Nasally Administered Lactococcus lactis Secreting Heme Oxygenase-1 Attenuates Murine Emphysema

 ,
,  , and
, and 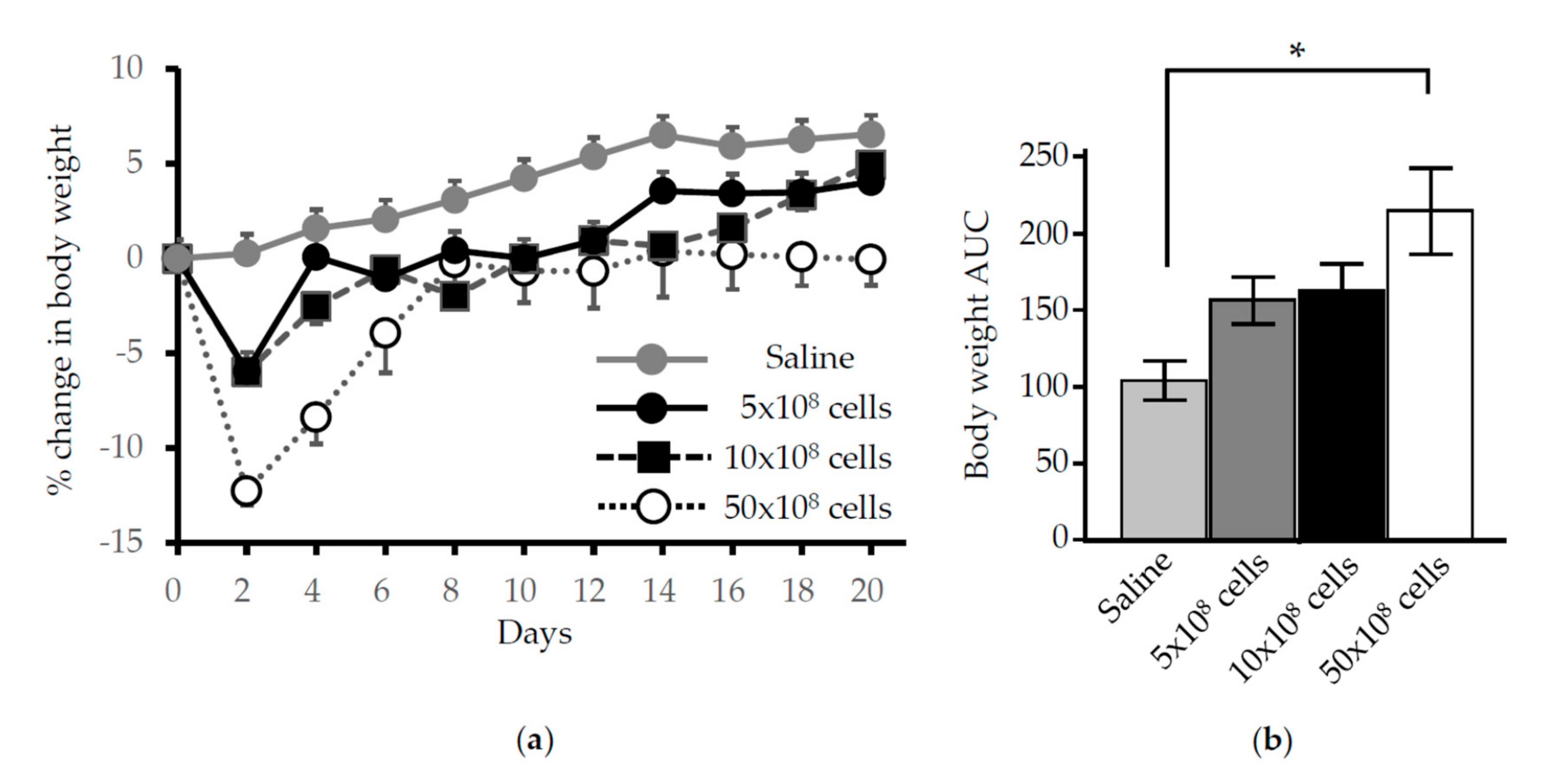{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Murine Emphysema Model
2.3. Bacterial Culture and Gene Expression
2.4. Administration of L. lactis
2.5. Bronchoalveolar Lavage
2.6. Western Blotting Assay
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Immunohistochemical Detection
2.9. Morphologic Evaluation and Quantification of Emphysema
2.10. Lung Function Measurements
2.11. Statistical Analysis
3. Results
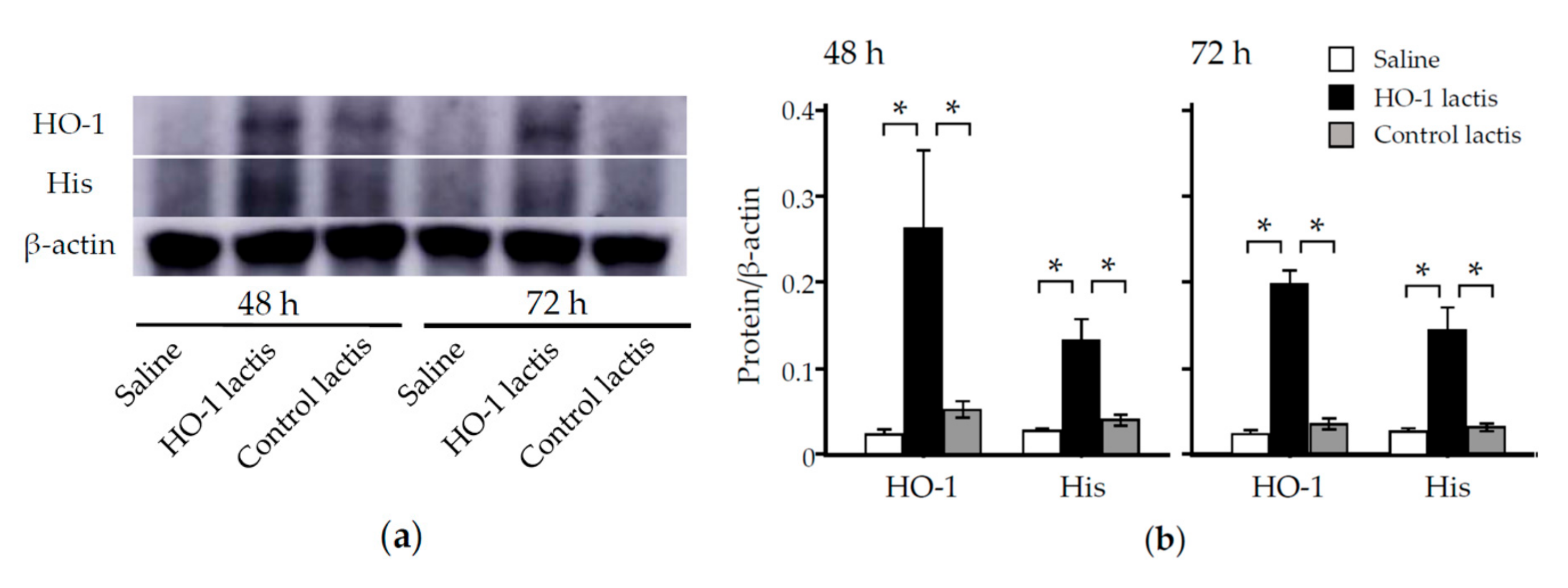
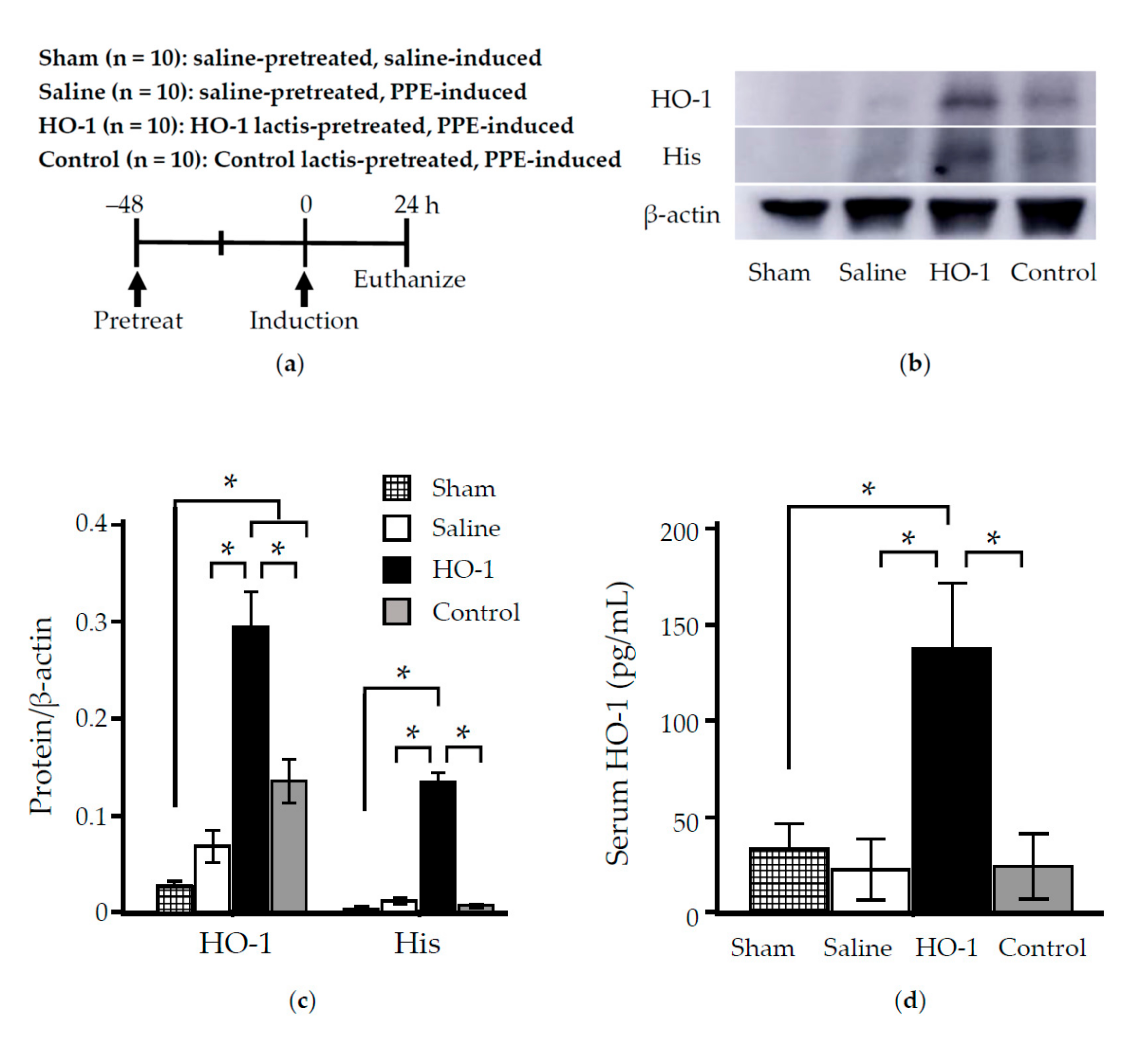
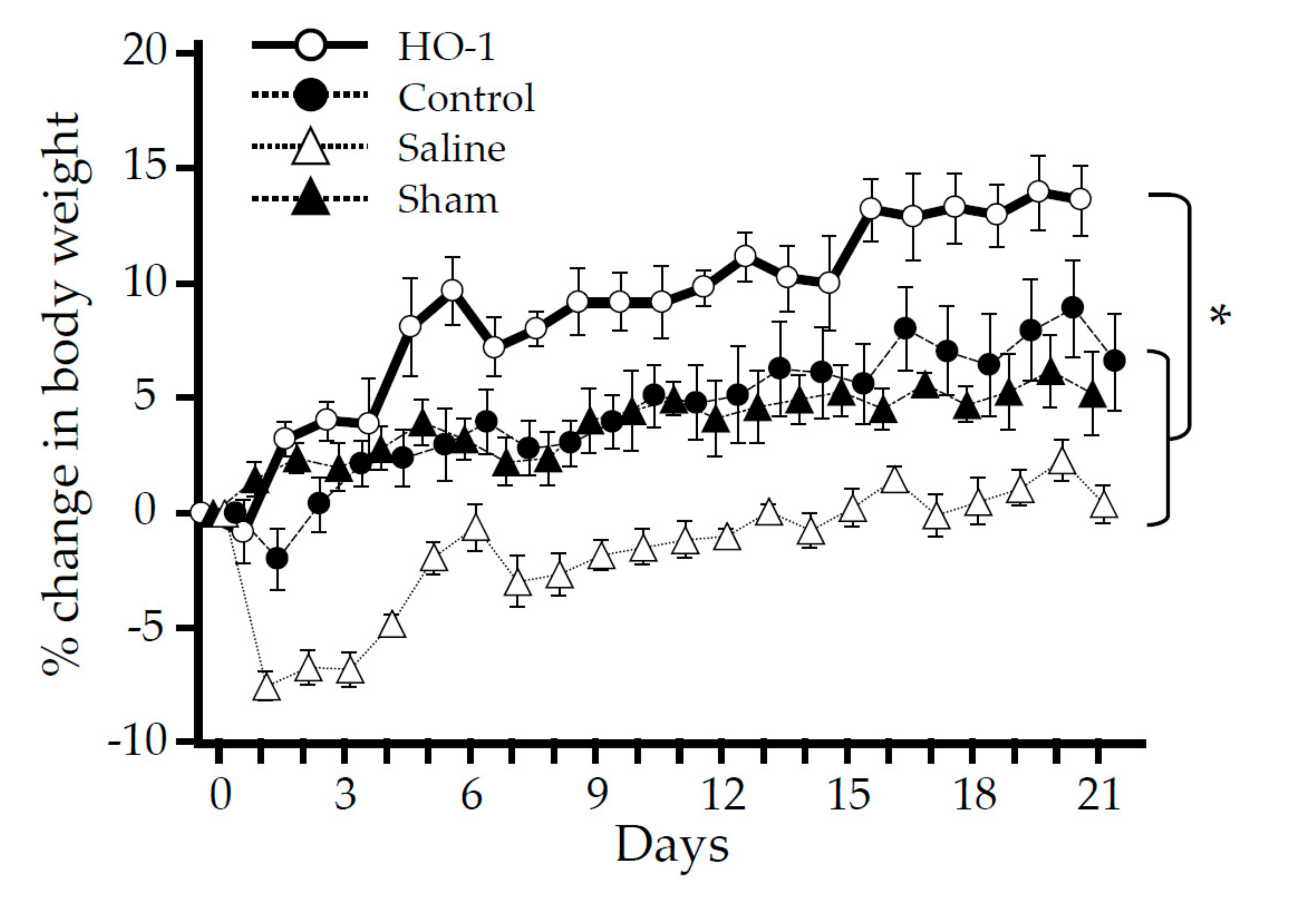
3.1. Time Course Analysis of Systemic and Local Effects of HO-1 Lactis Administration
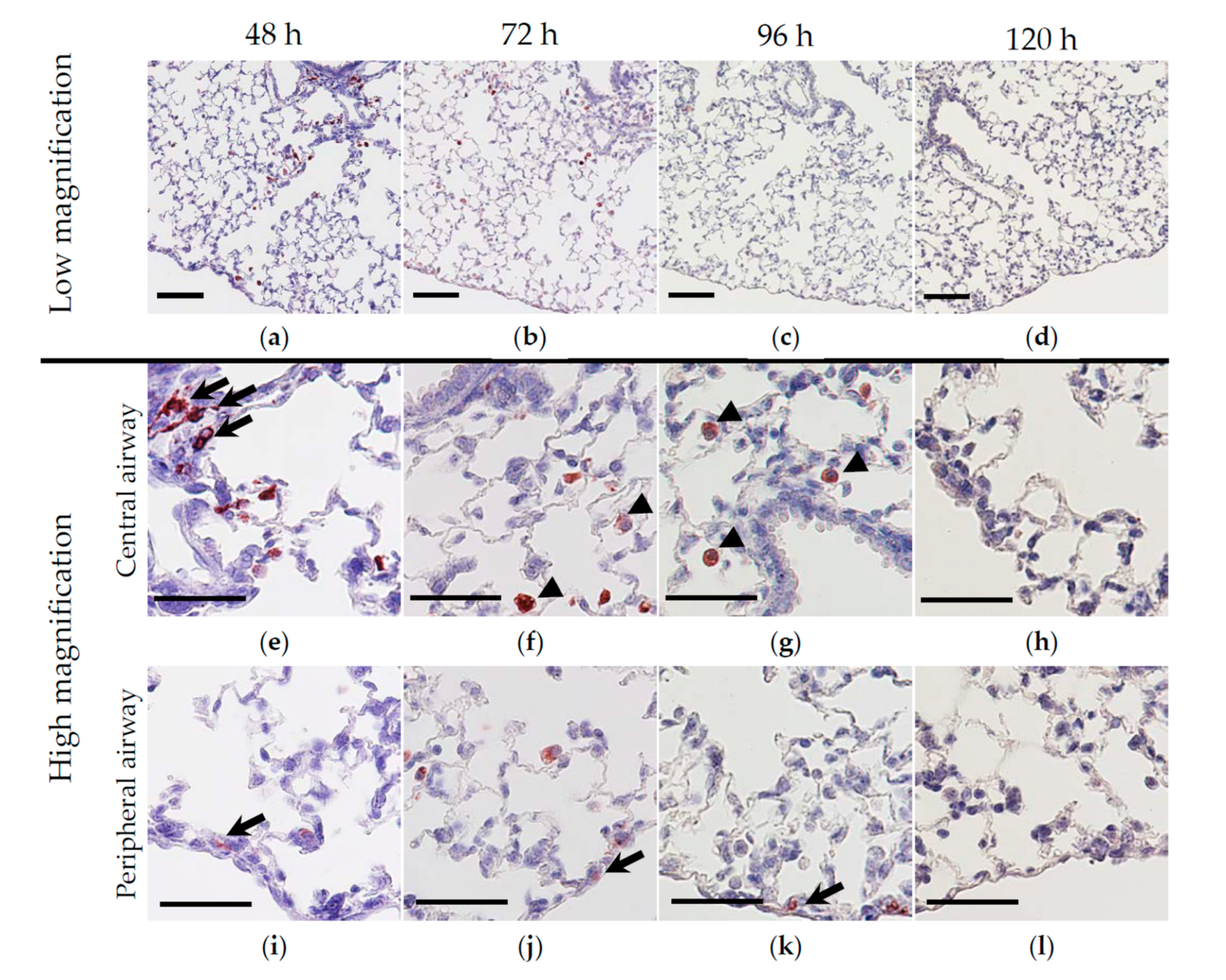
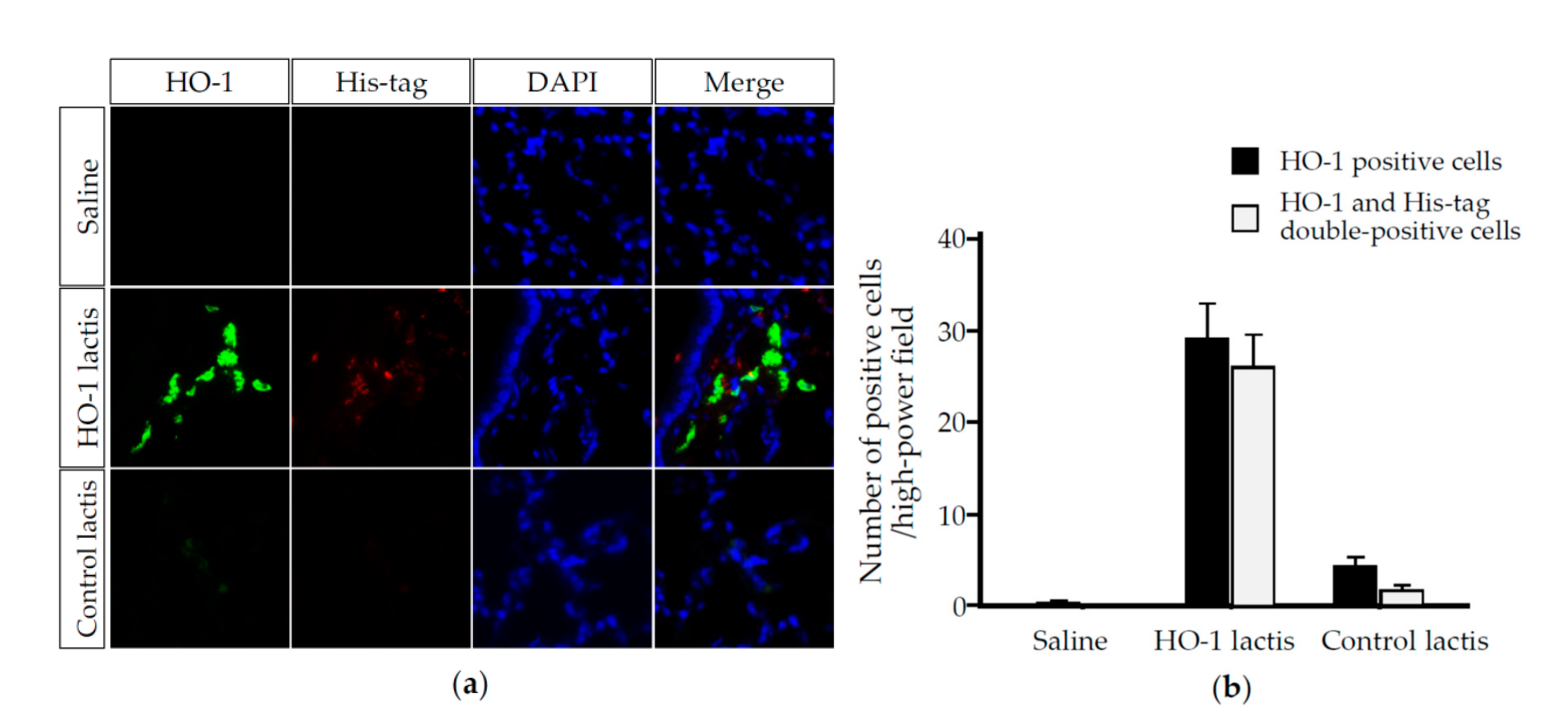
3.2. Nasally Administered HO-1 Lactis Cells Reach the Lung in PPE-Induced Emphysema
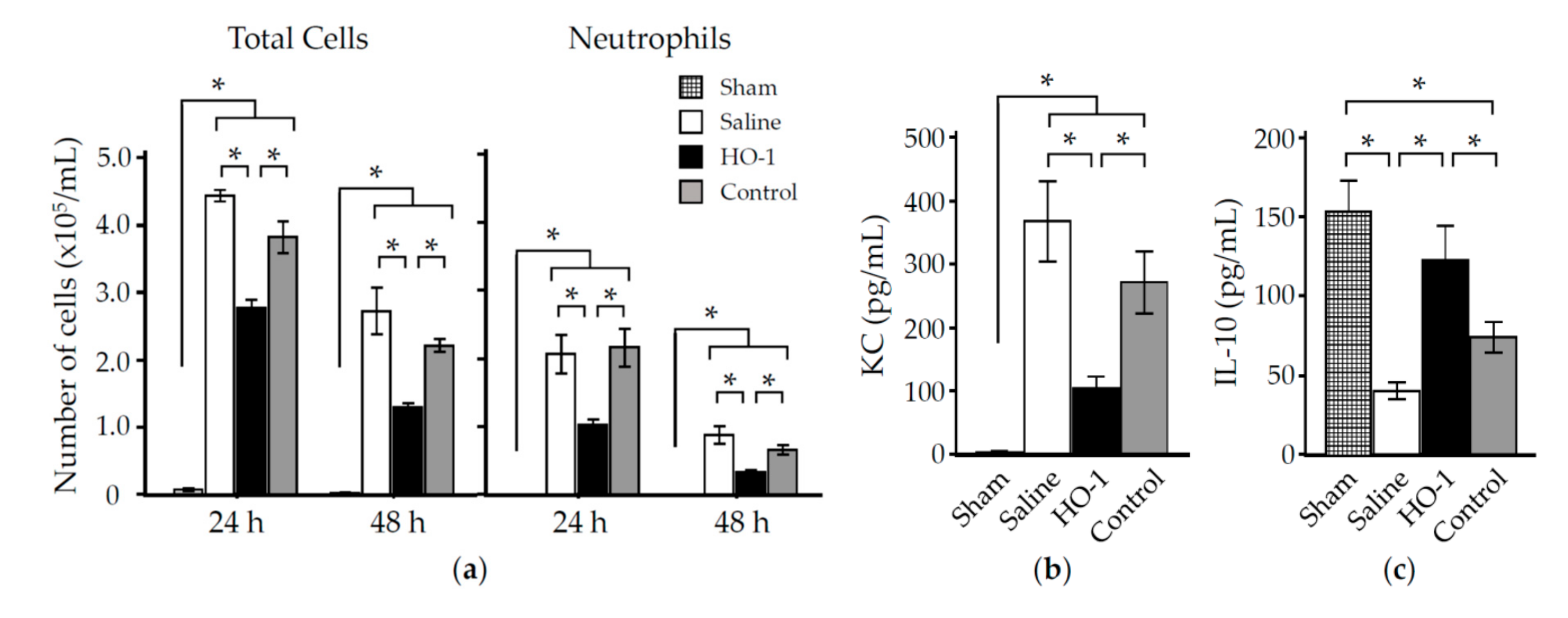
3.3. Nasal Administration of HO-1 Lactis Reduces PPE-Induced Lung Inflammation
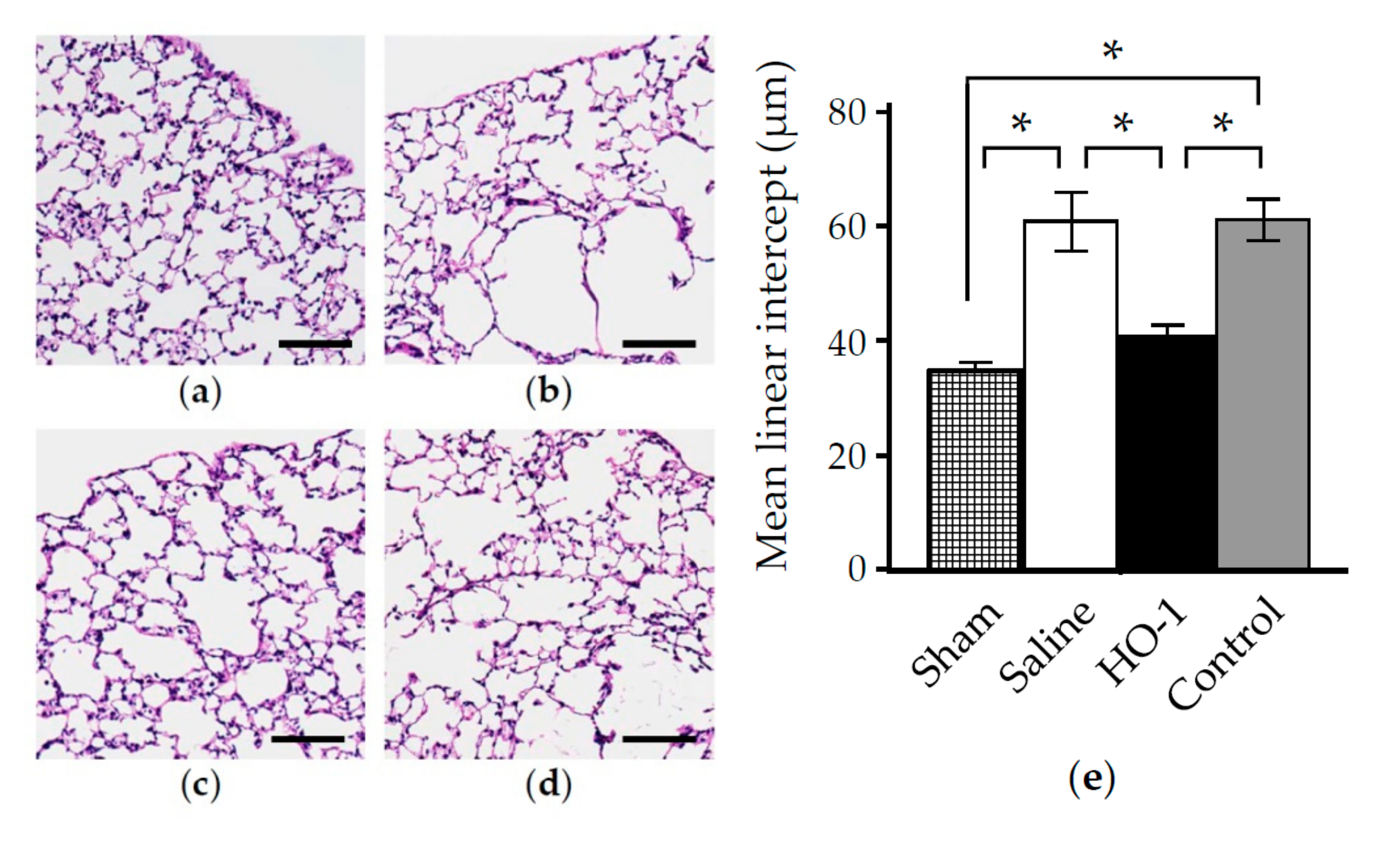
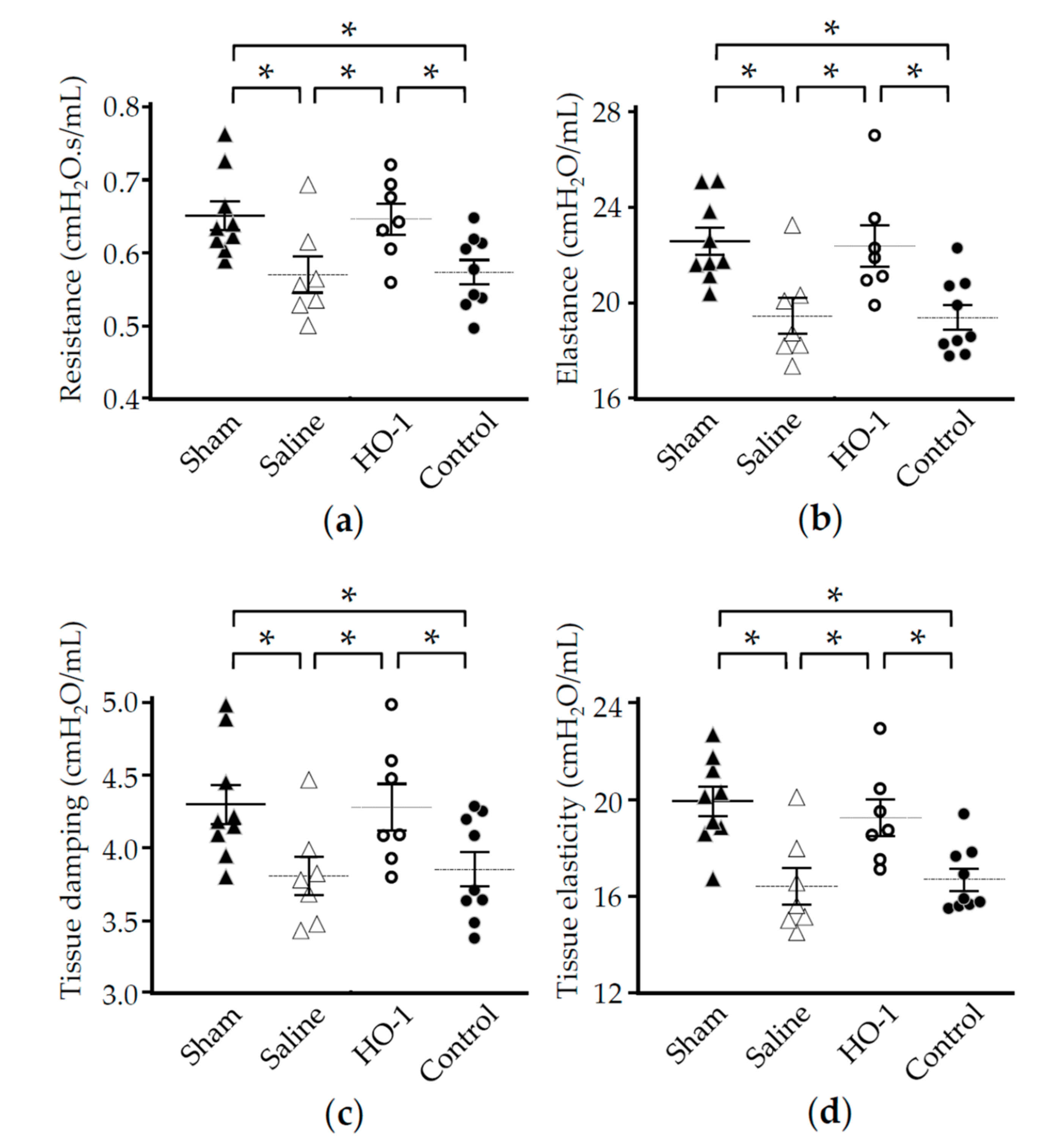
3.4. Nasal Administration of HO-1 Lactis Reduces the Physio-Pathologic Deterioration Induced by PPE
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- GBD, Chronic Respiratory Disease Collaborators. Global, regional, and national deaths, prevalence, disability-adjusted life years, and years lived with disability for chronic obstructive pulmonary disease and asthma, 1990-2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Respir. Med. 2017, 5, 691–706. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 13 July 2020).
- Chinai, B.; Hunter, K.; Roy, S. Outpatient Management of Chronic Obstructive Pulmonary Disease: Physician Adherence to the 2017 Global Initiative for Chronic Obstructive Lung Disease Guidelines and its Effect on Patient Outcomes. J. Clin. Med. Res. 2019, 11, 556–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sopori, M. Effects of cigarette smoke on the immune system. Nat. Rev. Immunol. 2002, 2, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Terzikhan, N.; Verhamme, K.M.; Hofman, A.; Stricker, B.H.; Brusselle, G.G.; Lahousse, L. Prevalence and incidence of COPD in smokers and non-smokers: The Rotterdam Study. Eur. J. Epidemiol. 2016, 31, 785–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, J.M.; Scott, A.L.; Mitzner, W. Immune-mediated inflammation in the pathogenesis of emphysema: Insights from mouse models. Cell Tissue Res. 2017, 367, 591–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldkorn, T.; Filosto, S.; Chung, S. Lung injury and lung cancer caused by cigarette smoke-induced oxidative stress: Molecular mechanisms and therapeutic opportunities involving the ceramide-generating machinery and epidermal growth factor receptor. Antioxid. Redox. Signal. 2014, 21, 2149–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houghton, A.M. Matrix metalloproteinases in destructive lung disease. Matrix Biol. 2015, 44, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Sharafkhaneh, A.; Hanania, N.A.; Kim, V. Pathogenesis of emphysema: From the bench to the bedside. Proc. Am. Thorac. Soc. 2008, 5, 475–477. [Google Scholar] [CrossRef] [PubMed]
- Grabiec, A.M.; Hussell, T. The role of airway macrophages in apoptotic cell clearance following acute and chronic lung inflammation. Semin. Immunopathol. 2016, 38, 409–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repine, J.E.; Bast, A.; Lankhorst, I. Oxidative stress in chronic obstructive pulmonary disease. Oxidative Stress Study Group. Am. J. Respir. Crit. Care Med. 1997, 156, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Tenhunen, R.; Marver, H.S.; Schmid, R. The enzymatic conversion of heme to bilirubin by microsomal heme oxygenase. Proc. Natl. Acad. Sci. USA 1968, 61, 748–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarady-Andrews, J.K.; Liu, F.; Gallo, D.; Nakao, A.; Overhaus, M.; Ollinger, R.; Choi, A.M.; Otterbein, L.E. Biliverdin administration protects against endotoxin-induced acute lung injury in rats. Am. J. Physiol. Lung. Cell Mol. Physiol. 2005, 289, L1131–L1137. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Bach, F.H.; Alam, J.; Soares, M.; Tao Lu, H.; Wysk, M.; Davis, R.J.; Flavell, R.A.; Choi, A.M. Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat. Med. 2000, 6, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Sarady, J.K.; Otterbein, S.L.; Liu, F.; Otterbein, L.E.; Choi, A.M. Carbon monoxide modulates endotoxin-induced production of granulocyte macrophage colony-stimulating factor in macrophages. Am. J. Respir. Cell Mol. Biol. 2002, 27, 739–745. [Google Scholar] [CrossRef]
- Balla, G.; Jacob, H.S.; Balla, J.; Rosenberg, M.; Nath, K.; Apple, F.; Eaton, J.W.; Vercellotti, G.M. Ferritin: A cytoprotective antioxidant strategem of endothelium. J. Biol. Chem. 1992, 267, 18148–18153. [Google Scholar]
- Fredenburgh, L.E.; Perrella, M.A.; Mitsialis, S.A. The role of heme oxygenase-1 in pulmonary disease. Am. J. Respir. Cell Mol. Biol. 2007, 36, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Poss, K.D.; Tonegawa, S. Heme oxygenase 1 is required for mammalian iron reutilization. Proc. Natl. Acad. Sci. USA 1997, 94, 10919–10924. [Google Scholar] [CrossRef] [Green Version]
- Otterbein, L.E.; Kolls, J.K.; Mantell, L.L.; Cook, J.L.; Alam, J.; Choi, A.M. Exogenous administration of heme oxygenase-1 by gene transfer provides protection against hyperoxia-induced lung injury. J. Clin. Investig. 1999, 103, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Takeno, M.; Honma, K.; Yamauchi, H.; Saito, Y.; Sasaki, T.; Morikubo, H.; Nagashima, Y.; Takagi, S.; Yamanaka, K.; et al. Heme oxygenase-1, a potential biomarker of chronic silicosis, attenuates silica-induced lung injury. Am. J. Respir. Crit. Care Med. 2006, 174, 906–914. [Google Scholar] [CrossRef]
- Shinohara, T.; Kaneko, T.; Nagashima, Y.; Ueda, A.; Tagawa, A.; Ishigatsubo, Y. Adenovirus-mediated transfer and overexpression of heme oxygenase 1 cDNA in lungs attenuates elastase-induced pulmonary emphysema in mice. Hum. Gene Ther. 2005, 16, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vos, W.M. Systems solutions by lactic acid bacteria: From paradigms to practice. Microb. Cell Fact. 2011, 10 (Suppl. 1), S2. [Google Scholar] [CrossRef] [Green Version]
- Cano-Garrido, O.; Seras-Franzoso, J.; Garcia-Fruitos, E. Lactic acid bacteria: Reviewing the potential of a promising delivery live vector for biomedical purposes. Microb. Cell Fact. 2015, 14, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.M.; Mercenier, A. Mucosal delivery of therapeutic and prophylactic molecules using lactic acid bacteria. Nat. Rev. Microbiol. 2008, 6, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fruitos, E. Lactic Acid Bacteria: A promising alternative for recombinant protein production. Microb. Cell Fact. 2012, 11, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolotin, A.; Wincker, P.; Mauger, S.; Jaillon, O.; Malarme, K.; Weissenbach, J.; Ehrlich, S.D.; Sorokin, A. The complete genome sequence of the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef] [Green Version]
- Daniel, C.; Roussel, Y.; Kleerebezem, M.; Pot, B. Recombinant lactic acid bacteria as mucosal biotherapeutic agents. Trends. Biotechnol. 2011, 29, 499–508. [Google Scholar] [CrossRef]
- Shigemori, S.; Watanabe, T.; Kudoh, K.; Ihara, M.; Nigar, S.; Yamamoto, Y.; Suda, Y.; Sato, T.; Kitazawa, H.; Shimosato, T. Oral delivery of Lactococcus lactis that secretes bioactive heme oxygenase-1 alleviates development of acute colitis in mice. Microb. Cell Fact. 2015, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Grossman, J. The evolution of inhaler technology. J. Asthma 1994, 31, 55–64. [Google Scholar] [CrossRef]
- Sato, T.; Shimosato, T. Development of a New Treatment Modality for Lung Diseases That Uses Innovative Fine Droplet Drying (fdd) Technology Offering Inhalable Nano/Microparticles-Incorporated Therapeutic Agents; Science Impact Ltd.: Bristol, UK, 2019; Volume 3. [Google Scholar]
- Dekhuijzen, P.N.; Vincken, W.; Virchow, J.C.; Roche, N.; Agusti, A.; Lavorini, F.; van Aalderen, W.M.; Price, D. Prescription of inhalers in asthma and COPD: Towards a rational, rapid and effective approach. Respir. Med. 2013, 107, 1817–1821. [Google Scholar] [CrossRef] [Green Version]
- Tashkin, D.P.; Celli, B.; Senn, S.; Burkhart, D.; Kesten, S.; Menjoge, S.; Decramer, M.; Investigators, U.S. A 4-year trial of tiotropium in chronic obstructive pulmonary disease. N. Engl. J. Med. 2008, 359, 1543–1554. [Google Scholar] [CrossRef] [Green Version]
- Graybill, J.R.; Ahrens, J.; Suchyta, M.; Coalson, J. Experimental pulmonary histoplasmosis and emphysema. Am. Rev. Respir. Dis. 1988, 137, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Shigemori, S.; Oshiro, K.; Wang, P.; Yamamoto, Y.; Wang, Y.; Sato, T.; Uyeno, Y.; Shimosato, T. Generation of dipeptidyl peptidase-IV-inhibiting peptides from beta-lactoglobulin secreted by Lactococcus lactis. Biomed Res. Int. 2014, 2014, 393598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigemori, S.; Yonekura, S.; Sato, T.; Otani, H.; Shimosato, T. Expression of the immunoreactive buckwheat major allergenic storage protein in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2013, 97, 3603–3611. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Villena, J.; Vintini, E.; Hebert, E.M.; Raya, R.; Alvarez, S. Nasal immunization with Lactococcus lactis expressing the pneumococcal protective protein A induces protective immunity in mice. Infect. Immun. 2008, 76, 2696–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomosada, Y.; Chiba, E.; Zelaya, H.; Takahashi, T.; Tsukida, K.; Kitazawa, H.; Alvarez, S.; Villena, J. Nasally administered Lactobacillus rhamnosus strains differentially modulate respiratory antiviral immune responses and induce protection against respiratory syncytial virus infection. BMC Immunol. 2013, 14, 40. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, K.; Sato, T.; Shigemori, S.; Shimosato, T.; Shinkai, M.; Kaneko, T. Regulatory role of heme oxygenase-1 in silica-induced lung injury. Respir. Res. 2018, 19, 144. [Google Scholar] [CrossRef]
- Sato, T.; Shimosato, T.; Ueda, A.; Ishigatsubo, Y.; Klinman, D.M. Intrapulmonary Delivery of CpG Microparticles Eliminates Lung Tumors. Mol. Cancer Ther. 2015, 14, 2198–2205. [Google Scholar] [CrossRef] [Green Version]
- Dunnill, M.S. Quantitative methods in the study of pulmonary pathology. Thorax 1962, 17, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Vanoirbeek, J.A.; Rinaldi, M.; De Vooght, V.; Haenen, S.; Bobic, S.; Gayan-Ramirez, G.; Hoet, P.H.; Verbeken, E.; Decramer, M.; Nemery, B.; et al. Noninvasive and invasive pulmonary function in mouse models of obstructive and restrictive respiratory diseases. Am. J. Respir. Cell Mol. Biol. 2010, 42, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.D. Animal models for COPD. Chest 2000, 117, 223S–227S. [Google Scholar] [CrossRef] [Green Version]
- Harata, G.; He, F.; Hiruta, N.; Kawase, M.; Kubota, A.; Hiramatsu, M.; Yausi, H. Intranasal administration of Lactobacillus rhamnosus GG protects mice from H1N1 influenza virus infection by regulating respiratory immune responses. Lett. Appl. Microbiol. 2010, 50, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Hashiba, T.; Suzuki, M.; Nagashima, Y.; Suzuki, S.; Inoue, S.; Tsuburai, T.; Matsuse, T.; Ishigatubo, Y. Adenovirus-mediated transfer of heme oxygenase-1 cDNA attenuates severe lung injury induced by the influenza virus in mice. Gene Ther. 2001, 8, 1499–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, S.; Suzuki, M.; Nagashima, Y.; Suzuki, S.; Hashiba, T.; Tsuburai, T.; Ikehara, K.; Matsuse, T.; Ishigatsubo, Y. Transfer of heme oxygenase 1 cDNA by a replication-deficient adenovirus enhances interleukin 10 production from alveolar macrophages that attenuates lipopolysaccharide-induced acute lung injury in mice. Hum. Gene Ther. 2001, 12, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Tsuburai, T.; Suzuki, M.; Nagashima, Y.; Suzuki, S.; Inoue, S.; Hasiba, T.; Ueda, A.; Ikehara, K.; Matsuse, T.; Ishigatsubo, Y. Adenovirus-mediated transfer and overexpression of heme oxygenase 1 cDNA in lung prevents bleomycin-induced pulmonary fibrosis via a Fas-Fas ligand-independent pathway. Hum. Gene Ther. 2002, 13, 1945–1960. [Google Scholar] [CrossRef]
- Shigemori, S.; Namai, F.; Yamamoto, Y.; Nigar, S.; Sato, T.; Ogita, T.; Shimosato, T. Genetically modified Lactococcus lactis producing a green fluorescent protein-bovine lactoferrin fusion protein suppresses proinflammatory cytokine expression in lipopolysaccharide-stimulated RAW 264.7 cells. J. Dairy Sci. 2017, 100, 7007–7015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzola, M.; MacNee, W.; Martinez, F.J.; Rabe, K.F.; Franciosi, L.G.; Barnes, P.J.; Brusasco, V.; Burge, P.S.; Calverley, P.M.; Celli, B.R.; et al. Outcomes for COPD pharmacological trials: From lung function to biomarkers. Eur. Respir. J. 2008, 31, 416–469. [Google Scholar] [CrossRef] [Green Version]
- Brusselle, G.G.; Bracke, K.R.; Maes, T.; D’Hulst, A.I.; Moerloose, K.B.; Joos, G.F.; Pauwels, R.A. Murine models of COPD. Pulm. Pharmacol. Ther. 2006, 19, 155–165. [Google Scholar] [CrossRef]
- Wright, J.L.; Cosio, M.; Churg, A. Animal models of chronic obstructive pulmonary disease. Am. J. Physiol. Lung. Cell Mol. Physiol. 2008, 295, L1–L15. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yumoto, K.; Sato, T.; Nakashima, K.; Namai, F.; Shigemori, S.; Shimosato, T.; Kaneko, T. Nasally Administered Lactococcus lactis Secreting Heme Oxygenase-1 Attenuates Murine Emphysema. Antioxidants 2020, 9, 1049. https://doi.org/10.3390/antiox9111049
Yumoto K, Sato T, Nakashima K, Namai F, Shigemori S, Shimosato T, Kaneko T. Nasally Administered Lactococcus lactis Secreting Heme Oxygenase-1 Attenuates Murine Emphysema. Antioxidants. 2020; 9(11):1049. https://doi.org/10.3390/antiox9111049
Chicago/Turabian StyleYumoto, Kentaro, Takashi Sato, Kentaro Nakashima, Fu Namai, Suguru Shigemori, Takeshi Shimosato, and Takeshi Kaneko. 2020. "Nasally Administered Lactococcus lactis Secreting Heme Oxygenase-1 Attenuates Murine Emphysema" Antioxidants 9, no. 11: 1049. https://doi.org/10.3390/antiox9111049