

Predicting SARS-CoV-2 Variant Spread in a Completely Seropositive Population Using Semi-Quantitative Antibody Measurements in Blood Donors
 , , , , , , , , , , , , add
Show full author list
, , , , , , , , , , , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Blood Donor Sampling Strategy
2.2. SARS-CoV-2 Serology Assay
2.3. Secondary Data Sources
2.4. Data Analysis and Statistics
3. Results
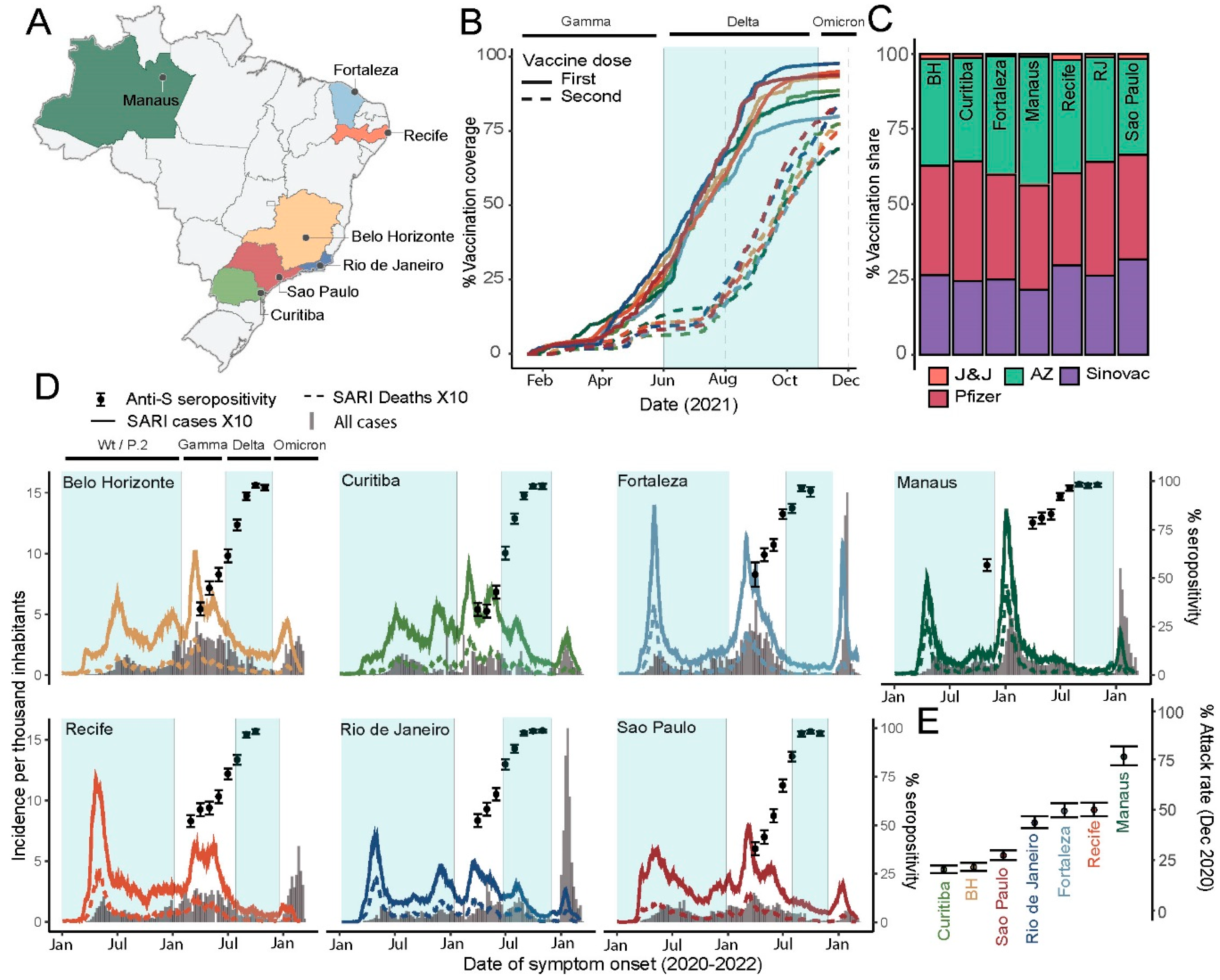
3.1. SARS-CoV-2 Infection, Vaccination and Seroconversion across Seven Brazilian Capitals
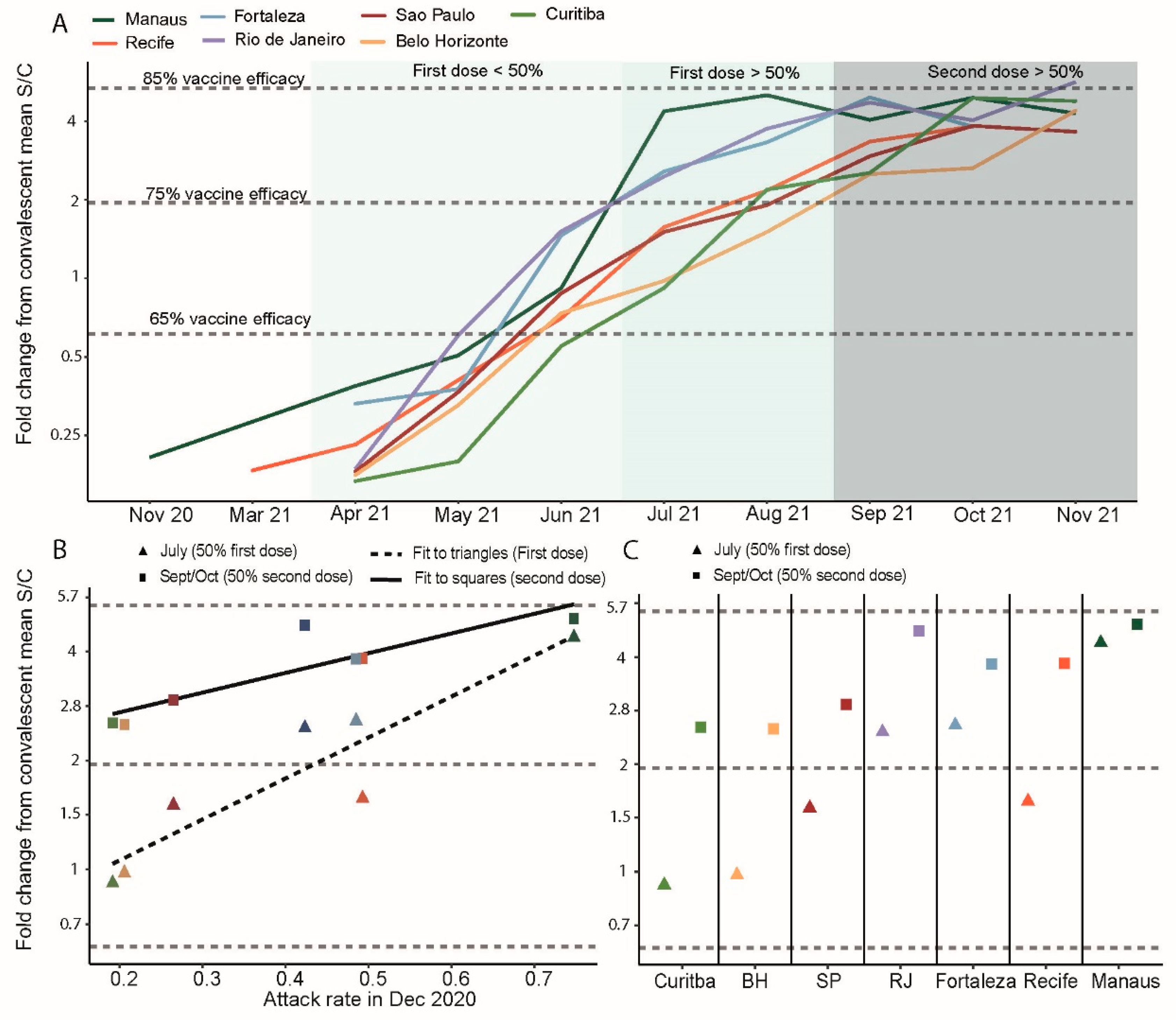
3.2. Convalescent-Normalized Mean S/C in Blood Donors, Vaccination Coverage and Prior Attack Rate
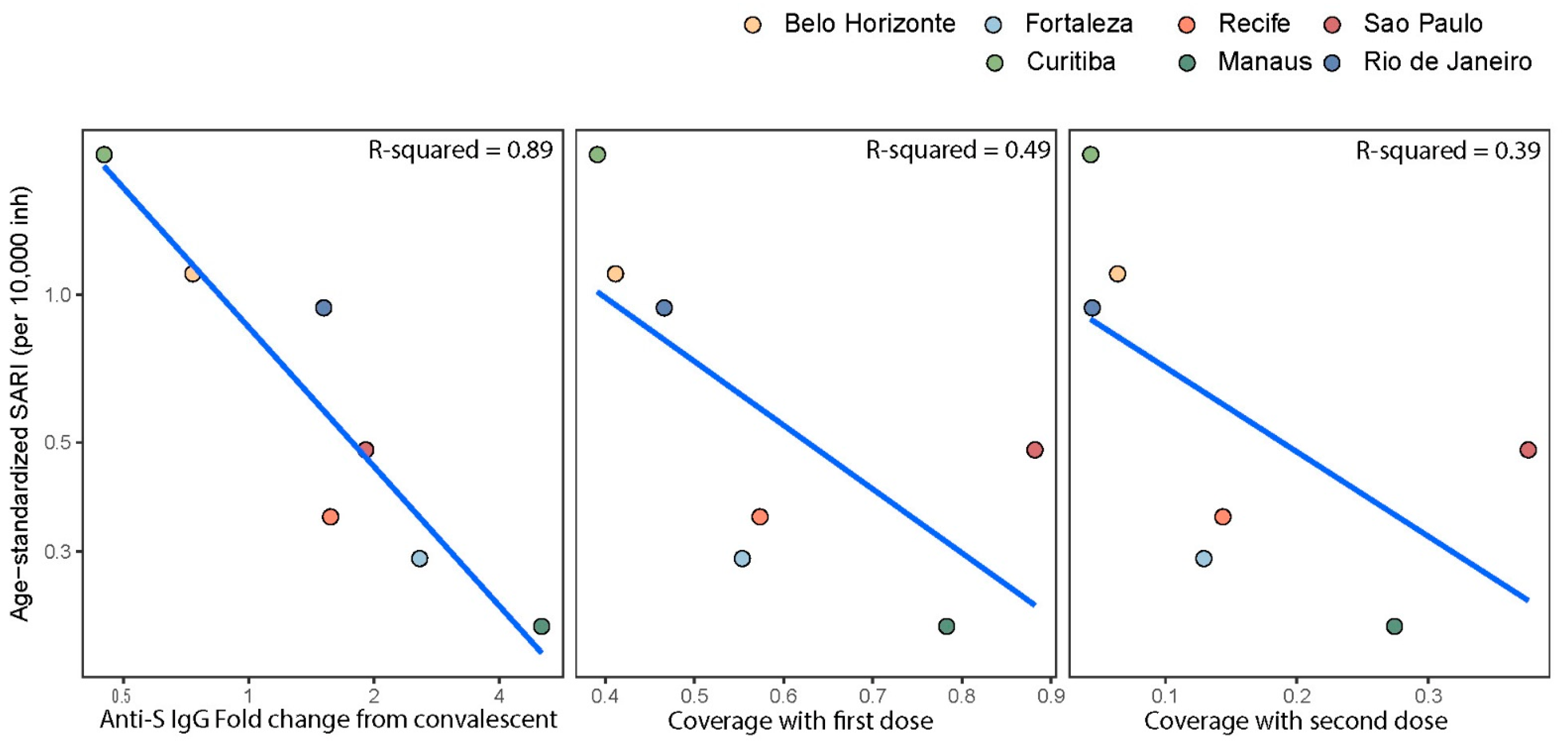
3.3. Anti-S IgG Levels in Blood Donors as Predictor of Delta’s Epidemic Penetrance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, J.M.; Stone, M.; Sulaeman, H.; Fink, R.V.; Dave, H.; Levy, M.E.; Di Germanio, C.; Green, V.; Notari, E.; Saa, P.; et al. Estimated US Infection- and Vaccine-Induced SARS-CoV-2 Seroprevalence Based on Blood Donations. JAMA 2021, 326, 1400. [Google Scholar] [CrossRef] [PubMed]
- Bingham, J.; Cable, R.; Coleman, C.; Glatt, T.N.; Grebe, E.; Mhlanga, L.; Nyano, C.; Pieterson, N.; Swanevelder, R.; Swarts, A.; et al. Estimates of Prevalence of Anti-SARS-CoV-2 Antibodies among Blood Donors in South Africa in March 2022. Available online: https://doi.org/10.21203/rs.3.rs-1687679/v2 (accessed on 30 May 2022).
- CDC. Coronavirus Disease 2019 (COVID-19). 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/variant-classifications.html (accessed on 25 May 2022).
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7. in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.; Vusirikala, A.; Flannagan, J.; Twohig, K.A.; Zaidi, A.; Chudasama, D.; Lamagni, T.; Groves, N.; Turner, C.; Rawlinson, C.; et al. Household transmission of COVID-19 cases associated with SARS-CoV-2 delta variant (B.1.617.2): National case-control study. Lancet Reg. Health Eur. 2022, 12, 100252. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.D.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hofmann-Winkler, H.; Krüger, N.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Arora, P.; Sidarovich, A.; Moldenhauer, A.-S.; Winkler, M.S.; et al. SARS-CoV-2 variant B.1.617 is resistant to bamlanivimab and evades antibodies induced by infection and vaccination. Cell Rep. 2021, 36, 109415. [Google Scholar] [CrossRef] [PubMed]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.S.; Prete, C.A., Jr.; Crispim, M.A.E.; Fraiji, N.A.; Pereira, R.H.M.; Parag, K.V.; da Silva Peixoto, P.; Kraemer, M.U.G.; et al. Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef]
- Buss, L.F.; Prete, C.A., Jr.; Abrahim, C.M.M.; Mendrone, A., Jr.; Salomon, T.; de Almeida-Neto, C.; França, R.F.O.; Belotti, M.C.; Carvalho, M.P.S.S.; Costa, A.G.; et al. Three-quarters attack rate of SARS-CoV-2 in the Brazilian Amazon during a largely unmitigated epidemic. Science 2021, 371, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. Hybrid immunity. Science 2021, 372, 1392–1393. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid epidemic expansion of the SARS-CoV-2 Omicron variant in southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Junior, C.P.; Buss, L.; Salomon, T.; Oikawa, M.; Pereira, R.; Moura, I.; Delerino, L.; Barral-Netto, M.; Tavares, N.; Franca, R.; et al. SARS-CoV-2 Antibody Dynamics in Blood Donors and COVID-19 Epidemiology in Eight Brazilian State Capitals. 2022, in review. Available online: https://www.researchsquare.com/article/rs-1363260/v1 (accessed on 25 May 2022).
- Stone, M.; Grebe, E.; Sulaeman, H.; di Germanio, C.; Dave, H.; Kelly, K.; Biggerstaff, B.J.; Crews, B.O.; Tran, N.; Jerome, K.R.; et al. Evaluation of Commercially Available High-Throughput SARS-CoV-2 Serologic Assays for Serosurveillance and Related Applications. Emerg. Infect. Dis. 2022, 28, 672–683. [Google Scholar] [CrossRef] [PubMed]
- OPENDATASUS. Available online: https://opendatasus.saude.gov.br/ (accessed on 25 May 2022).
- LEPP. DEMOGRAFIA|UFRN. 2017. Available online: https://demografiaufrn.net/laboratorios/lepp/ (accessed on 25 May 2022).
- De Souza, W.M.; Buss, L.F.; Candido, D.d.; Carrera, J.; Li, S.; Zarebski, A.E.; Pereira, R.H.M.; Prete, C.A., Jr.; de Souza-Santos, A.A.; Parag, K.V.; et al. Epidemiological and clinical characteristics of the COVID-19 epidemic in Brazil. Nat. Hum. Behav. 2020, 4, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Brizzi, A.; Whittaker, C.; Servo, L.M.S.; Hawryluk, I.; Prete, C.A., Jr.; de Souza, W.M.; Aguiar, R.S.; Araujo, L.J.T.; Bastos, L.S.; Blenkinsop, A.; et al. Spatial and temporal fluctuations in COVID-19 fatality rates in Brazilian hospitals. Nat. Med. 2022. Available online: https://www.nature.com/articles/s41591-022-01807-1 (accessed on 27 May 2022).
- GISAID—Initiative. Available online: https://www.gisaid.org/ (accessed on 25 May 2022).
- Venables, W.; Ripley, B. Modern Applied Statistics with S., 4th ed.; Springer: New York, NY, USA, 2002; Available online: https://www.stats.ox.ac.uk/pub/MASS4/ (accessed on 30 August 2022).
- Earle, K.A.; Ambrosino, D.M.; Fiore-Gartland, A.; Goldblatt, D.; Gilbert, P.B.; Siber, G.R.; Dull, P.; Plotkin, S.A. Evidence for antibody as a protective correlate for COVID-19 vaccines. Vaccine 2021, 39, 4423–4428. [Google Scholar] [CrossRef] [PubMed]
- Mendrone-Junior, A.; Dinardo, C.L.; Ferreira, S.C.; Nishya, A.; Salles, N.A.; de Almeida Neto, C.; Hamasaki, D.T.; Facincani, T.; de Oliveira Alves, L.B.; Machado, R.R.G.; et al. Correlation between SARS-CoV-2 antibody screening by immunoassay and neutralizing antibody testing. Transfusion 2021, 61, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Otter, A.D.; D’Arcangel, S.; Whitaker, H.; Hewson, J.; Foulkes, S.; Atti, A.; Cole, M.; Linley, E.; Tonge, S.; Hettiarachchi, N.; et al. Determinants of SARS-CoV-2 Anti-Spike Antibody Levels Following BNT162b2 Vaccination: Cross-sectional Analysis of 6000 SIREN Study Participants. Infectious Diseases (Except HIV/AIDS), 2022. Available online: http://medrxiv.org/lookup/doi/10.1101/2022.04.21.22274025 (accessed on 25 May 2022).
- Goldberg, Y.; Mandel, M.; Bar-On, Y.M.; Bodenheimer, O.; Freedman, L.S.; Ash, N.; Alroy-Preis, S.; Huppert, A.; Milo, R. Protection and Waning of Natural and Hybrid Immunity to SARS-CoV-2. N. Engl. J. Med. 2022, 386, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Obermeyer, F.; Jankowiak, M.; Barkas, N.; Schaffner, S.F.; Pyle, J.D.; Yurkovetskiy, L.; Bosso, M.; Park, D.J.; Babadi, M.; MacInnis, B.L.; et al. Analysis of 6.4 million SARS-CoV-2 genomes identifies mutations associated with fitness. Science 2022, 376, 1327–1332. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Model Terms Per Capita × 10 1 | Point Estimate (95% CI) Fold Change form Mean Convalescent | p-Value |
|---|---|---|
| First dose coverage | 1.18 (1.10–1.26) | <0.001 |
| Second dose coverage | 1.15 (1.07–1.24) | <0.001 |
| Attack rate | 1.07 (0.99–1.16) | 0.10 |
| First dose × attack rate | 1.03 (1.01–1.05) | 0.002 |
| Second dose × attack rate | 0.97 (0.95–0.99) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buss, L.; Prete, C.A., Jr.; Whittaker, C.; Salomon, T.; Oikawa, M.K.; Pereira, R.H.M.; Moura, I.C.G.; Delerino, L.; Franca, R.F.O.; Miyajima, F.; et al. Predicting SARS-CoV-2 Variant Spread in a Completely Seropositive Population Using Semi-Quantitative Antibody Measurements in Blood Donors. Vaccines 2022, 10, 1437. https://doi.org/10.3390/vaccines10091437
Buss L, Prete CA Jr., Whittaker C, Salomon T, Oikawa MK, Pereira RHM, Moura ICG, Delerino L, Franca RFO, Miyajima F, et al. Predicting SARS-CoV-2 Variant Spread in a Completely Seropositive Population Using Semi-Quantitative Antibody Measurements in Blood Donors. Vaccines. 2022; 10(9):1437. https://doi.org/10.3390/vaccines10091437
Chicago/Turabian StyleBuss, Lewis, Carlos A. Prete, Jr., Charles Whittaker, Tassila Salomon, Marcio K. Oikawa, Rafael H. M. Pereira, Isabel C. G. Moura, Lucas Delerino, Rafael F. O. Franca, Fabio Miyajima, and et al. 2022. "Predicting SARS-CoV-2 Variant Spread in a Completely Seropositive Population Using Semi-Quantitative Antibody Measurements in Blood Donors" Vaccines 10, no. 9: 1437. https://doi.org/10.3390/vaccines10091437