
Candida parapsilosis Cell Wall Proteome Characterization and Effectiveness against Hematogenously Disseminated Candidiasis in a Murine Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Candida Species
2.2. Antifungal Susceptibility Profiling
2.3. Extraction of Cell Wall Proteins
2.4. Sample Preparation for LC–MS Analysis
2.5. LC–MS Data Acquisition
2.6. LC–MS Database Searching
2.7. Bioinformatic and Functional Analyses
2.8. In Vivo Study
2.9. Immunization
2.10. Histopathology
2.11. Statistical Analysis
3. Results
3.1. Antifungal Susceptibility Profiling
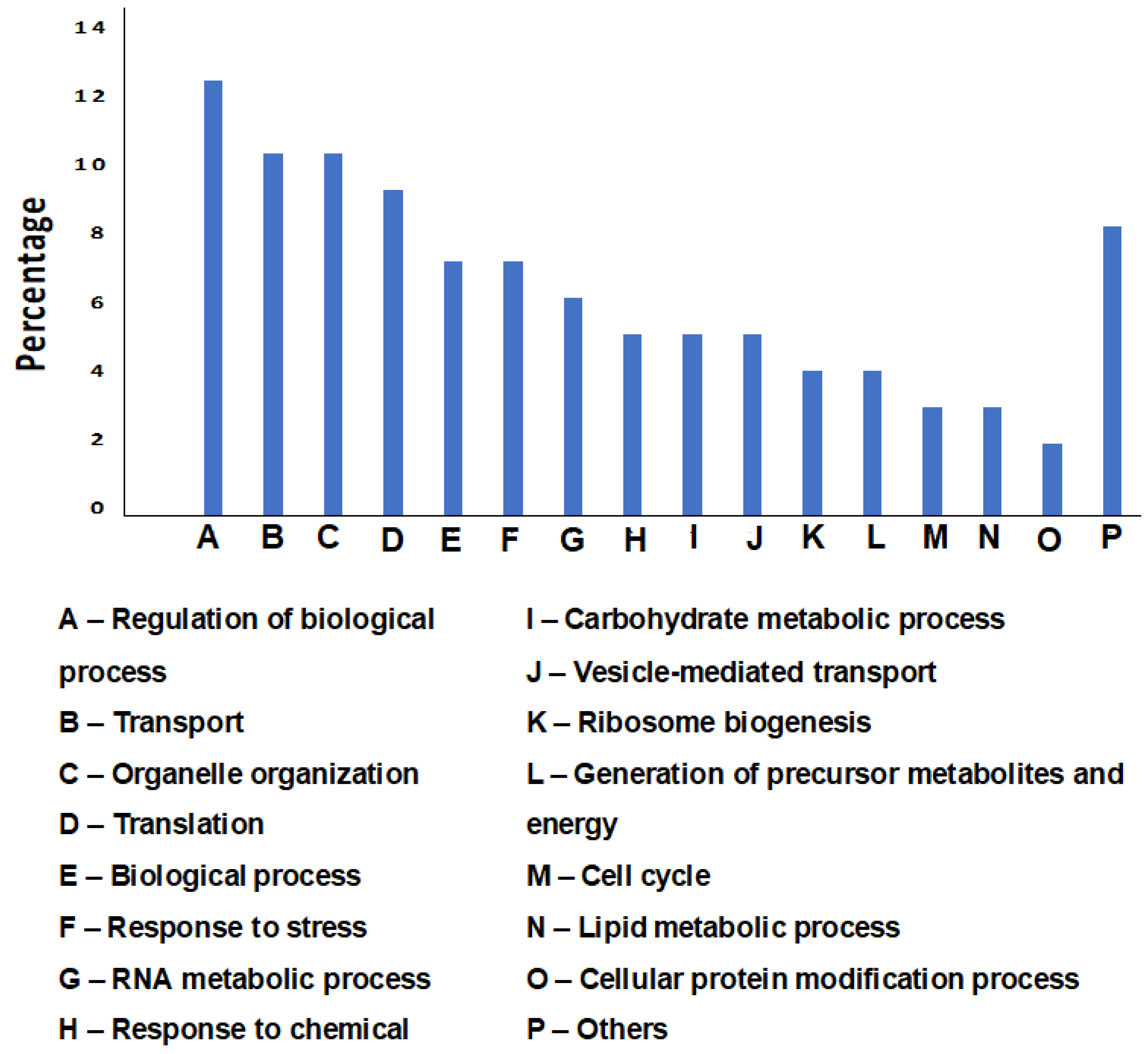
3.2. Proteomic Analysis of the Cell Wall-Associated Proteins and Protective Antigen Prediction
3.3. Functional Analysis of Predicted Antigens of C. parapsilosis MRU# 4112 Strain Cell Wall Extract
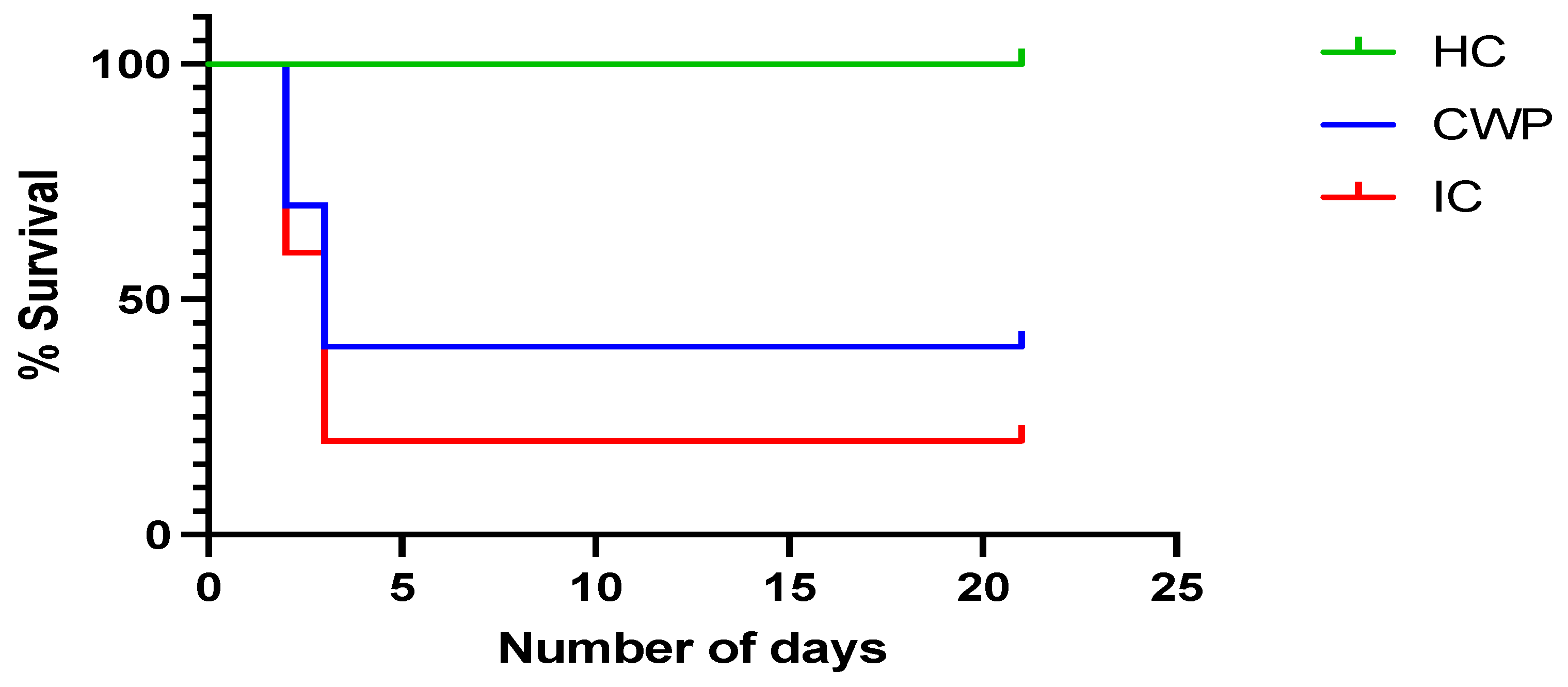
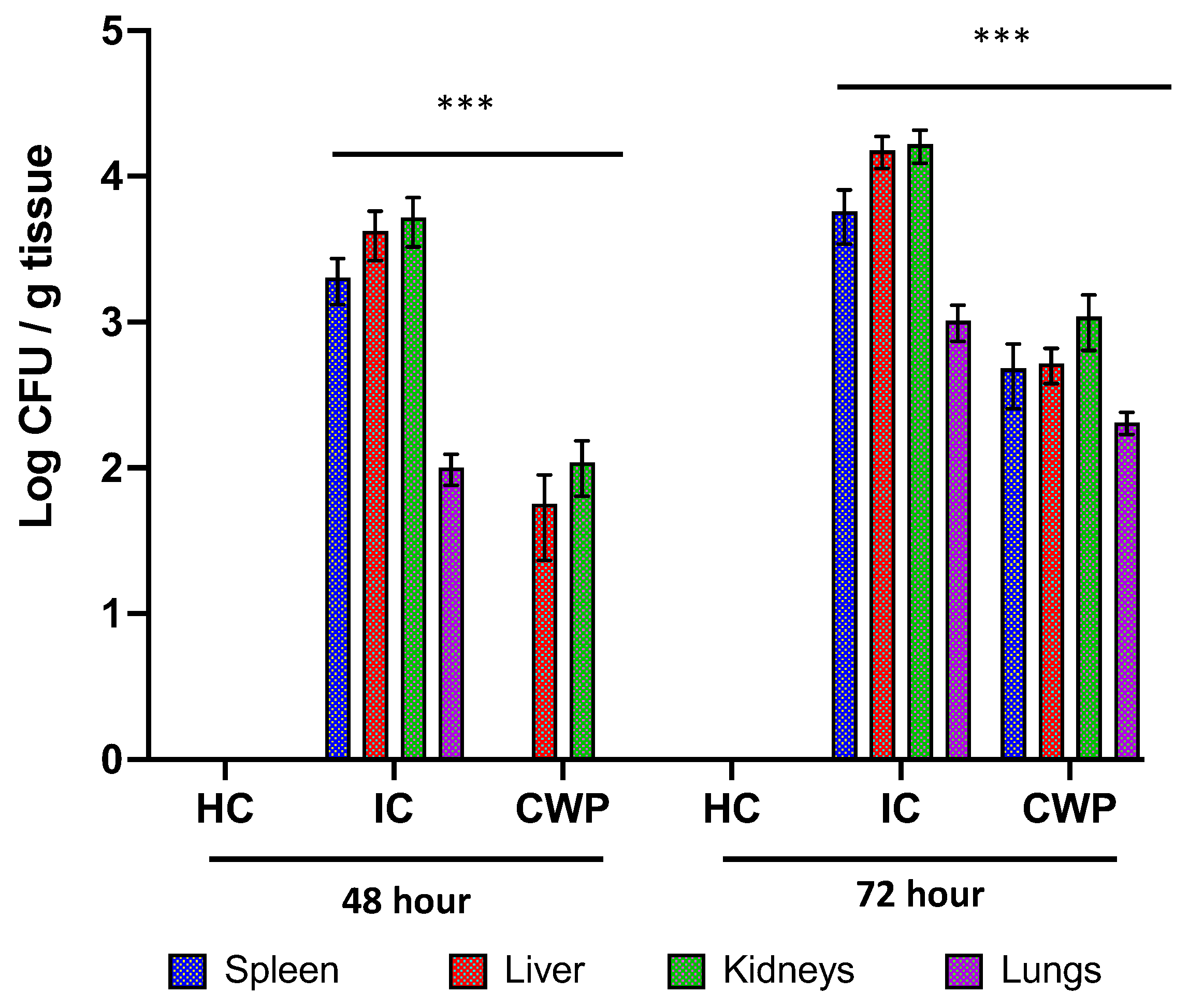
3.4. In Vivo Studies
Effectiveness of Immunization
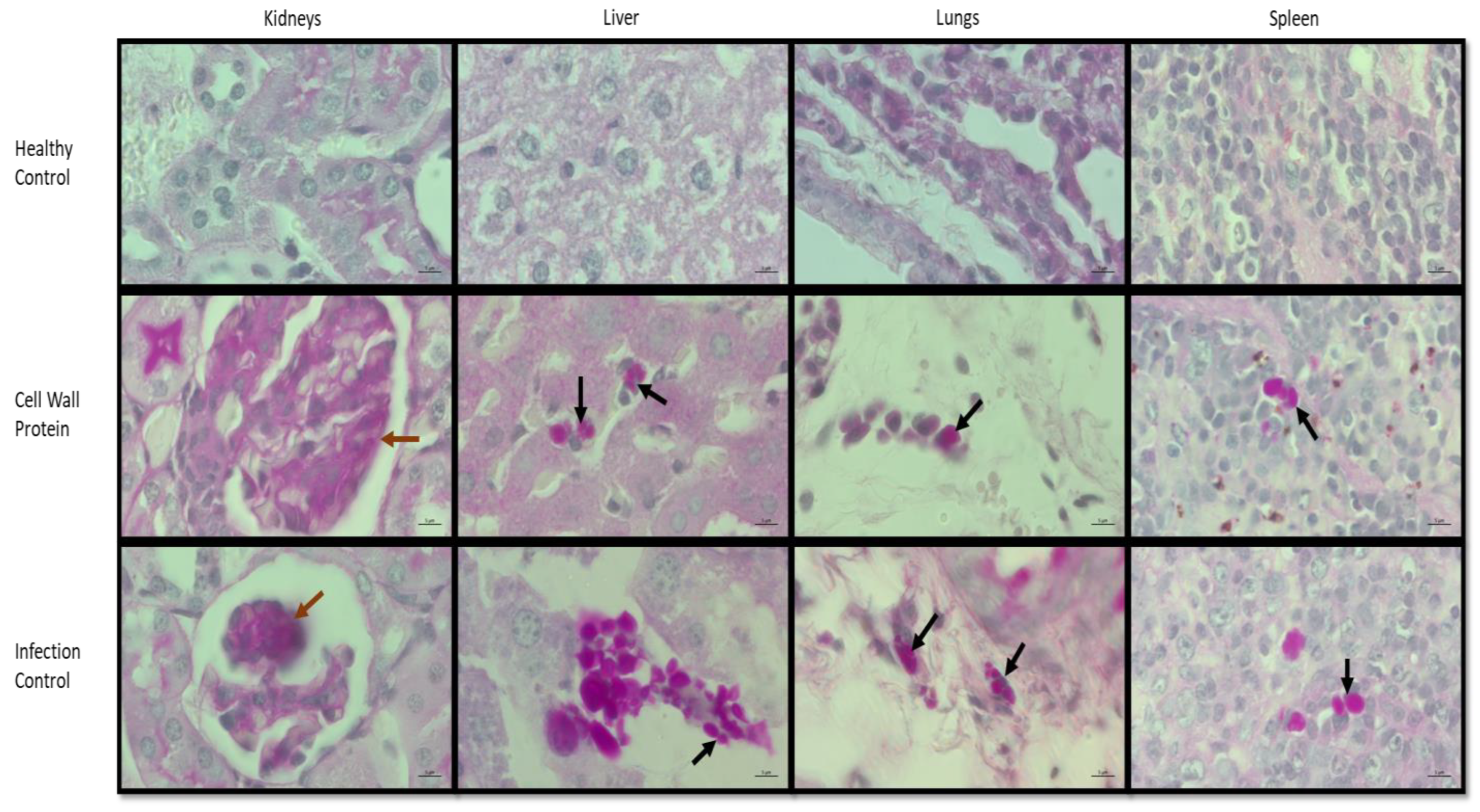
3.5. Histopathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species. Front. Microbiol. 2016, 7, 2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govender, N.P.; Patel, J.; Magobo, R.E.; Naicker, S.; Wadula, J.; Whitelaw, A.; Coovadia, Y.; Kularatne, R.; Govind, C.; Lockhart, S.R.; et al. Emergence of azole-resistant Candida parapsilosis causing bloodstream infection: Results from laboratory-based sentinel surveillance in South Africa. J. Antimicrob. Chemother. 2016, 71, 1994–2004. [Google Scholar] [CrossRef] [Green Version]
- Magobo, R.E.; Naicker, S.D.; Wadula, J.; Nchabeleng, M.; Coovadia, Y.; Hoosen, A.; Lockhart, S.R.; Govender, N.P.; the TRAC-South Africa Group. Detection of neonatal unit clusters of Candida parapsilosis fungaemia by microsatellite genotyping: Results from laboratory-based sentinel surveillance, South Africa, 2009–2010. Mycoses 2017, 60, 320–327. [Google Scholar] [CrossRef]
- Magobo, R.E.; Lockhart, S.R.; Govender, N.P. Fluconazole-resistant Candida parapsilosis strains with a Y132F substitution in the ERG11 gene causing invasive infections in a neonatal unit, South Africa. Mycoses 2020, 63, 471–477. [Google Scholar] [CrossRef]
- Trofa, D.; Gacser, A.; Nosanchuk, J.D. Candida parapsilosis, an emerging fungal pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gow, N.A.; van de Veerdonk, F.L.; Brown, A.J.; Netea, M.G. Candida albicans morphogenesis and host defence: Discriminating invasion from colonization. Nat. Rev. Microbiol. 2011, 10, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Free, S.J. Fungal cell wall organization and biosynthesis. Adv. Genet. 2013, 81, 33–82. [Google Scholar] [CrossRef]
- Perez-Garcia, L.A.; Csonka, K.; Flores-Carreon, A.; Estrada-Mata, E.; Mellado-Mojica, E.; Nemeth, T.; Lopez-Ramirez, L.A.; Toth, R.; Lopez, M.G.; Vizler, C.; et al. Role of Protein Glycosylation in Candida parapsilosis Cell Wall Integrity and Host Interaction. Front Microbiol 2016, 7, 306. [Google Scholar] [CrossRef] [Green Version]
- Nanjappa, S.G.; Klein, B.S. Vaccine immunity against fungal infections. Curr. Opin. Immunol. 2014, 28, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Piccione, D.; Mirabelli, S.; Minto, N.; Bouklas, T. Difficult but not impossible: In Search of an anti-Candida Vaccine. Curr. Trop. Med. Rep. 2019, 6, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Ahmadipour, S.; Field, R.A.; Miller, G.J. Prospects for anti-Candida therapy through targeting the cell wall: A mini-review. Cell Surf. 2021, 7, 100063. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Srivastava, V.; Ahmad, A. Abrogation of pathogenic attributes in drug resistant Candida auris strains by farnesol. PLoS ONE 2020, 15, e0233102. [Google Scholar] [CrossRef]
- El Khoury, P.; Awad, A.; Wex, B.; Khalaf, R.A. Proteomic analysis of a Candida albicans pir32 null strain reveals proteins involved in adhesion, filamentation and virulence. PLoS ONE 2018, 13, e0194403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Uppuluri, P.; Mamouei, Z.; Alqarihi, A.; Elhassan, H.; French, S.; Lockhart, S.R.; Chiller, T.; Edwards, J.E., Jr.; Ibra-him, A.S. The NDV-3A vaccine protects mice from multidrug resistant Candida auris infection. PLOS Pathog. 2019, 15, e1007460. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.; Horn, T.; Frimodt-Møller, N. In Vivo Pathogenicity of Eight Medically Relevant Candida Species in an Animal Model. Infection 2002, 30, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Mice with disseminated candidiasis die of progressive sepsis. J. Infect. Dis. 2005, 192, 336–343. [Google Scholar] [CrossRef]
- Trammell, R.A.; Toth, L.A. Markers for predicting death as an outcome for mice used in infectious disease research. Comp. Med. 2011, 61, 492–498. Available online: http://aalas.publisher.ingentaconnect.com/content/aalas/cm/2011/00000061/00000006/art00002# (accessed on 20 February 2023). [PubMed]
- Dimopoulou, D.; Hamilos, G.; Tzardi, M.; Lewis, R.E.; Samonis, G.; Kontoyiannis, D.P. Anidulafungin versus caspofungin in a mouse model of candidiasis caused by anidulafungin-susceptible Candida parapsilosis isolates with different degrees of caspofungin susceptibility. Antimicrob. Agents Chemother. 2014, 58, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, T.; Miyazaki, T.; Ito, Y.; Wakayama, M.; Shibuya, K.; Yamashita, K.; Takazono, T.; Saijo, T.; Shimamura, S.; Yamamoto, K.; et al. Virulence assessment of six major pathogenic Candida species in the mouse model of invasive candidiasis caused by fungal translocation. Sci. Rep. 2020, 10, 3814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer, L.L.; Green, C.B.; Oh, S.H.; Zhao, X. Discovering the secrets of the Candida albicans agglutinin-like sequence (ALS) gene family—A sticky pursuit. Med. Mycol. 2008, 46, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, M.N.; Glass, K.A.; Longley, S.J.; Kim, D.J.; Laforce-Nesbitt, S.S.; Wortzel, J.D.; Shaw, S.K.; Bliss, J.M. Role of the Inducible Adhesin CpAls7 in Binding of Candida parapsilosis to the Extracellular Matrix under Fluid Shear. Infect. Immun. 2018, 86, e00892-17. [Google Scholar] [CrossRef] [Green Version]
- Bertini, A.; Zoppo, M.; Lombardi, L.; Rizzato, C.; De Carolis, E.; Vella, A.; Torelli, R.; Sanguinetti, M.; Tavanti, A. Targeted gene disruption in Candida parapsilosis demonstrates a role for CPAR2_404800 in adhesion to a biotic surface and in a murine model of ascending urinary tract infection. Virulence 2016, 7, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Karkowska-Kuleta, J.; Zajac, D.; Bras, G.; Bochenska, O.; Rapala-Kozik, M.; Kozik, A. Binding of human plasminogen and high-molecular-mass kininogen by cell surface-exposed proteins of Candida parapsilosis. Acta Biochim. Pol. 2017, 64, 391–400. [Google Scholar] [CrossRef]
- Duggan, S.; Leonhardt, I.; Hunniger, K.; Kurzai, O. Host response to Candida albicans bloodstream infection and sepsis. Virulence 2015, 6, 316–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, K.; Schorr, C.; Reboli, A.C.; Zanotti, S.; Tsigrelis, C. Incidence and mortality of sepsis, severe sepsis, and septic shock in intensive care unit patients with candidemia. Infect. Dis. 2015, 47, 584–587. [Google Scholar] [CrossRef]
- Lepingle, A.; Casaregola, S.; Neuveglise, C.; Bon, E.; Nguyen, H.; Artiguenave, F.; Wincker, P.; Gaillardin, C. Genomic exploration of the hemiascomycetous yeasts: 14. Debaryomyces hansenii var. hansenii. FEBS Lett. 2000, 487, 82–86. [Google Scholar] [CrossRef] [Green Version]
- Rooney, J.P.; Patil, A.; Joseph, F.; Endres, L.; Begley, U.; Zappala, M.R.; Cunningham, R.P.; Begley, T.J. Cross-species Functionome analysis identifies proteins associated with DNA repair, translation and aerobic respiration as conserved modulators of UV-toxicity. Genomics 2011, 97, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Sanchez, S.; Aubert, S.; Iraqui, I.; Janbon, G.; Ghigo, J.M.; d’Enfert, C. Candida albicans biofilms: A developmental state associated with specific and stable gene expression patterns. Eukaryot. Cell 2004, 3, 536–545. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Lee, R.E.; Barker, K.S.; Lee, R.E.; Wei, L.; Homayouni, R.; Rogers, P.D. Genome-wide expression profiling of the response to azole, polyene, echinocandin, and pyrimidine antifungal agents in Candida albicans. Antimicrob. Agents Chemother. 2005, 49, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Cabezon, V.; Llama-Palacios, A.; Nombela, C.; Monteoliva, L.; Gil, C. Analysis of Candida albicans plasma membrane proteome. Proteomics 2009, 9, 4770–4786. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, C.L.; Wykoff, D.D. Candida glabrata PHO4 is necessary and sufficient for Pho2-independent transcription of phosphate starvation genes. Genetics 2009, 182, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Teichert, U.; Mechler, B.; Muller, H.; Wolf, D.H. Lysosomal (vacuolar) proteinases of yeast are essential catalysts for protein degradation, differentiation, and cell survival. J. Biol. Chem. 1989, 264, 16037–16045. [Google Scholar] [CrossRef]
- Zubenko, G.S.; Jones, E.W. Protein degradation, meiosis and sporulation in proteinase-deficient mutants of Saccharomyces cerevisiae. Genetics 1981, 97, 45–64. [Google Scholar] [CrossRef]
- Hecht, K.A.; O’Donnell, A.F.; Brodsky, J.L. The proteolytic landscape of the yeast vacuole. Cell. Logist. 2014, 4, e28023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Bona, A.; Parra-Giraldo, C.M.; Hernaez, M.L.; Reales-Calderon, J.A.; Solis, N.V.; Filler, S.G.; Monteoliva, L.; Gil, C. Candida albicans cell shaving uncovers new proteins involved in cell wall integrity, yeast to hypha transition, stress response and host-pathogen interaction. J. Proteom. 2015, 127, 340–351. [Google Scholar] [CrossRef] [Green Version]
- Naik, R.R.; Nebes, V.; Jones, E.W. Regulation of the proteinase B structural gene PRB1 in Saccharomyces cerevisiae. J. Bacteriol. 1997, 179, 1469–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Vashisht, A.A.; Tan, Y.; Su, T.; Wohlschlegel, J.A. PRB1 is required for clipping of the histone H3 N terminal tail in Saccharomyces cerevisiae. PLoS ONE 2014, 9, e90496. [Google Scholar] [CrossRef] [Green Version]
- Monteoliva, L.; Matas, M.L.; Gil, C.; Nombela, C.; Pla, J. Large-scale identification of putative exported proteins in Candida albicans by genetic selection. Eukaryot. Cell 2002, 1, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Fradin, C.; De Groot, P.; MacCallum, D.; Schaller, M.; Klis, F.; Odds, F.C.; Hube, B. Granulocytes govern the transcriptional response, morphology and proliferation of Candida albicans in human blood. Mol. Microbiol. 2005, 56, 397–415. [Google Scholar] [CrossRef]
- Roetzer, A.; Gregori, C.; Jennings, A.M.; Quintin, J.; Ferrandon, D.; Butler, G.; Kuchler, K.; Ammerer, G.; Schuller, C. Candida glabrata environmental stress response involves Saccharomyces cerevisiae Msn2/4 orthologous transcription factors. Mol. Microbiol. 2008, 69, 603–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Martinez, A.E.; Gomez-Molero, E.; Sanchez-Virosta, P.; Dekker, H.L.; de Boer, A.; Eraso, E.; Bader, O.; de Groot, P.W.J. High Biofilm Formation of Non-Smooth Candida parapsilosis Correlates with Increased Incorporation of GPI-Modified Wall Adhesins. Pathogens 2021, 10, 493. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.P.; Miranda, I.M.; Guida, A.; Synnott, J.; Rocha, R.; Silva, R.; Amorim, A.; Pina-Vaz, C.; Butler, G.; Rodrigues, A.G. Transcriptional profiling of azole-resistant Candida parapsilosis strains. Antimicrob. Agents Chemother. 2011, 55, 3546–3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhof, T.; Seibold, M.; Thewes, S.; Laue, M.; Han, C.O.; Hube, B.; von Dohren, H. Comparison of susceptibility and transcription profile of the new antifungal hassallidin A with caspofungin. Biochem. Biophys. Res. Commun. 2006, 349, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Gil-Bona, A.; Monteoliva, L.; Gil, C. Global Proteomic Profiling of the Secretome of Candida albicans ecm33 Cell Wall Mutant Reveals the Involvement of Ecm33 in Sap2 Secretion. J. Proteome Res. 2015, 14, 4270–4281. [Google Scholar] [CrossRef]
- Medrano-Diaz, C.L.; Vega-Gonzalez, A.; Ruiz-Baca, E.; Moreno, A.; Cuellar-Cruz, M. Moonlighting proteins induce protection in a mouse model against Candida species. Microb. Pathog. 2018, 124, 21–29. [Google Scholar] [CrossRef]
- Shen, H.; Yu, Y.; Chen, S.M.; Sun, J.J.; Fang, W.; Guo, S.Y.; Hou, W.T.; Qiu, X.R.; Zhang, Y.; Chen, Y.L.; et al. Dectin-1 Facilitates IL-18 Production for the Generation of Protective Antibodies Against Candida albicans. Front. Microbiol. 2020, 11, 1648. [Google Scholar] [CrossRef]
- Nami, S.; Mohammadi, R.; Vakili, M.; Khezripour, K.; Mirzaei, H.; Morovati, H. Fungal vaccines, mechanism of actions and immunology: A comprehensive review. Biomed. Pharmacother. 2019, 109, 333–344. [Google Scholar] [CrossRef]
- Shi, H.; Dong, S.; Zhang, X.; Chen, X.; Gao, X.; Wang, L. Phage vaccines displaying YGKDVKDLFDYAQE epitope induce protection against systemic candidiasis in mouse model. Vaccine 2018, 36, 5717–5724. [Google Scholar] [CrossRef]
- Alqarihi, A.; Singh, S.; Edwards, J.E., Jr.; Ibrahim, A.S.; Uppuluri, P. NDV-3A vaccination prevents C. albicans colonization of jugular vein catheters in mice. Sci. Rep. 2019, 9, 6194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, A.S.; Luo, G.; Gebremariam, T.; Lee, H.; Schmidt, C.S.; Hennessey, J.P., Jr.; French, S.W.; Yeaman, M.R.; Filler, S.G.; Edwards, J.E., Jr. NDV-3 protects mice from vulvovaginal candidiasis through T- and B-cell immune response. Vaccine 2013, 31, 5549–5556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D.; Turner, S.A.; Butler, G.; Vagvolgyi, C.; Gacser, A. Candida parapsilosis: From Genes to the Bedside. Clin. Microbiol. Rev. 2019, 32, e00111-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 01IS MRU#: 4315 | 02IS MRU#: 4282 | 03IS MRU#: 4090 | 04IS MRU#: 4112 | ||||
| µg/mL | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC |
| FLZ | 312.500 | FS | 625.000 | FS | 312.500 | FS | 312.500 | FS |
| AmB | 0.977 | 1.953 | 0.977 | 1.953 | 0.977 | 1.953 | 31.250 | 62.500 |
| NYST | 62.500 | 125.000 | 31.250 | FS | 62.500 | FS | 125.000 | FS |
| CAS | 0.125 | 0.250 | 0.125 | 0.250 | 0.125 | 0.250 | 0.500 | 1.000 |
| Strain | 05IS MRU#: 4120 | 06IS MRU#: 4334 | 07IS MRU#: 4033 | 08IS MRU#: 4108 | ||||
| µg/mL | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC |
| FLZ | 312.500 | FS | 312.500 | FS | 156.250 | FS | 312.500 | FS |
| AmB | 1.953 | 31.250 | 1.953 | 7.813 | 3.906 | 62.500 | 15.625 | 31.250 |
| NYST | 62.500 | 250.000 | 125.000 | FS | 62.500 | FS | 31.250 | FS |
| CAS | 0.250 | 0.500 | 0.125 | 0.500 | 0.250 | 2.000 | 0.125 | 0.250 |
| Strain | 09IS MRU#: 4036 | 10IS MRU#: 4086 | 11IS MRU#: 5473 | 12IS MRU#: 5503 | ||||
| µg/mL | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC |
| FLZ | 312.500 | FS | 312.500 | FS | 625.000 | FS | 312.500 | FS |
| AmB | 7.813 | 31.250 | 7.813 | 31.250 | 7.813 | 62.500 | 1.953 | 31.250 |
| NYST | 31.250 | 62.500 | 15.625 | 31.250 | 7.813 | 125.000 | 31.250 | 125.000 |
| CAS | 0.500 | 2.000 | 0.063 | 0.125 | 0.125 | 0.250 | 1.000 | 4.000 |
| Gene | Protein Names | Localization | Type |
|---|---|---|---|
| CPAR2_100640 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_104370 | Uncharacterized protein | Nucleus | Soluble |
| CPAR2_104690 | GTP-binding nuclear protein | Cytoplasm | Soluble |
| CPAR2_105980 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_106320 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_109010 | Uncharacterized protein | Plastid | Soluble |
| CPAR2_110110 | HTH cro/C1-type domain-containing protein | Nucleus | Soluble |
| CPAR2_201550 | Glycine cleavage system H protein | Mitochondrion | Soluble |
| CPAR2_203410 | Uncharacterized protein | Mitochondrion | Soluble |
| CPAR2_204240 | Proteosome_alpha_1 domain-containing protein | Nucleus | Soluble |
| CPAR2_205620 | Ribosomal_S10 domain-containing protein | Cytoplasm | Soluble |
| CPAR2_207050 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_211070 | NET domain-containing protein | Nucleus | Soluble |
| CPAR2_212330 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_300430 | Uncharacterized protein | Nucleus | Soluble |
| CPAR2_302060 | Histone-glutamine methyltransferase | Nucleus | Soluble |
| CPAR2_304080 | Adenylyl-sulfate kinase | Cytoplasm | Soluble |
| CPAR2_401210 | Proliferating cell nuclear antigen | Nucleus | Soluble |
| CPAR2_403290 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_404800 | Candida ALS N domain-containing protein | Cell surface | Membrane |
| CPAR2_405130 | Uncharacterized protein | Mitochondrion | Membrane |
| CPAR2_503400 | Pre-mRNA-processing protein 46 | Nucleus | Soluble |
| CPAR2_503520 | Branched-chain-amino-acid aminotransferase | Cytoplasm | Soluble |
| CPAR2_504170 | 40S ribosomal protein S24 | Cytoplasm | Soluble |
| CPAR2_600900 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_601260 | CS domain-containing protein | Nucleus | Soluble |
| CPAR2_602580 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_701270 | Uncharacterized protein | Endoplasmic reticulum | Membrane |
| CPAR2_701500 | Uncharacterized protein | Cytoplasm | Soluble |
| CPAR2_701770 | H/ACA ribonucleoprotein complex subunit NOP10 | Nucleus | Soluble |
| CPAR2_703940 | Uncharacterized protein | Extracellular | Soluble |
| CPAR2_800880 | Actin-related protein 2/3 complex subunit 4 | Cytoplasm | Soluble |
| CPAR2_802120 | ANAPC4_WD40 domain-containing protein | Nucleus | Soluble |
| CPAR2_805040 | Uncharacterized protein | Extracellular | Soluble |
| Gene | Protein Names | C. albicans Ortholog Gene | C. albicans Ortholog Protein Name |
|---|---|---|---|
| CPAR2_404800 | Candida ALS N domain-containing protein | ALS7/C3_06320W | Agglutinin-like protein 7 |
| CPAR2_405130 | Uncharacterized protein | C3_02270W | Uncharacterized protein |
| CPAR2_701270 | Uncharacterized protein | PHO88/CR_09320C | Inorganic phosphate transport protein PHO88 |
| CPAR2_703940 | Uncharacterized protein | C7_03860W | Uncharacterized protein |
| CPAR2_805040 | Uncharacterized protein | C1_10170W | Uncharacterized protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Srivastava, V.; Naicker, P.; Khan, A.; Ahmad, A. Candida parapsilosis Cell Wall Proteome Characterization and Effectiveness against Hematogenously Disseminated Candidiasis in a Murine Model. Vaccines 2023, 11, 674. https://doi.org/10.3390/vaccines11030674
Gong X, Srivastava V, Naicker P, Khan A, Ahmad A. Candida parapsilosis Cell Wall Proteome Characterization and Effectiveness against Hematogenously Disseminated Candidiasis in a Murine Model. Vaccines. 2023; 11(3):674. https://doi.org/10.3390/vaccines11030674
Chicago/Turabian StyleGong, Xiaolong, Vartika Srivastava, Previn Naicker, Amber Khan, and Aijaz Ahmad. 2023. "Candida parapsilosis Cell Wall Proteome Characterization and Effectiveness against Hematogenously Disseminated Candidiasis in a Murine Model" Vaccines 11, no. 3: 674. https://doi.org/10.3390/vaccines11030674