
Ebola Virus Disease Vaccines: Development, Current Perspectives & Challenges
 , ,
, ,  , ,
, ,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. An Overview of Current Vaccines
2.1. Virus-Like Particles Vaccines
2.2. DNA Vaccines
2.3. Recombinant Whole Virus Vaccine
2.4. Replication Incompetent Vaccines
2.5. Replication Competent Vaccines
3. Human Clinical Trials of Ebola Vaccines
4. Animal Models for EVD Vaccine Development
5. Challenges for Vaccine Development
5.1. Selection of Immunogen
5.2. Rapid-Responding Vaccination
5.3. Cross-Protective Immunity
5.4. Long-Term Protection
5.5. Mechanism of Protection
6. Current Vaccine Considerations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly, J.D.; Frankfurter, R.G.; Tavs, J.M.; Barrie, M.B.; McGinnis, T.; Kamara, M.; Freeman, A.; Quiwah, K.; Davidson, M.C.; Dighero-Kemp, B.; et al. Association of Lower Exposure Risk with Paucisymptomatic/Asymptomatic Infection, Less Severe Disease, and Unrecognized Ebola Virus Disease: A Seroepidemiological Study. Open Forum Infect. Dis. 2022, 9, ofac052. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.B.; Li, Z.X.; Du, Y.; Cao, G.W. Ebola virus disease: From epidemiology to prophylaxis. Mil. Med. Res. 2015, 2, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weyer, J.; Grobbelaar, A.; Blumberg, L. Ebola virus disease: History, epidemiology and outbreaks. Curr. Infect. Dis. Rep. 2015, 17, 480. [Google Scholar] [CrossRef] [PubMed]
- Mondiale de la Santé, O.; World Health Organization. WHO’s Health Emergencies Programme: Acute emergencies monthly summary–October and November 2022—Programme OMS de gestion des situations d’urgence sanitaire: Résumé mensuel des situations d’urgence aiguë–octobre et novembre 2022. Wkly. Epidemiol. Rec. Relev. Épidémiol. Hebd. 2022, 97, 640–644. [Google Scholar]
- Chang, T.; Wu, J.; Chang, L. Clinical characteristics and diagnostic challenges of pediatric COVID-19: A systematic review and meta-analysis. J. Formos. Med. Assoc. 2020, 119, 982–989. [Google Scholar] [CrossRef]
- Sridhar, S. Clinical development of Ebola vaccines. Ther. Adv. Vaccines 2015, 3, 125–138. [Google Scholar] [CrossRef]
- Warfield, K.L.; Olinger, G.; Deal, E.M.; Swenson, D.L.; Bailey, M.; Negley, D.L.; Hart, M.K.; Bavari, S. Induction of humoral and CD8+ T cell responses are required for protection against lethal Ebola virus infection. J. Immunol. 2005, 175, 1184–1191. [Google Scholar] [CrossRef] [Green Version]
- Warfield, K.L.; Swenson, D.L.; Olinger, G.G.; Kalina, W.V.; Aman, M.J.; Bavari, S. Ebola virus-like particle-based vaccine protects nonhuman primates against lethal Ebola virus challenge. J. Infect. Dis. 2007, 196, S430–S437. [Google Scholar] [CrossRef] [Green Version]
- Warfield, K.L.; Aman, M.J. Advances in virus-like particle vaccines for filoviruses. J. Infect. Dis. 2011, 204, S1053–S1059. [Google Scholar] [CrossRef]
- Lu, S.; Wang, S.; Grimes-Serrano, J.M. Current progress of DNA vaccine studies in humans. Expert Rev. Vaccines 2008, 7, 175–191. [Google Scholar] [CrossRef]
- Martin, J.E.; Sullivan, N.J.; Enama, M.E.; Gordon, I.J.; Roederer, M.; Koup, R.A.; Bailer, R.T.; Chakrabarti, B.K.; Bailey, M.A.; Gomez, P.L. A DNA vaccine for Ebola virus is safe and immunogenic in a phase I clinical trial. Clin. Vaccine Immunol. 2006, 13, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Grant-Klein, R.J.; Van Deusen, N.M.; Badger, C.V.; Hannaman, D.; Dupuy, L.C.; Schmaljohn, C.S. A multiagent filovirus DNA vaccine delivered by intramuscular electroporation completely protects mice from ebola and Marburg virus challenge. Hum. Vaccines Immunother. 2012, 8, 1703–1706. [Google Scholar] [CrossRef] [Green Version]
- Shedlock, D.J.; Aviles, J.; Talbott, K.T.; Wong, G.; Wu, S.J.; Villarreal, D.O.; Myles, D.J.; Croyle, M.A.; Yan, J.; Kobinger, G.P. Induction of broad cytotoxic T cells by protective DNA vaccination against Marburg and Ebola. Mol. Ther. 2013, 21, 1432–1444. [Google Scholar] [CrossRef] [Green Version]
- Gustafsson, C.; Govindarajan, S.; Minshull, J. Codon bias and heterologous protein expression. Trends Biotechnol. 2004, 22, 346–353. [Google Scholar] [CrossRef]
- Grant-Klein, R.J.; Altamura, L.A.; Badger, C.V.; Bounds, C.E.; Van Deusen, N.M.; Kwilas, S.A.; Vu, H.A.; Warfield, K.L.; Hooper, J.W.; Hannaman, D. Codon-optimized filovirus DNA vaccines delivered by intramuscular electroporation protect cynomolgus macaques from lethal Ebola and Marburg virus challenges. Hum. Vaccines Immunother. 2015, 11, 1991–2004. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Hu, Y.; Liang, Q.; Wei, M.; Zhu, F. Ebola vaccines in clinical trial: The promising candidates. Hum. Vaccines Immunother. 2017, 13, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Kibuuka, H.; Berkowitz, N.M.; Millard, M.; Enama, M.E.; Tindikahwa, A.; Sekiziyivu, A.B.; Costner, P.; Sitar, S.; Glover, D.; Hu, Z.; et al. Safety and immunogenicity of Ebola virus and Marburg virus glycoprotein DNA vaccines assessed separately and concomitantly in healthy Ugandan adults: A phase 1b, randomised, double-blind, placebo-controlled clinical trial. Lancet 2015, 385, 1545–1554. [Google Scholar] [CrossRef]
- Sarwar, U.N.; Costner, P.; Enama, M.E.; Berkowitz, N.; Hu, Z.; Hendel, C.S.; Sitar, S.; Plummer, S.; Mulangu, S.; Bailer, R.T. Safety and immunogenicity of DNA vaccines encoding Ebolavirus and Marburgvirus wild-type glycoproteins in a phase I clinical trial. J. Infect. Dis. 2015, 211, 549–557. [Google Scholar] [CrossRef]
- Rudge, T.L.; Machesky, N.J.; Sankovich, K.A.; Lemmon, E.E.; Badorrek, C.S.; Overman, R.; Niemuth, N.A.; Anderson, M.S. Assays for the Evaluation of the Immune Response to Marburg and Ebola Sudan Vaccination—Filovirus Animal Nonclinical Group Anti-Marburg Virus Glycoprotein Immunoglobulin G Enzyme-Linked Immunosorbent Assay and a Pseudovirion Neutralization Assay. Vaccines 2022, 10, 1211. [Google Scholar]
- Halfmann, P.; Kim, J.H.; Ebihara, H.; Noda, T.; Neumann, G.; Feldmann, H.; Kawaoka, Y. Generation of biologically contained Ebola viruses. Proc. Natl. Acad. Sci. USA 2008, 105, 1129–1133. [Google Scholar] [CrossRef] [Green Version]
- Marzi, A.; Feldmann, H. Ebola virus vaccines: An overview of current approaches. Expert Rev. Vaccines 2014, 13, 521–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensley, L.E.; Mulangu, S.; Asiedu, C.; Johnson, J.; Honko, A.N.; Stanley, D.; Fabozzi, G.; Nichol, S.T.; Ksiazek, T.G.; Rollin, P.E.; et al. Demonstration of cross-protective vaccine immunity against an emerging pathogenic Ebolavirus Species. PLoS Pathog. 2010, 6, e1000904. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, N.J.; Sanchez, A.; Rollin, P.E.; Yang, Z.Y.; Nabel, G.J. Development of a preventive vaccine for Ebola virus infection in primates. Nature 2000, 408, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.; Richardson, J.S.; Pillet, S.; Patel, A.; Qiu, X.; Alimonti, J.; Hogan, J.; Zhang, Y.; Takada, A.; Feldmann, H.; et al. Immune parameters correlate with protection against ebola virus infection in rodents and nonhuman primates. Sci. Transl. Med. 2012, 4, 158ra146. [Google Scholar] [CrossRef] [Green Version]
- Geisbert, T.W.; Pushko, P.; Anderson, K.; Smith, J.; Davis, K.J.; Jahrling, P.B. Evaluation in nonhuman primates of vaccines against Ebola virus. Emerg. Infect. Dis. 2002, 8, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, Y.; Caposio, P.; Parkins, C.J.; Botto, S.; Messaoudi, I.; Cicin-Sain, L.; Feldmann, H.; Jarvis, M.A. A replicating cytomegalovirus-based vaccine encoding a single Ebola virus nucleoprotein CTL epitope confers protection against Ebola virus. PLoS Negl. Trop. Dis. 2011, 5, e1275. [Google Scholar] [CrossRef] [Green Version]
- Durbin, A.P.; Skiadopoulos, M.H.; McAuliffe, J.M.; Riggs, J.M.; Surman, S.R.; Collins, P.L.; Murphy, B.R. Human parainfluenza virus type 3 (PIV3) expressing the hemagglutinin protein of measles virus provides a potential method for immunization against measles virus and PIV3 in early infancy. J. Virol. 2000, 74, 6821–6831. [Google Scholar] [CrossRef] [Green Version]
- Bukreyev, A.; Yang, L.; Zaki, S.R.; Shieh, W.J.; Rollin, P.E.; Murphy, B.R.; Collins, P.L.; Sanchez, A. A single intranasal inoculation with a paramyxovirus-vectored vaccine protects guinea pigs against a lethal-dose Ebola virus challenge. J. Virol. 2006, 80, 2267–2279. [Google Scholar] [CrossRef] [Green Version]
- Marzi, A.; Ebihara, H.; Callison, J.; Groseth, A.; Williams, K.J.; Geisbert, T.W.; Feldmann, H. Vesicular stomatitis virus-based Ebola vaccines with improved cross-protective efficacy. J. Infect. Dis. 2011, 204 (Suppl. 3), S1066–S1074. [Google Scholar] [CrossRef]
- Henao-Restrepo, A.M.; Camacho, A.; Longini, I.M.; Watson, C.H.; Edmunds, W.J.; Egger, M.; Carroll, M.W.; Dean, N.E.; Diatta, I.; Doumbia, M.; et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: Final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ça Suffit!). Lancet 2017, 389, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Milligan, I.D.; Gibani, M.M.; Sewell, R.; Clutterbuck, E.A.; Campbell, D.; Plested, E.; Nuthall, E.; Voysey, M.; Silva-Reyes, L.; McElrath, M.J.; et al. Safety and Immunogenicity of Novel Adenovirus Type 26- and Modified Vaccinia Ankara-Vectored Ebola Vaccines: A Randomized Clinical Trial. JAMA 2016, 315, 1610–1623. [Google Scholar] [CrossRef]
- Zhu, F.C.; Wurie, A.H.; Hou, L.H.; Liang, Q.; Li, Y.H.; Russell, J.B.; Wu, S.P.; Li, J.X.; Hu, Y.M.; Guo, Q.; et al. Safety and immunogenicity of a recombinant adenovirus type-5 vector-based Ebola vaccine in healthy adults in Sierra Leone: A single-centre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 389, 621–628. [Google Scholar] [CrossRef] [Green Version]
- Ewer, K.; Rampling, T.; Venkatraman, N.; Bowyer, G.; Wright, D.; Lambe, T.; Imoukhuede, E.B.; Payne, R.; Fehling, S.K.; Strecker, T.; et al. A Monovalent Chimpanzee Adenovirus Ebola Vaccine Boosted with MVA. N. Engl. J. Med. 2016, 374, 1635–1646. [Google Scholar] [CrossRef]
- Woolsey, C.; Geisbert, T.W. Current state of Ebola virus vaccines: A snapshot. PLoS Pathog. 2021, 17, e1010078. [Google Scholar] [CrossRef]
- Blaney, J.E.; Marzi, A.; Willet, M.; Papaneri, A.B.; Wirblich, C.; Feldmann, F.; Holbrook, M.; Jahrling, P.; Feldmann, H.; Schnell, M.J. Antibody quality and protection from lethal Ebola virus challenge in nonhuman primates immunized with rabies virus based bivalent vaccine. PLoS Pathog. 2013, 9, e1003389. [Google Scholar] [CrossRef]
- Marzi, A.; Halfmann, P.; Hill-Batorski, L.; Feldmann, F.; Shupert, W.L.; Neumann, G.; Feldmann, H.; Kawaoka, Y. Vaccines. An Ebola whole-virus vaccine is protective in nonhuman primates. Science 2015, 348, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.K.; Xu, R.; Matassov, D.; Latham, T.E.; Ota-Setlik, A.; Gerardi, C.S.; Luckay, A.; Witko, S.E.; Hermida, L.; Higgins, T.; et al. Safety and immunogenicity of a highly attenuated rVSVN4CT1-EBOVGP1 Ebola virus vaccine: A randomised, double-blind, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis. 2020, 20, 455–466. [Google Scholar] [CrossRef]
- Fries, L.; Cho, I.; Krähling, V.; Fehling, S.K.; Strecker, T.; Becker, S.; Hooper, J.W.; Kwilas, S.A.; Agrawal, S.; Wen, J.; et al. Randomized, Blinded, Dose-Ranging Trial of an Ebola Virus Glycoprotein Nanoparticle Vaccine with Matrix-M Adjuvant in Healthy Adults. J. Infect. Dis. 2020, 222, 572–582. [Google Scholar] [CrossRef]
- Matz, K.M.; Marzi, A.; Feldmann, H. Ebola vaccine trials: Progress in vaccine safety and immunogenicity. Expert Rev. Vaccines 2019, 18, 1229–1242. [Google Scholar] [CrossRef]
- Sharma, A.R.; Lee, Y.H.; Nath, S.; Lee, S.S. Recent developments and strategies of Ebola virus vaccines. Curr. Opin. Pharmacol. 2021, 60, 46–53. [Google Scholar] [CrossRef]
- Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatulin, A.I.; Dzharullaeva, A.S.; Tukhvatulina, N.M.; Shcheblyakov, D.V.; Shmarov, M.M.; Tokarskaya, E.A.; Simakova, Y.V.; Egorova, D.A.; et al. Safety and immunogenicity of GamEvac-Combi, a heterologous VSV- and Ad5-vectored Ebola vaccine: An open phase I/II trial in healthy adults in Russia. Hum. Vaccines Immunother. 2017, 13, 613–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldmann, H.; Feldmann, F.; Marzi, A. Ebola: Lessons on Vaccine Development. Annu. Rev. Microbiol. 2018, 72, 423–446. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, S.; Banadyga, L.; Bray, M.; Ebihara, H. Small Animal Models for Studying Filovirus Pathogenesis. In Marburg- and Ebolaviruses: From Ecosystems to Molecules; Mühlberger, E., Hensley, L.L., Towner, J.S., Eds.; Springer International: Cham, Switzerland, 2017; pp. 195–227. [Google Scholar]
- Reynolds, P.; Marzi, A. Ebola and Marburg virus vaccines. Virus Genes 2017, 53, 501–515. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Strong, J.E.; Feldmann, H. Considerations in the Use of Nonhuman Primate Models of Ebola Virus and Marburg Virus Infection. J. Infect. Dis. 2015, 212, S91–S97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavot, V. Ebola virus vaccines: Where do we stand? Clin. Immunol. 2016, 173, 44–49. [Google Scholar] [CrossRef]
- Dowall, S.D.; Callan, J.; Zeltina, A.; Al-Abdulla, I.; Strecker, T.; Fehling, S.K.; Krähling, V.; Bosworth, A.; Rayner, E.; Taylor, I.; et al. Development of a Cost-effective Ovine Polyclonal Antibody-Based Product, EBOTAb, to Treat Ebola Virus Infection. J. Infect. Dis. 2016, 213, 1124–1133. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Baranwal, M. Immunoinformatics aided design of peptide-based vaccines against ebolaviruses. Vitam. Horm. 2021, 117, 157–187. [Google Scholar] [CrossRef]
- Fausther-Bovendo, H.; Kobinger, G. The road to effective and accessible antibody therapies against Ebola virus. Curr. Opin. Virol. 2022, 54, 101210. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Daddario-DiCaprio, K.M.; Williams, K.J.; Geisbert, J.B.; Leung, A.; Feldmann, F.; Hensley, L.E.; Feldmann, H.; Jones, S.M. Recombinant vesicular stomatitis virus vector mediates postexposure protection against Sudan Ebola hemorrhagic fever in nonhuman primates. J. Virol. 2008, 82, 5664–5668. [Google Scholar] [CrossRef] [Green Version]
- Medaglini, D.; Siegrist, C.A. Immunomonitoring of human responses to the rVSV-ZEBOV Ebola vaccine. Curr. Opin. Virol. 2017, 23, 88–94. [Google Scholar] [CrossRef]
- Groseth, A.; Feldmann, H.; Strong, J.E. The ecology of Ebola virus. Trends Microbiol. 2007, 15, 408–416. [Google Scholar] [CrossRef]
- Wong, G.; Audet, J.; Fernando, L.; Fausther-Bovendo, H.; Alimonti, J.B.; Kobinger, G.P.; Qiu, X. Immunization with vesicular stomatitis virus vaccine expressing the Ebola glycoprotein provides sustained long-term protection in rodents. Vaccine 2014, 32, 5722–5729. [Google Scholar] [CrossRef]
- Choi, J.H.; Jonsson-Schmunk, K.; Qiu, X.; Shedlock, D.J.; Strong, J.; Xu, J.X.; Michie, K.L.; Audet, J.; Fernando, L.; Myers, M.J.; et al. A Single Dose Respiratory Recombinant Adenovirus-Based Vaccine Provides Long-Term Protection for Non-Human Primates from Lethal Ebola Infection. Mol. Pharm. 2015, 12, 2712–2731. [Google Scholar] [CrossRef] [Green Version]
- Davey, R.T., Jr.; Dodd, L.; Proschan, M.A.; Neaton, J.; Neuhaus Nordwall, J.; Koopmeiners, J.S.; Beigel, J.; Tierney, J.; Lane, H.C.; Fauci, A.S.; et al. A Randomized, Controlled Trial of ZMapp for Ebola Virus Infection. N. Engl. J. Med. 2016, 375, 1448–1456. [Google Scholar] [CrossRef]
- Alizadeh, M.; Amini-Khoei, H.; Tahmasebian, S.; Ghatrehsamani, M.; Ghatreh Samani, K.; Edalatpanah, Y.; Rostampur, S.; Salehi, M.; Ghasemi-Dehnoo, M.; Azadegan-Dehkordi, F.; et al. Designing a novel multi-epitope vaccine against Ebola virus using reverse vaccinology approach. Sci. Rep. 2022, 12, 7757. [Google Scholar] [CrossRef]
- Larivière, Y.; Zola, T.; Stoppie, E.; Maketa, V.; Matangila, J.; Mitashi, P.; De Bie, J.; Muhindo-Mavoko, H.; Van Geertruyden, J.P.; Van Damme, P. Open-label, randomised, clinical trial to evaluate the immunogenicity and safety of a prophylactic vaccination of healthcare providers by administration of a heterologous vaccine regimen against Ebola in the Democratic Republic of the Congo: The study protocol. BMJ Open 2021, 11, e046835. [Google Scholar] [CrossRef]
- Lévy, Y.; Lane, C.; Piot, P.; Beavogui, A.H.; Kieh, M.; Leigh, B.; Doumbia, S.; D’Ortenzio, E.; Lévy-Marchal, C.; Pierson, J.; et al. Prevention of Ebola virus disease through vaccination: Where we are in 2018. Lancet 2018, 392, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, S.B.; Bolay, F.; Kieh, M.; Grandits, G.; Badio, M.; Ballou, R.; Eckes, R.; Feinberg, M.; Follmann, D.; Grund, B.; et al. Phase 2 Placebo-Controlled Trial of Two Vaccines to Prevent Ebola in Liberia. N. Engl. J. Med. 2017, 377, 1438–1447. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Mendoza, S.; Nieto-Gómez, R.; Angulo, C. A Perspective on the Development of Plant-Made Vaccines in the Fight against Ebola Virus. Front. Immunol. 2017, 8, 252. [Google Scholar] [CrossRef] [Green Version]



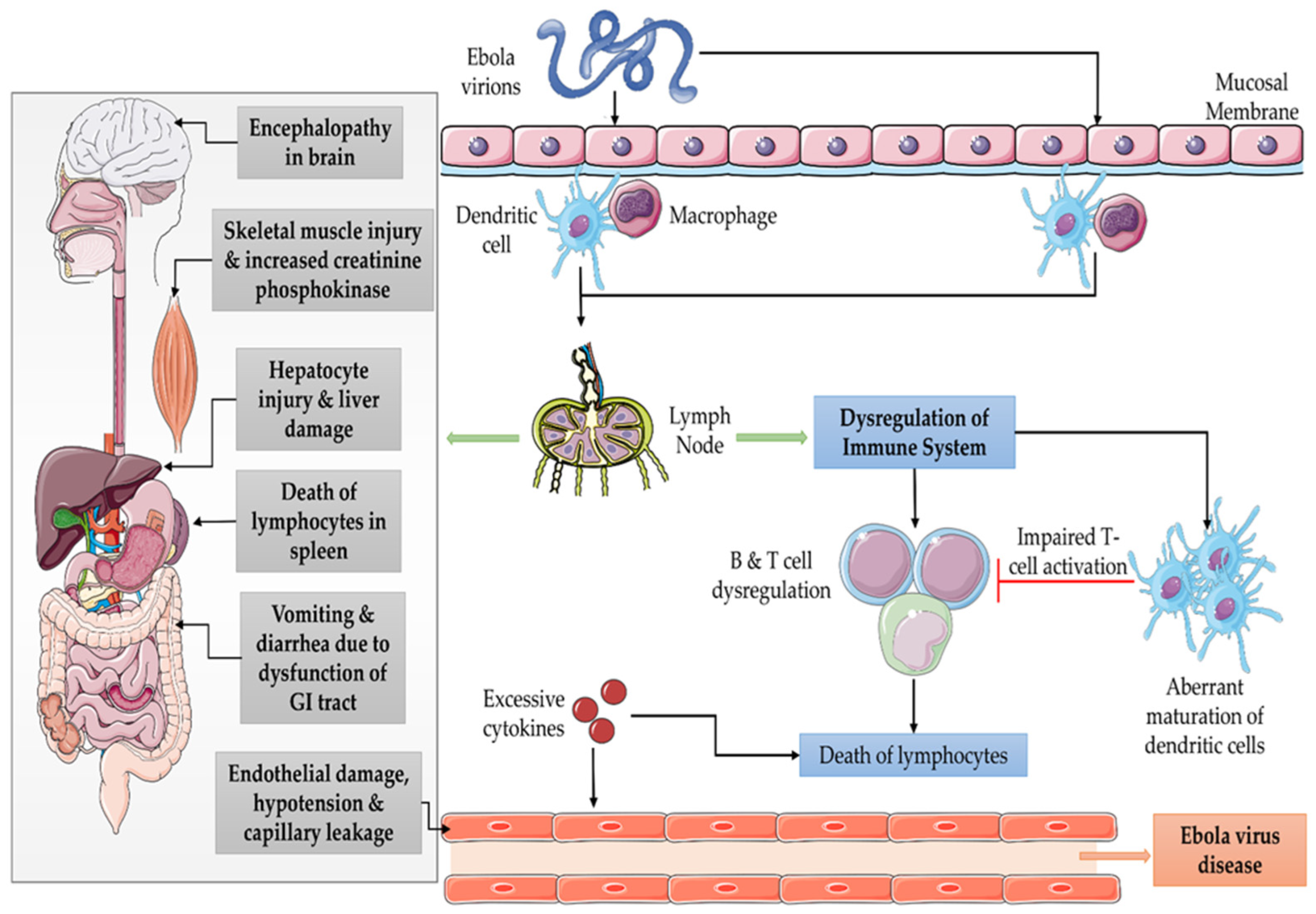{kind=link}
{kind=link}
| Year | Country | EVD | Cases | Deaths | Case Fatality |
|---|---|---|---|---|---|
| 2021 | Guinea | Zaire | Ongoing | ||
| 2021 | Democratic Republic of the Congo | Zaire | Ongoing | ||
| 2020 | Democratic Republic of the Congo | Zaire | 130 | 55 | 42% |
| 2018–2020 | Democratic Republic of the Congo | Zaire | 3481 | 2299 | 66% |
| 2018 | Democratic Republic of the Congo | Zaire | 54 | 33 | 61% |
| 2017 | Democratic Republic of the Congo | Zaire | 8 | 4 | 50% |
| 2015 | Italy | Zaire | 1 | 0 | 0% |
| 2014 | Spain | Zaire | 1 | 0 | 0% |
| 2014 | UK | Zaire | 1 | 0 | 0% |
| 2014 | USA | Zaire | 4 | 1 | 25% |
| 2014 | Senegal | Zaire | 1 | 0 | 0% |
| 2014 | Mali | Zaire | 8 | 6 | 75% |
| 2014 | Nigeria | Zaire | 20 | 8 | 40% |
| 2014–2016 | Sierra Leone | Zaire | 14,124 | 3956 | 28% |
| 2014–2016 | Liberia | Zaire | 10,675 | 4809 | 45% |
| 2014–2016 | Guinea | Zaire | 3811 | 2543 | 67% |
| 2014 | Democratic Republic of the Congo | ||||
| 2012 | Democratic Republic of the Congo | Bundibugyo | 57 | 29 | 51% |
| 2012 | Uganda | Sudan | 7 | 4 | 57% |
| 2012 | Uganda | Sudan | 24 | 17 | 71% |
| 2011 | Uganda | Sudan | 1 | 1 | 100% |
| 2008 | Democratic Republic of the Congo | Zaire | 32 | 14 | 44% |
| 2007 | Uganda | Bundibugyo | 149 | 37 | 25% |
| 2007 | Democratic Republic of the Congo | Zaire | 264 | 187 | 71% |
| 2005 | Congo | Zaire | 12 | 10 | 83% |
| 2004 | Sudan | Sudan | 17 | 7 | 41% |
| 2003 (Nov–Dec) | Congo | Zaire | 35 | 29 | 83% |
| 2003 (Jan–Apr) | Congo | Zaire | 143 | 128 | 90% |
| 2001–2002 | Congo | Zaire | 59 | 44 | 75% |
| 2001–2002 | Gabon | Zaire | 65 | 53 | 82% |
| 2000 | Uganda | Sudan | 425 | 224 | 53% |
| 1996 | South Africa (ex-Gabon) | Zaire | 1 | 1 | 100% |
| 1996 (Jul–Dec) | Gabon | Zaire | 60 | 45 | 75% |
| 1996 (Jan–Apr) | Gabon | Zaire | 31 | 21 | 68% |
| 1995 | Democratic Republic of the Congo | Zaire | 315 | 254 | 81% |
| 1994 | Côte d’Ivoire | Taï Forest | 1 | 0 | 0% |
| 1994 | Gabon | Zaire | 52 | 31 | 60% |
| 1979 | Sudan | Sudan | 34 | 22 | 65% |
| 1977 | Democratic Republic of the Congo | Zaire | 1 | 1 | 100% |
| 1976 | Sudan | Sudan | 284 | 151 | 53% |
| 1976 | Democratic Republic of the Congo | Zaire | 318 | 280 | 88% |
| Vaccine Name | Manufacturer | Type/Category | Other Components | Present Status | Disadvantages and AE | Reference |
|---|---|---|---|---|---|---|
| rVSV-ZEBOV-GP; V920; rVSVAG-ZEBOV-GP (ERVEBO®) | Merck | Live, attenuated vaccine, Recombinant with vesicular stomatitis virus (rVSV). replication-competent | Monovalent, expresses EBOV Glycoprotein (GP) (Kikwit variant) | Approved by US FDA for 18 years and older | Only targets EBOV, which was responsible for the 2013–2016 outbreaks and more recent flareups Reports of arthritis as AE Synovial joints of vaccinated individuals’ reports finding of infectious virus causing secondary spread Stringent storage temperature | [30] |
| Ad26.ZEBOV MVA-BN-Filo boost (Zabdeno/Mvabea) | Johnson & Johnson (Janssen facility)/ Bavarian Nordic | Based on human adenovirus serotype 26 (Ad26) | Multivalent, EBOV GP, TAFV NP, SUDV GP, and MARV GP | Licensed by EMA (exceptional circumstances); Submission g to WHO | Non ideal candidate as per immunogenicity Mvabea vaccine shows lack of immunogenecity against Bundibugyo or Bombali ebolaviruses | [31] |
| Ad5-EBOV | BIT CanSino (China | Recombinant vaccine based on human adenovirus serotype 5 vector (Ad5) | Monovalent, expresses EBOV GP (Makona variant) | Approved by China Food and Drug Administration (CFDA) (2017) based on animal rule | EBOV specific; prior immunity to Ad5 decreases the effectiveness Suitable for 18 to 60 years of age; Lack of clinical data; antibodies decline 85% at day 168 | [32] |
| ChAd3-EBOZ (+/−) Mvabea (cAd3-ZEBOV/ChAd3-EBO-Z) | GlaxoSmithKline, NIAID, Okairos | Recombined with attenuated version of a chimpanzee adenovirus (cAd3) unable to replicate in human,7 | Monovalent, expresses EBOV GP (Mayinga variant)/Mvabea expresses EBOV, SUDV, MARV GP’s and TAFV NP | Not yet licensed by the US FDA or EMA Phase II trials completed in Europe, the US, and Africa | Enhanced vaccines doses for immunogenicity; antibody titre declines by roughly 50% at 180 days after vaccination; booster needed; storage condition issues | [33] |
| GamEvac-Combi and GamEvacLyo | Gamaleya Research Institute of Epidemiology & Microbiology (Russia) | Heterologous primary booster (rVSV and Ad5) expressing EBOV GP | Licensed under Ministry of Health of Russian Federation | prime + booster at 21 days both required; Age limitation from 18 to 55 years age; Un- published safety and efficacy. Preexisting neutralizing Ad5 GP responses in half-dose only | [34] | |
| HPIV3/ΔHNF/EbovZ GP vaccine | NIAID | Live-Attenuated Human Parainfluenza Virus Type 3 Vectored Vaccine | Expressing Ebolavirus Zaire Glycoprotein as the Sole Envelope Glycoprotein | [34] | ||
| Rabies Vector-based GP vaccine | Thomas Jefferson University & NIAID | Recombinant vaccine based on rabies virus (RABV) as vector | Bivalent, Chemically inactivated RABV expressing EBOV GP | Non-human primate (NHP) challenge completed | In early developmental stage | [35] |
| EBOVΔVP30 | University of Wisconsin | Hydrogen peroxide inactivated whole virus | based on a replication-defective EBOV (EBOVΔVP30) | NHP challenge complete | In early developmental stage | [36] |
| Vesiculovax | Auro Vaccines | Attenuated recombinant rVSV vector based | Expression of GP (Mayinga strain of Zaire ebolavirus) | Phase I | In early developmental stage | [37] |
| EBOV DNA Vaccine | NIAID | DNA vaccine | Encodes the envelope GP (Zaire &Sudan species) and the nucleoprotein | Phase I | In early developmental stage | [11] |
| EBOV DNA Vaccine | NIAID | DNA vaccine | Ebola virus (Zaire and Sudan) glycoproteins and (MAR) encoding Marburg virus glycoprotein | Phase 1b | In early developmental stage | [17] |
| Ebola Virus Glycoprotein Nanoparticle Vaccine | US Army Medical Research Institute of Infectious Diseases, Novavax | Recombinant nanoparticle vaccine | EBOV GP nanoconjugated | Phase 1 | In early developmental stage | [38] |
| Ebola Virus | Animal Models | Route of Inoculation | Display of Human Symptoms | Cost of Handling | Ease of Handling | Applications |
|---|---|---|---|---|---|---|
| Wild Type EBOV | Non-human primate | Multiple routes of inoculation | +++ | +++ | +++ | Confirmatory model |
| Ferret | Intranasal | ++ | ++ | ++ | Not well established | |
| Humanized mouse | Intraperitoneal | ++ | +++ | + | Not well established | |
| Guinea Pig adapted EBOV | Guinea Pig | Intraperitoneal, Subcutaneous, Aerosol | + | + | + | Screening model |
| Mouse adapted EBOV | Laboratory mouse | Intraperitoneal, Intranasal, Aerosol | + | + | + | Screening model |
| Hamster | Intraperitoneal | ++ | + | + | Screening model |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, S.; Kishore, S.; Nag, S.; Dhasmana, A.; Preetam, S.; Mitra, O.; León-Figueroa, D.A.; Mohanty, A.; Chattu, V.K.; Assefi, M.; et al. Ebola Virus Disease Vaccines: Development, Current Perspectives & Challenges. Vaccines 2023, 11, 268. https://doi.org/10.3390/vaccines11020268
Malik S, Kishore S, Nag S, Dhasmana A, Preetam S, Mitra O, León-Figueroa DA, Mohanty A, Chattu VK, Assefi M, et al. Ebola Virus Disease Vaccines: Development, Current Perspectives & Challenges. Vaccines. 2023; 11(2):268. https://doi.org/10.3390/vaccines11020268
Chicago/Turabian StyleMalik, Sumira, Shristi Kishore, Sagnik Nag, Archna Dhasmana, Subham Preetam, Oishi Mitra, Darwin A. León-Figueroa, Aroop Mohanty, Vijay Kumar Chattu, Marjan Assefi, and et al. 2023. "Ebola Virus Disease Vaccines: Development, Current Perspectives & Challenges" Vaccines 11, no. 2: 268. https://doi.org/10.3390/vaccines11020268