Inflamma-miR-21 Negatively Regulates Myogenesis during Ageing




Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Satellite Cells Isolation
2.3. Satellite Cell Transfection and Myogenesis
2.4. Isolation of Primary Myoblasts from Mouse Skeletal Muscle
2.5. Cell Culture of Primary Myoblasts
2.6. Transfections of Primary Myoblasts
2.7. Real-Time PCR and Western Blotting
2.8. MTT Assay and Live/Dead Staining of Mouse Myoblasts
2.9. Figure Preparation and Statistics
3. Results
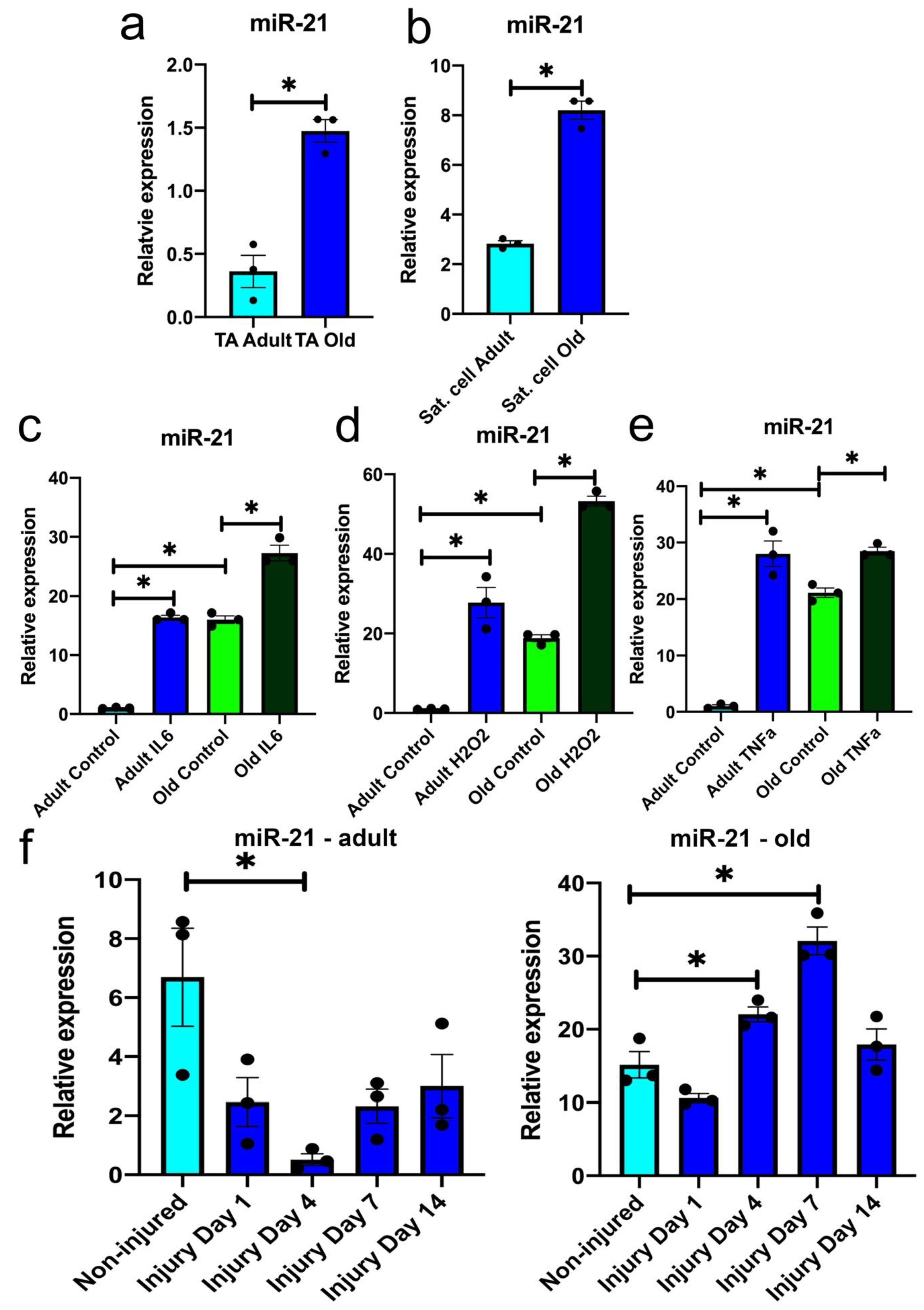
3.1. miR-21 Expression Is Upregulated during Ageing
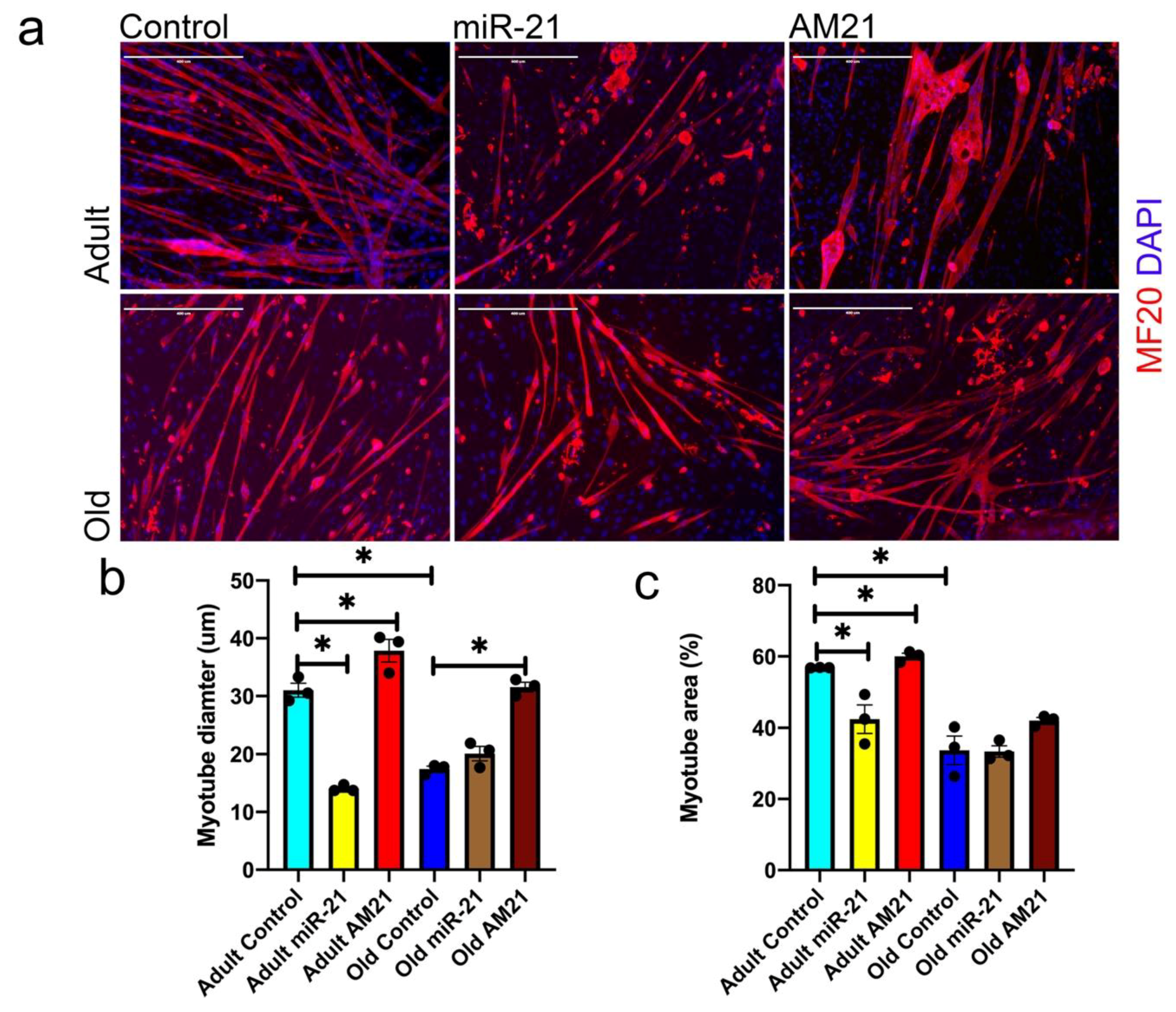
3.2. miR-21 Negatively Regulates Myogenic Potential of Satellite Cells
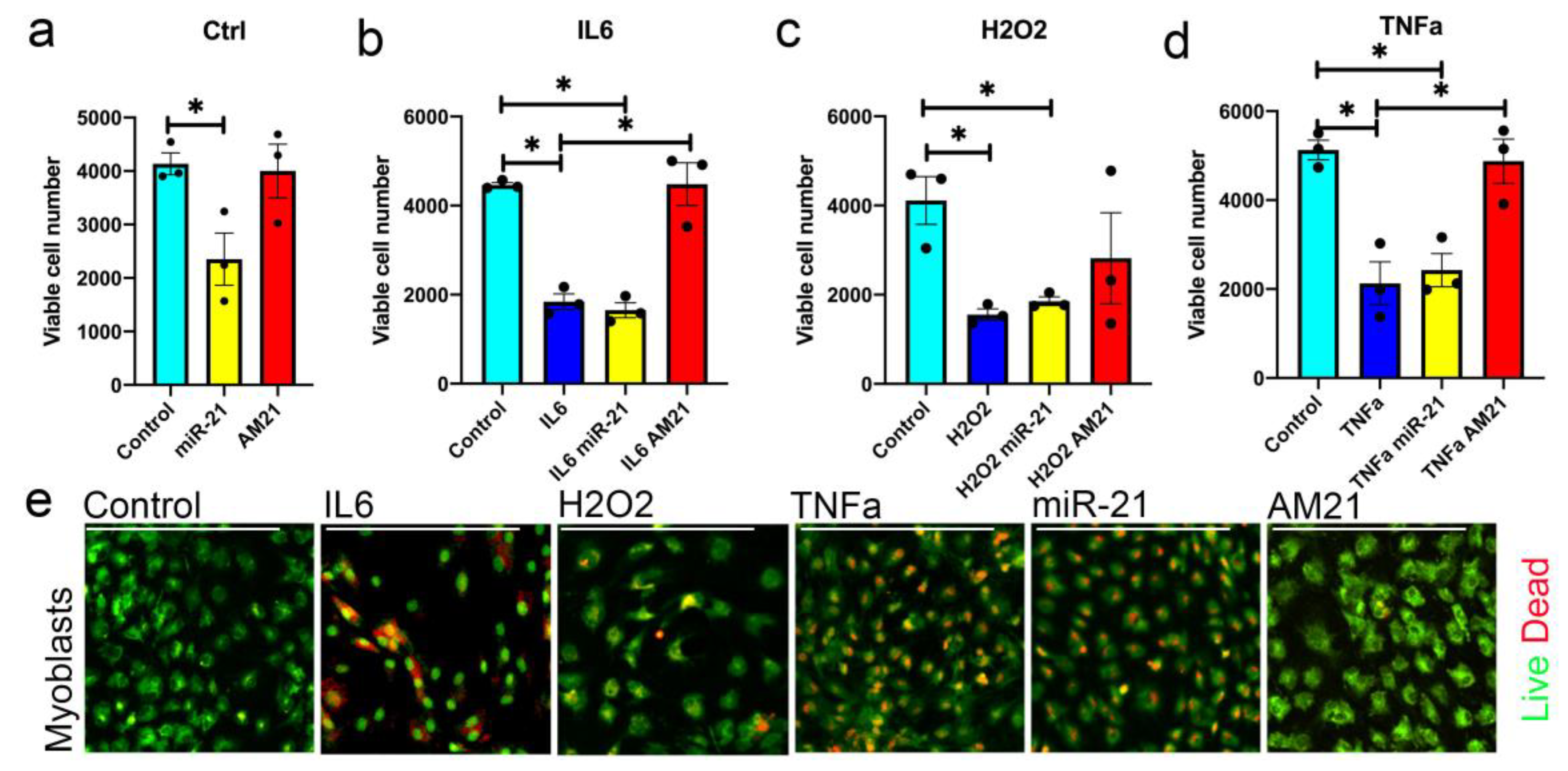
3.3. miR-21 Negatively Regulates Myoblast Viability
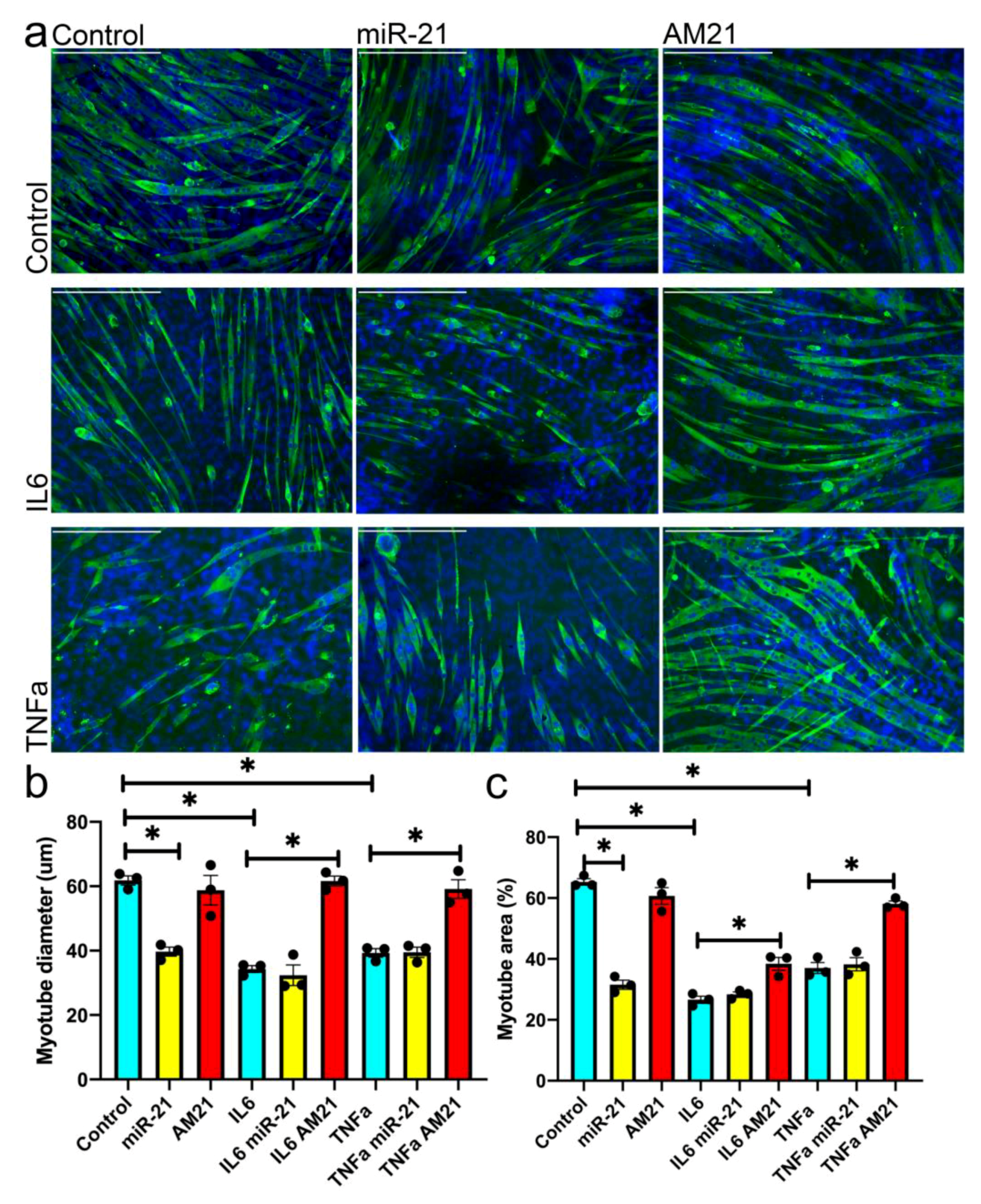
3.4. miR-21 Regulates Myogenesis In Vitro
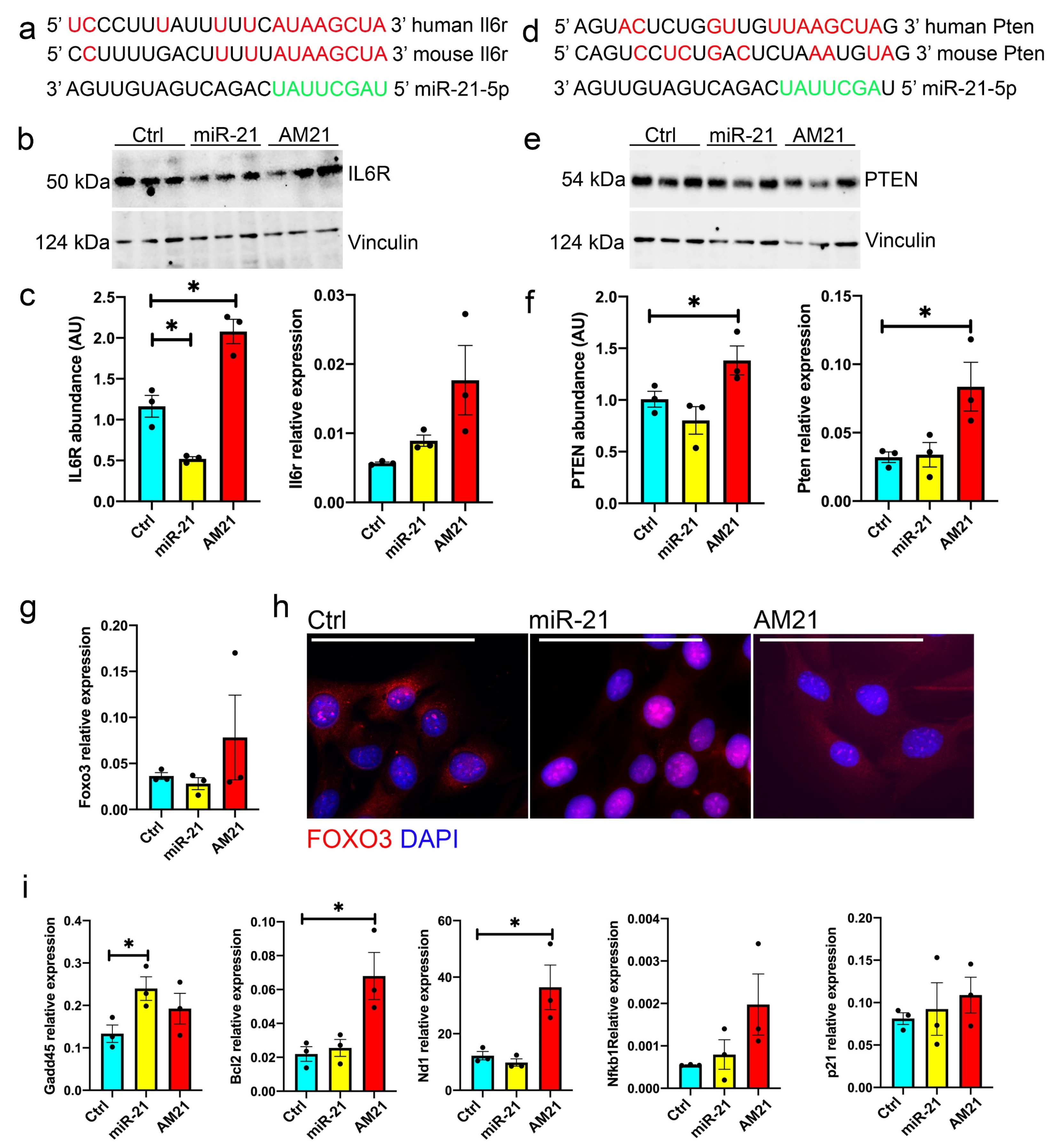
3.5. miR-21 Regulates the Expression of IL6R and PTEN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| DMEM | Sigma Aldrich, Dorset, UK | D5796 |
| Foetal bovine serum | Sigma Aldrich, Dorset UK | F7524 |
| Horse serum | ThermoFisher Scientific, Altrincham, UK | 26050088 |
| BSA | Sigma, Arklow, Ireland | A2153 |
| Glycine | Sigma, Arklow, Ireland | G8898 |
| Triton X | Sigma, Arklow, Ireland | ×100 (lab-grade) or T8787 (for mol biology) |
| DAPI, 1 μg/mL | Sigma Aldrich, Dorset UK | D9542 |
| Bradford assay | BioRad, Deeside, UK | |
| Trypsin | Sigma, Arklow, Ireland | T4049 |
| 2-propanol | Sigma, Arklow, Ireland | I9516 |
| Chloroform:isoamyl alcohol 24:1 | Sigma, Arklow, Ireland | C0549 |
| Hydromount | National Diagnostics, Hull, UK | HS-106 |
| Trizol | ThermoFisher Scientific, Paisley, UK | 15596026 |
| miR-21-5p mimic miRScript | Qiagen, Manchester, UK | 219600 |
| miR-21-5p inhibitor miRScript | Qiagen, Manchester, UK | 219300 |
| Lipofectamine 2000 | Life Technologies, Paisley, UK | 11668027 |
| Mouse IL-6 | Life Technologies, Paisley, UK | PMC0063 |
| Mouse TNFα | Sigma, Gillingham, UK | T7539 |
| Reagent or Resource | Source | Identifier |
|---|---|---|
| ImageJ | Imagej.nih.gov | ImageJ |
| Prism8 | Graphpad | Prism8 |
| Photoshop | Adobe | Photoshop CC release 2017.1.6 |
| AxioVision Software version 4.8 | Zeiss; licence: University of Liverpool | AxioVision Software version 4.8 |
| Reagent or Resource | Source | Identifier |
|---|---|---|
| SuperscriptII | ThermoFisher Scientific, Loughborough, UK | 18064014 |
| miRScript RT II | Qiagen, Manchester, UK | 218161 |
| Qiagent Quanti-Tech | Qiagen, Manchester, UK | Part of 218073 |
| miRScript SybrGreen | Qiagen, Manchester, UK | 218073 |
References
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; Busquets, S.; Stemmler, B.; López-Soriano, F.J. Cachexia and sarcopenia: Mechanisms and potential targets for intervention. Curr. Opin. Pharmacol. 2015, 22, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Della Gatta, P.; Cameron-Smith, D.; Gatta, P.D. Aging and its effects on inflammation in skeletal muscle at rest and following exercise-induced muscle injury. Am. J. Physiol. Integr. Comp. Physiol. 2010, 298, R1485–R1495. [Google Scholar] [CrossRef] [Green Version]
- Hogan, K.A.; Cho, D.S.; Arneson, P.C.; Samani, A.; Palines, P.; Yang, Y.; Doles, J.D. Tumor-derived cytokines impair myogenesis and alter the skeletal muscle immune microenvironment. Cytokine 2018, 107, 9–17. [Google Scholar] [CrossRef]
- Le Moal, E.; Pialoux, V.; Juban, G.; Groussard, C.; Zouhal, H.; Chazaud, B.; Mounier, R. Redox Control of Skeletal Muscle Regeneration. Antioxidants Redox Signal. 2017, 27, 276–310. [Google Scholar] [CrossRef] [PubMed]
- Nelke, C.; Dziewas, R.; Minnerup, J.; Meuth, S.G.; Ruck, T. Skeletal muscle as potential central link between sarcopenia and immune senescence. EBioMedicine 2019, 49, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Perandini, L.; Chimin, P.; Lutkemeyer, D.D.S.; Câmara, N.O.S. Chronic inflammation in skeletal muscle impairs satellite cells function during regeneration: Can physical exercise restore the satellite cell niche? FEBS J. 2018, 285, 1973–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.E.; Partridge, T.A. Muscle satellite cells. Int. J. Biochem. Cell Biol. 2003, 35, 1151–1156. [Google Scholar] [CrossRef]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt Signaling During Aging Alters Muscle Stem Cell Fate and Increases Fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef]
- Collins, C.A.; Zammit, P.S.; Ruiz, A.P.; Morgan, J.; Partridge, T.A. A Population of Myogenic Stem Cells That Survives Skeletal Muscle Aging. STEM CELLS 2007, 25, 885–894. [Google Scholar] [CrossRef]
- Verdijk, L.; Koopman, R.; Schaart, G.; Meijer, K.; Savelberg, H.H.C.M.; Van Loon, L.J.C. Satellite cell content is specifically reduced in type II skeletal muscle fibers in the elderly. Am. J. Physiol. Metab. 2007, 292, E151–E157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano-Arroquia, A.; McCormick, R.; Molloy, A.P.; McArdle, A.; Goljanek-Whysall, K. Age-related changes in miR-143-3p:Igfbp5 interactions affect muscle regeneration. Aging Cell 2016, 15, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arneson, P.C.; Doles, J.D. Impaired Muscle Regeneration in Cancer-Associated Cachexia. Trends Cancer 2019, 5, 579–582. [Google Scholar] [CrossRef]
- Blau, H.M.; Cosgrove, B.D.; Van Ho, A.T. The central role of muscle stem cells in regenerative failure with aging. Nat. Med. 2015, 21, 854–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sannicandro, A.J.; Soriano-Arroquia, A.; Goljanek-Whysall, K. Micro(RNA)-managing muscle wasting. J. Appl. Physiol. 2019, 127, 619–632. [Google Scholar] [CrossRef]
- Sannicandro, A.J.; McDonagh, B.; Goljanek-Whysall, K. MicroRNAs as potential therapeutic targets for muscle wasting during cancer cachexia. Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 157–163. [Google Scholar] [CrossRef]
- Goljanek-Whysall, K.; Iwanejko, L.A.; Vasilaki, A.; Pekovic-Vaughan, V.; McDonagh, B. Ageing in relation to skeletal muscle dysfunction: Redox homoeostasis to regulation of gene expression. Mamm. Genome 2016, 27, 341–357. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, J.J.; Esser, K.A. MicroRNA-1 and microRNA-133a expression are decreased during skeletal muscle hypertrophy. J. Appl. Physiol. 2007, 102, 306–313. [Google Scholar] [CrossRef]
- Goljanek-Whysall, K.; Mok, G.F.; Alrefaei, A.F.; Kennerley, N.; Wheeler, G.N.; Munsterberg, A. myomiR-dependent switching of BAF60 variant incorporation into Brg1 chromatin remodeling complexes during embryo myogenesis. Development 2014, 141, 3378–3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goljanek-Whysall, K.; Pais, H.; Rathjen, T.; Sweetman, D.; Dalmay, T.; Munsterberg, A. Regulation of multiple target genes by miR-1 and miR-206 is pivotal for C2C12 myoblast differentiation. Development 2012, 139, 3590–3600. [Google Scholar] [CrossRef]
- Goljanek-Whysall, K.; Sweetman, D.; Abu-Elmagd, M.; Chapnik, E.; Dalmay, T.; Hornstein, E.; Munsterberg, A. MicroRNA regulation of the paired-box transcription factor Pax3 confers robustness to developmental timing of myogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 11936–11941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Bezprozvannaya, S.; Shelton, J.M.; Frisard, M.I.; Hulver, M.W.; McMillan, R.P.; Wu, Y.; Voelker, K.A.; Grange, R.W.; Richardson, J.A.; et al. Mice lacking microRNA 133a develop dynamin 2–dependent centronuclear myopathy. J. Clin. Investig. 2011, 121, 3258–3268. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.M.; Goljanek-Whysall, K. microRNAs: Modulators of the underlying pathophysiology of sarcopenia? Ageing Res. Rev. 2015, 24, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Goljanek-Whysall, K.; Soriano-Arroquia, A.; McCormick, R.; Chinda, C.A.; McDonagh, B. miR-181a Regulates p62/SQSTM1, Parkin and Protein DJ-1 Promoting Mitochondrial Dynamics in Skeletal Muscle Ageing. Aging Cell 2020. [Google Scholar] [CrossRef] [Green Version]
- Drummond, M.J.; McCarthy, J.J.; Fry, C.S.; Esser, K.A.; Rasmussen, B.B. Aging differentially affects human skeletal muscle microRNA expression at rest and after an anabolic stimulus of resistance exercise and essential amino acids. Am. J. Physiol. Metab. 2008, 295, E1333–E1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Park, Y.-K.; Lee, K.-P.; Lee, S.-M.; Kang, T.-W.; Kim, H.-J.; Dho, S.H.; Kim, S.-Y.; Kwon, K.-S. Genome-wide profiling of the microRNA-mRNA regulatory network in skeletal muscle with aging. Aging 2014, 6, 524–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano-Arroquia, A.; House, L.; Tregilgas, L.; Laird, E.; Goljanek-Whysall, K. The functional consequences of age-related changes in microRNA expression in skeletal muscle. Biogerontology 2016, 17, 641–654. [Google Scholar] [CrossRef] [Green Version]
- He, W.A.; Calore, F.; Londhe, P.; Canella, A.; Guttridge, D.C.; Croce, C.M. Microvesicles containing miRNAs promote muscle cell death in cancer cachexia via TLR7. Proc. Natl. Acad. Sci. USA 2014, 111, 4525–4529. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, I.; Eran, A.; Nishino, I.; Moggio, M.; Lamperti, C.; Amato, A.A.; Lidov, H.G.; Kang, P.B.; North, K.N.; Mitrani-Rosenbaum, S.; et al. Distinctive patterns of microRNA expression in primary muscular disorders. Proc. Natl. Acad. Sci. USA 2007, 104, 17016–17021. [Google Scholar] [CrossRef] [Green Version]
- Cheung, T.H.; Quach, N.L.; Charville, G.W.; Liu, L.; Park, L.; Edalati, A.; Yoo, B.; Hoang, P.; Rando, T.A. Maintenance of muscle stem-cell quiescence by microRNA-489. Nature 2012, 482, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, F.; Spazzafumo, L.; Santini, G.; Lazzarini, R.; Albertini, M.C.; Rippo, M.R.; Galeazzi, R.; Abbatecola, A.M.; Marcheselli, F.; Monti, D.; et al. Age-related differences in the expression of circulating microRNAs: miR-21 as a new circulating marker of inflammaging. Mech. Ageing Dev. 2012, 133, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Rusanova, I.; Diaz-Casado, M.E.; Fernández-Ortiz, M.; Martínez, P.A.; Guerra-Librero, A.; García-García, F.J.; Escames, G.; Mañas, L.; Acuña-Castroviejo, D. Analysis of Plasma MicroRNAs as Predictors and Biomarkers of Aging and Frailty in Humans. Oxidative Med. Cell. Longev. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Hu, B.; Jadhav, R.R.; Jin, J.; Zhang, H.; Cavanagh, M.M.; Akondy, R.S.; Ahmed, R.; Weyand, C.M.; Goronzy, J.J. Activation of miR-21-Regulated Pathways in Immune Aging Selects against Signatures Characteristic of Memory T Cells. Cell Rep. 2018, 25, 2148–2162.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheedy, F. Turning 21: Induction of miR-21 as a Key Switch in the Inflammatory Response. Front. Immunol. 2015, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Medina, P.P.; Nolde, M.; Slack, F.J. OncomiR addiction in an in vivo model of microRNA-21-induced pre-B-cell lymphoma. Nature 2010, 467, 86–90. [Google Scholar] [CrossRef]
- Soares, R.J.R.; Cagnin, S.; Chemello, F.; Silvestrin, M.; Musarò, A.; De Pitta, C.; Lanfranchi, G.; Sandri, M. Involvement of MicroRNAs in the Regulation of Muscle Wasting during Catabolic Conditions*. J. Biol. Chem. 2014, 289, 21909–21925. [Google Scholar] [CrossRef] [Green Version]
- Ardite, E.; Perdiguero, E.; Vidal, B.; Gutarra, S.; Serrano, A.L.; Muñoz-Cánoves, P. PAI-1–regulated miR-21 defines a novel age-associated fibrogenic pathway in muscular dystrophy. J. Cell Biol. 2012, 196, 163–175. [Google Scholar] [CrossRef]
- Bai, L.; Liang, R.; Yang, Y.; Hou, X.; Wang, Z.; Zhu, S.; Wang, C.; Tang, Z.; Li, K. MicroRNA-21 Regulates PI3K/Akt/mTOR Signaling by Targeting TGFβI during Skeletal Muscle Development in Pigs. PLoS ONE 2015, 10, e0119396. [Google Scholar] [CrossRef] [Green Version]
- Soriano-Arroquia, A.; Clegg, P.D.; Molloy, A.P.; Goljanek-Whysall, K. Preparation and Culture of Myogenic Precursor Cells/Primary Myoblasts from Skeletal Muscle of Adult and Aged Humans. J. Vis. Exp. 2017, e55047. [Google Scholar] [CrossRef] [Green Version]
- Brzeszczyńska, J.; Meyer, A.; McGregor, R.; Schilb, A.; Degen, S.; Tadini, V.; Johns, N.; Langen, R.; Schols, A.; Glass, D.J.; et al. Alterations in the in vitro and in vivo regulation of muscle regeneration in healthy ageing and the influence of sarcopenia. J. Cachex-Sarcopenia Muscle 2017, 9, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Van Der Poel, C.; Gosselin, L.E.; Schertzer, J.D.; Ryall, J.; Swiderski, K.; Wondemaghen, M.; Lynch, G.S. Ageing prolongs inflammatory marker expression in regenerating rat skeletal muscles after injury. J. Inflamm. 2011, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Yuan, X.; Xu, A.; Zhu, X.; Zhan, Y.; Wang, S.; Liu, M. Human cancer cells suppress behaviors of endothelial progenitor cells through miR-21 targeting IL6R. Microvasc. Res. 2018, 120, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, J.; Tang, L.; Liao, L.; Xu, Q.; Zhu, S. The Regulation and Function of miR-21-FOXO3a-miR-34b/c Signaling in Breast Cancer. Int. J. Mol. Sci. 2015, 16, 3148–3162. [Google Scholar] [CrossRef] [Green Version]
- Ning, S.-L.; Zhu, H.; Shao, J.; Liu, Y.-C.; Lan, J.; Miao, J. MiR-21 inhibitor improves locomotor function recovery by inhibiting IL-6R/JAK-STAT pathway-mediated inflammation after spinal cord injury in model of rat. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 433–440. [Google Scholar]
- Zhang, J.-G.; Wang, J.-J.; Zhao, F.; Liu, Q.; Jiang, K.; Yang, G.-H. MicroRNA-21 (miR-21) represses tumor suppressor PTEN and promotes growth and invasion in non-small cell lung cancer (NSCLC). Clin. Chim. Acta 2010, 411, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Zegeye, M.M.; Lindkvist, M.; Fälker, K.; Kumawat, A.K.; Paramel, G.; Grenegård, M.; Sirsjö, A.; Ljungberg, L.U. Activation of the JAK/STAT3 and PI3K/AKT pathways are crucial for IL-6 trans-signaling-mediated pro-inflammatory response in human vascular endothelial cells. Cell Commun. Signal. 2018, 16, 55. [Google Scholar] [CrossRef]
- Lo, Y.-T.; Tzeng, T.-F.; Liu, I.-M. Role of Tumor Suppressor PTEN in Tumor Necrosis Factor ?-Induced Inhibition of Insulin Signaling in Murine Skeletal Muscle C 2 C 12 Cells. Horm. Metab. Res. 2007, 39, 173–178. [Google Scholar] [CrossRef]
- Thoma, A.; Lightfoot, A. NF-kB and Inflammatory Cytokine Signalling: Role in Skeletal Muscle Atrophy. Results Probl. Cell Differ. 2018, 1088, 267–279. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.-K. The functional analysis of MicroRNAs involved in NF-κB signaling. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1764–1774. [Google Scholar]
- Ling, M.; Li, Y.; Xu, Y.; Pang, Y.; Shen, L.; Jiang, R.; Zhao, Y.; Yang, X.; Zhang, J.; Zhou, J.; et al. Regulation of miRNA-21 by reactive oxygen species-activated ERK/NF-κB in arsenite-induced cell transformation. Free Radic. Biol. Med. 2012, 52, 1508–1518. [Google Scholar] [CrossRef]
- He, N.; Zhang, Y.L.; Zhang, Y.; Feng, B.; Zheng, Z.; Wang, D.; Zhang, S.; Guo, Q.; Ye, H. Circulating MicroRNAs in Plasma Decrease in Response to Sarcopenia in the Elderly. Front. Genet. 2020, 11, 167. [Google Scholar] [CrossRef] [Green Version]
- Howard, E.E.; Pasiakos, S.M.; Blesso, C.N.; Fussell, M.A.; Rodriguez, N.R. Divergent Roles of Inflammation in Skeletal Muscle Recovery from Injury. Front. Physiol. 2020, 11, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.; Newcomb, P.; Holly, J. Multifaceted roles of TNF-? in myoblast destruction: A multitude of signal transduction pathways. J. Cell. Physiol. 2003, 198, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.-B.; Bao, X.-B.; Ma, Z. Effects of miR-21 on proliferation and apoptosis in human gastric adenocarcinoma cells. Oncol. Lett. 2017, 15, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zhu, K.; Wang, Y.; Yu, H.; Guo, J. Overexpression of miR-21-5p promotes proliferation and invasion of colon adenocarcinoma cells through targeting CHL1. Mol. Med. 2018, 24, 36. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Cheng, M.; Qiao, S.; Wang, Y.; Li, H.; Wang, N. Gga-miR-21 inhibits chicken pre-adipocyte proliferation in part by down-regulating Kruppel-like factor 5. Poult. Sci. 2017, 96, 200–210. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Yu, H.-N.; Noh, E.-M.; Youn, H.J.; Song, E.-K.; Han, M.-K.; Park, C.-S.; Kim, B.-S.; Park, Y.-S.; Park, B.K.; et al. TNF-alpha upregulates PTEN via NF-kappaB signaling pathways in human leukemic cells. Exp. Mol. Med. 2007, 39, 121–127. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Hu, Z.; Wang, X.; Sheng, H.; Zhang, L.; Du, J.; Mitch, W.E.; Lee, I.H. PTEN Expression Contributes to the Regulation of Muscle Protein Degradation in Diabetes. Diabetes 2007, 56, 2449–2456. [Google Scholar] [CrossRef] [Green Version]
- Yue, F.; Bi, P.; Wang, C.; Li, J.; Liu, X.; Kuang, S. Conditional Loss of Pten in Myogenic Progenitors Leads to Postnatal Skeletal Muscle Hypertrophy but Age-Dependent Exhaustion of Satellite Cells. Cell Rep. 2016, 17, 2340–2353. [Google Scholar] [CrossRef] [Green Version]
- Yue, F.; Bi, P.; Wang, C.; Shan, T.; Nie, Y.; Ratliff, T.L.; Gavin, T.P.; Kuang, S. Pten is necessary for the quiescence and maintenance of adult muscle stem cells. Nat. Commun. 2017, 8, 14328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, T.S.; Schuler, G.; Adams, V. Skeletal muscle wasting in cachexia and sarcopenia: Molecular pathophysiology and impact of exercise training. J. Cachexia Sarcopenia Muscle 2015, 6, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo Transcription Factors Induce the Atrophy-Related Ubiquitin Ligase Atrogin-1 and Cause Skeletal Muscle Atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Fasano, C.; Disciglio, V.; Bertora, S.; Signorile, M.L.; Simone, C. FOXO3a from the Nucleus to the Mitochondria: A Round Trip in Cellular Stress Response. Cells 2019, 8, 1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, S.M.; Dyle, M.C.; Kunkel, S.D.; Bullard, S.A.; Bongers, K.S.; Fox, D.K.; Dierdorff, J.M.; Foster, E.D.; Adams, C.M. Stress-induced Skeletal Muscle Gadd45a Expression Reprograms Myonuclei and Causes Muscle Atrophy*. J. Biol. Chem. 2012, 287, 27290–27301. [Google Scholar] [CrossRef] [Green Version]
- Bullard, S.A.; Seo, S.; Schilling, B.; Dyle, M.C.; Dierdorff, J.M.; Ebert, S.M.; Delau, A.D.; Gibson, B.W.; Adams, C.M. Gadd45a Protein Promotes Skeletal Muscle Atrophy by Forming a Complex with the Protein Kinase MEKK4*♦. J. Biol. Chem. 2016, 291, 17496–17509. [Google Scholar] [CrossRef] [Green Version]
- Belizário, J.; Fontes-Oliveira, C.C.; Borges, J.P.; Kashiabara, J.A.; Vannier, E. Skeletal muscle wasting and renewal: A pivotal role of myokine IL-6. SpringerPlus 2016, 5, 619. [Google Scholar] [CrossRef] [Green Version]
- Tamura, Y.; Phan, C.; Tu, L.; Le Hiress, M.; Thuillet, R.; Jutant, E.; Fadel, E.; Savale, L.; Huertas, A.; Humbert, M.; et al. Ectopic upregulation of membrane-bound IL6R drives vascular remodeling in pulmonary arterial hypertension. J. Clin. Investig. 2018, 128, 1956–1970. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Shi, Y.; Tan, G.; Yang, C.H.; Fan, M.; Pfeffer, L.M.; Wu, Z.H. DNA damage induces NF-κB-dependent microRNA-21 up-regulation and promotes breast cancer cell invasion. J. Biol. Chem. 2019, 22, 18015. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Q.; Li, X.L.; Wang, L.; Du, W.J.; Guo, R.; Liang, H.H.; Liu, X.; Liang, D.S.; Lu, Y.J.; Shan, H.L.; et al. Matrine inhibits breast cancer growth via miR-21/PTEN/Akt pathway in MCF-7 cells. Cell Physiol. Biochem. 2012, 30, 631–641. [Google Scholar] [CrossRef]





| Gene Name | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| β2-microglobulin | GGAGAATGGGAAGCCGAACA | TCTCGATCCCAGTAGACGGT |
| S29 | GGCAGTACGCGAAGGACATA | CAAGGTCGCTTAGTCCAACTTA |
| S18 | CGGCTACCACATCCAAGGAAGG | CCCGCTCCCAAGATCCAACTAC |
| Foxo-3 | AGTGGATGGTGCGCTGTGT | CTGTGCAGGGACAGGTTGT |
| IL-6r | CTTGGATAGCAGAGCCCAGG | CTCGTGGTTGGCAGAGTCTT |
| Pten | TTGGCGGTGTCATAATGTCT | GCAGAAAGACTTGAAGGCGTA |
| Nfkb1 | ACACGAGGCTACAACTCTGC | GGTACCCCCAGAGACCTCAT |
| P21 | GGCAGACCAGCATGACAGATTTC | CGGATTAGGGCTTCCTCTTGG |
| Nd-1 | CCTATCACCCTTGCCATCAT | GAGGCTGTTGCTTGTGTGAC |
| Gadd45 | CTGTGTGCTGGTGACGAACC | TCCATGTAGCGACTTTCCCG |
| miR-21 | Cat. 218300 | Universal Primer, part of cat. 218073 |
| Rnu-6 | Cat. MS00033740 | Universal Primer, part of cat. 218073 |
| Snord-61 | Cat. MS00033705 | Universal Primer, part of cat. 218073 |
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Foxo3 | Cell Signalling | 2497 |
| Phospho-Foxo3 | Cell Signalling | 9465 |
| Pten | Cell Signaling Technology, Boston, USA | 9188S |
| IL-6R | Abcam, Cambridge, UK | ab83054 |
| Vinculin | Abcam, Cambridge, UK | ab18058 |
| NF-kB | Abcam, Cambridge, UK | ab32360 |
| MF20 | Developmental Studies Hybridoma Bank, Iowa, USA | MF20 supernatant |
| IRDye 800CW Goat anti-Rabbit IgG | Li-Cor Biosciences, Cambridge, UK | 926-32211 |
| IRDye 800CW Goat anti-Mouse IgG | Li-Cor Biosciences, Cambridge, UK | 926-68020 |
| anti-mouse –Alexa488 | ThermoFisher Scientific, Altrincham, UK | A28175 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borja-Gonzalez, M.; Casas-Martinez, J.C.; McDonagh, B.; Goljanek-Whysall, K. Inflamma-miR-21 Negatively Regulates Myogenesis during Ageing. Antioxidants 2020, 9, 345. https://doi.org/10.3390/antiox9040345
Borja-Gonzalez M, Casas-Martinez JC, McDonagh B, Goljanek-Whysall K. Inflamma-miR-21 Negatively Regulates Myogenesis during Ageing. Antioxidants. 2020; 9(4):345. https://doi.org/10.3390/antiox9040345
Chicago/Turabian StyleBorja-Gonzalez, Maria, Jose C. Casas-Martinez, Brian McDonagh, and Katarzyna Goljanek-Whysall. 2020. "Inflamma-miR-21 Negatively Regulates Myogenesis during Ageing" Antioxidants 9, no. 4: 345. https://doi.org/10.3390/antiox9040345