Bioengineering Virus-like Particles as Vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Modis, Y.; High, K.; Towne, V.; Meng, Y.; Wang, Y.; Alexandroff, J.; Brown, M.; Carragher, B.; Potter, C.S.; et al. Disassembly and reassembly of human papillomavirus virus-like particles produces more virion-like antibody reactivity. Virol. J. 2012, 9, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, Y.Y. RTS, S/AS01 malaria vaccine (Mosquirix®): A profile of its use. Drugs Ther. Perspect. 2022, 38, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Ashley, C.E.; Carnes, E.C.; Phillips, G.K.; Durfee, P.N.; Buley, M.D.; Lino, C.A.; Padilla, D.P.; Phillips, B.; Carter, M.B.; Willman, C.L.; et al. Cell-specific delivery of diverse cargos by bacteriophage MS2 virus-like particles. ACS Nano 2011, 5, 5729–5745. [Google Scholar] [CrossRef] [Green Version]
- Yacoby, I.; Shamis, M.; Bar, H.; Shabat, D.; Benhar, I. Targeting antibacterial agents by using drug-carrying filamentous bacteriophages. Antimicrob. Agents Chemother. 2006, 50, 2087–2097. [Google Scholar] [CrossRef] [Green Version]
- Crooke, S.N.; Schimer, J.; Raji, I.; Wu, B.; Oyelere, A.K.; Finn, M.G. Lung Tissue Delivery of Virus-Like Particles Mediated by Macrolide Antibiotics. Mol. Pharm. 2019, 16, 2947–2955. [Google Scholar] [CrossRef]
- Legendre, D.; Fastrez, J. Production in Saccharomyces cerevisiae of MS2 virus-like particles packaging functional heterologous mRNAs. J. Biotechnol. 2005, 117, 183–194. [Google Scholar] [CrossRef]
- Sun, S.; Li, W.; Sun, Y.; Pan, Y.; Li, J. A new RNA vaccine platform based on MS2 virus-like particles produced in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2011, 407, 124–128. [Google Scholar] [CrossRef]
- Spice, A.J.; Aw, R.; Bracewell, D.G.; Polizzi, K.M. Synthesis and Assembly of Hepatitis B Virus-Like Particles in a Pichia pastoris Cell-Free System. Front. Bioeng. Biotechnol. 2020, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornesello, A.L.; Tagliamonte, M.; Buonaguro, F.M.; Tornesello, M.L.; Buonaguro, L. Virus-like Particles as Preventive and Therapeutic Cancer Vaccines. Vaccines 2022, 10, 227. [Google Scholar] [CrossRef] [PubMed]
- Fei, W.; Shui, G.; Zhang, Y.; Krahmer, N.; Ferguson, C.; Kapterian, T.S.; Lin, R.C.; Dawes, I.W.; Brown, A.J.; Li, P.; et al. A role for phosphatidic acid in the formation of “supersized” lipid droplets. PLoS Genet. 2011, 7, e1002201. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Feature | Bacteria | Insect Cells | Mammalian Cells | Plant | Yeast |
|---|---|---|---|---|---|
| Production cost | Low | High | High | Moderate | Low |
| Growth media | Simple | Complex | Complex | Simple | Simple |
| Growth | Fast | Slow | Slow | Very slow | Fast |
| Growth duration | Very small | Small | Small | Long | Small |
| Indoor/Outdoor | Indoor | Indoor | Indoor | Outdoor/polyhouse | Indoor |
| Scale-up | Easy | Very difficult | Very difficult | Difficult | Easy |
| Secretion | No | Yes | Yes | NA | Yes |
| Enveloped/non-enveloped | Non-enveloped | Enveloped/Non-enveloped | Enveloped/Non-enveloped | NA | Non-enveloped/Enveloped possible |
| Speed of transformant screening | Very fast | Slow | Slow | Very slow | Fast |
| Effect of seasonal variations | No | No | No | Yes | No |
| S. No | Protein Antigen | Virus | Protein Localization | Promoter | References |
|---|---|---|---|---|---|
| 1 | VP2 | Human parvovirus 4 | IC | Hybrid GAL10-PYK1promoter | [62] |
| 2 | Capsid protein | Hepatitis E virus | IC | GAL promoter | [63] |
| 3 | Capsid protein | Porcine circovirus type 2 | EC | GPD, TEF2 | [64] |
| 4 | Nucleocapsid protein | Sendai virus | IC | GAL7 | [65] |
| 5 | VP2, VP1 | Human bocaviruses | IC | [66] | |
| 6 | Surface antigen | Hepatitis B virus | IC | GAL | [67,68,69] |
| 7 | p55(gag) | HIV-1 | EC | [70] | |
| 8 | VP1 | Human polyomaviruses | IC | GAL | [71] |
| 9 | L1 | HPV 16 | IC | GAL10 | [72,73] |
| 10 | Nucleocapsid protein | Tioman virus | IC | GAL10 | [74] |
| 11 | L-HDAg and surface antigen | Hepatitis delta virus | IC | GAD | [75] |
| 12 | Capsid protein | Porcine circovirus type 2 | IC | GAL10 | [76] |
| 13 | Capsid protein | Enterovirus 71 | IC | GAL10 | [77] |
| 14 | VP1 | Human and non-human polyomaviruses | IC | GAL | [78] |
| 15 | Capsid protein | Adeno-associated virus | IC | GAL1 | [79] |
| 16 | Nucleocapsid protein | Human parainfluenza virus 4 | IC | GAL7 | [80] |
| 17 * | Coat protein | Cacteriophage Qbeta virus | IC | GAL | [81] |
| 18 | Capsid protein | Nervous necrosis virus | IC | GAL | [82] |
| 19 | VP1,2 | Parvovirus B19 | IC | ADH2/GAPDH | [83,84] |
| 20 | VP1 | Bird polyomaviruses | IC | GAL | [85] |
| 21 | P1 | Enterovirus 71 and Coxsackievirus A16 | IC | GAL1 | [86] |
| 22 | Capsid protein | Porcine circovirus type 2 | IC | GAL1 | [87] |
| 23 | VP2,6,7 | Rotavirus | IC | PGK1, TEF1 | [88,89] |
| 24 | VP2 | Human parvovirus 4 | IC | GAL1-10 | [90] |
| 25 | Nucleocapsid protein | Human parainfluenza virus 2 | IC | GAL | [91] |
| 26 | Nucleocapsid protein | Menangle virus | IC | GAL7 | [92] |
| 27 | Gag | HIV-1 | IC | GAP | [93] |
| 28 | VP2 | Porcine parvovirus | IC | GAL1-10 | [94] |
| 29 | P1, CD3 | Coxsackievirus A16 | IC | GAL1 | [95] |
| 30 | Capsid protein | Porcine circovirus type 2 | IC | GAL | [96] |
| 31 | VP1,2 | Hepatitis B/Polyomavirus | IC | GAL7 | [97] |
| 32 | VP1 | Hamster polyomavirus | IC | [98] | |
| 33 | L1/L1 + L2 | Cottontail rabbit papillomavirus | IC | GAL1-10 | [99] |
| 34 | L1 | HPV 11 | IC | GAL | [100,101,102] |
| 35 | Coat protein | Potyvirus (Johnsongrass mosaic virus) | IC | ADC1 | [103] |
| 36 | P1, CD3 | Poliovirus type I | IC | [104] | |
| 37 | HIV-1 | IC | [105] | ||
| 38 | L1 | HPV 16 | IC | GAL | [106] |
| 39 | L1 | HPV 6,11 16 | IC | GAL | [107] |
| 40 | VP1 with Puumala hantavirus nucleocapsid protein segments | Hamster polyomavirus | IC | Hybrid GAL10-PYK1 | [108] |
| 41 | M protein | Hepatitis B virus | IC | GAL10/CYC1 | [109] |
| 42 | VP1,2 | Goose hemorrhagic polyomavirus | IC | GAL | [110] |
| 43 | CEA/VP1 | Hamster polyomavirus | IC | GAL | [111] |
| 44 | E7 oncoprotein of HPV16 | Hepatitis B virus | IC | [112] | |
| 45 | Capsid protein | Red-spotted grouper nervous necrosis virus | IC | GAL10 | [113] |
| 46 | L1 | HPV 58 | IC | GAL10 | [114] |
| 47 | C69R variant of surface antigen | Hepatitis B virus | IC | GAL10/CYC1 | [115] |
| 48 | VP1 | Human polyomavirus 2 | IC | GAL | [116] |
| 49 | VP2 | Parvovirus B19 | IC | GAL1 | [117] |
| 50 | L1 | HPV 11 | IC | GAL110-11 | [118] |
| 51 | VP1 with pre-S1 region of the Hepatitis B virus | Hamster polyomavirus | IC | GAL | [119] |
| 52 | Surface antigen | Hepatitis B virus | IC | GAL10 | [120] |
| 53 | L1 | HPV 16 | IC | [121] |
| S. No | Protein Antigen | Disease | Secretion Localization | Promoter | References |
|---|---|---|---|---|---|
| 1 | Porcine circovirus type 2b capsid protein | Postweaning multisystemic wasting disease (PMWS) in pigs | IC | MOX (methanol) | [194] |
| 2 | Bovine viral diarrhea virus (BVDV) glycoprotein E2 | Bovine diarrhea | IC | FMD (formate dehydrogenase) | [195] |
| 3 | Membrane integral small surface protein (dS) of the duck Hepatitis B virus | Bovine diarrhea | IC | MOX (methanol) | [196] |
| 4 | Surface antigen | Hepatitis E | IC | [197] | |
| 5 | Sporozoite antigen | Malaria | IC | [198] | |
| 6 | Sporozoite antigen | Malaria | IC | MOX (methanol) | [199] |
| 7 | Glycoprotein E2 ectodomain | Hepatitis C | EC | [200] |
| S. No | Protein Antigen | Disease | Secretion Localization | Yeast Species | References |
|---|---|---|---|---|---|
| 1 | Capsid protein of porcine circovirus | Postweaning multisystemic wasting syndrome (PMWS) and porcine circovirus diseases (PCVDs) | IC | Kluyveromyces marxianus | [201] |
| 2 | VP1 of murine polyomavirus | Cancer | IC | Kluyveromyces lactis | [202] |
| 3 | VP2 of porcine parvovirus | Embryonic and fetal loss, death, and mummification | IC | Kluyveromyces marxianus | [203] |
| 4 | Capsid protein of red-spotted grouper nervous necrosis virus | Viral encephalopathy and retinopathy | IC | Yarrowia lipolytica | [204] |
| 5 | Totivirus capsid protein | NA | IC | Xanthophyllomyces dendrorhous | [205] |
| 6 | Tobacco mosaic virus coat protein | Mosaic-like mottling and discoloration on the leaves | IC | Schizosaccharomyces pombe | [206] |
| Trade Name | Infectious Agent | Disease | Company | Antigen | Status |
|---|---|---|---|---|---|
| Gardasil® | HPV | Cervical carcinoma | Merck | L1 | Approved |
| Gardasil9® | HPV | Cervical carcinoma | Merck | L1 | Approved |
| Mosquirix™ | P. falciparum | Malaria | Univ. of Rochester AVEG, Rochester, New York, United States | P. falciparum circumsporozoite protein fused to the Hepatitis B surface antigen | Approved |
| NA | HIV | Acquired immune deficiency syndrome | HIV p17/p24: Ty-VLP | Clinical trial Phase 1(Clinical trial No.: NCT00001053) [43] | |
| Hepavax-Gene | HBV | Hepatocellular carcinoma | Crucell (Dusseldorf-Germany) | SHBs, MHBs | Licensed * |
| Fendrix | HBV | Hepatocellular carcinoma | GSK (Belgium) | SHBs | Licensed |
| Heplisav-B | HBV | Hepatocellular carcinoma | Dynavax (Oakland-USA) | SHBs | Licensed * |
| Engerix | HBV | Hepatocellular carcinoma | GSK (UK) | SHBs | Licensed |
| Recombivax HB (H-B-Vac®-II) | HBV | Hepatocellular carcinoma | Merck Vaccine (Canada) | SHBs | Licensed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srivastava, V.; Nand, K.N.; Ahmad, A.; Kumar, R. Yeast-Based Virus-like Particles as an Emerging Platform for Vaccine Development and Delivery. Vaccines 2023, 11, 479. https://doi.org/10.3390/vaccines11020479
Srivastava V, Nand KN, Ahmad A, Kumar R. Yeast-Based Virus-like Particles as an Emerging Platform for Vaccine Development and Delivery. Vaccines. 2023; 11(2):479. https://doi.org/10.3390/vaccines11020479
Chicago/Turabian StyleSrivastava, Vartika, Kripa N. Nand, Aijaz Ahmad, and Ravinder Kumar. 2023. "Yeast-Based Virus-like Particles as an Emerging Platform for Vaccine Development and Delivery" Vaccines 11, no. 2: 479. https://doi.org/10.3390/vaccines11020479