

Silybin Alleviated Hepatic Injury by Regulating Redox Balance, Inflammatory Response, and Mitochondrial Function in Weaned Piglets under Paraquat-Induced Oxidative Stress

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics Approval
2.2. Animals and Treatment
2.3. Sample Collection
2.4. Plasma Biochemical Analysis
2.5. Determination of Inflammatory Cytokine Content
2.6. Determination of Enzyme Activity
2.7. Adenosine Triphosphate Content Assay
2.8. Histopathological Staining
2.9. Real-Time Quantitative PCR Analysis (RT-qPCR)
2.10. Western Blotting Analysis
2.11. Statistical Analysis
3. Results
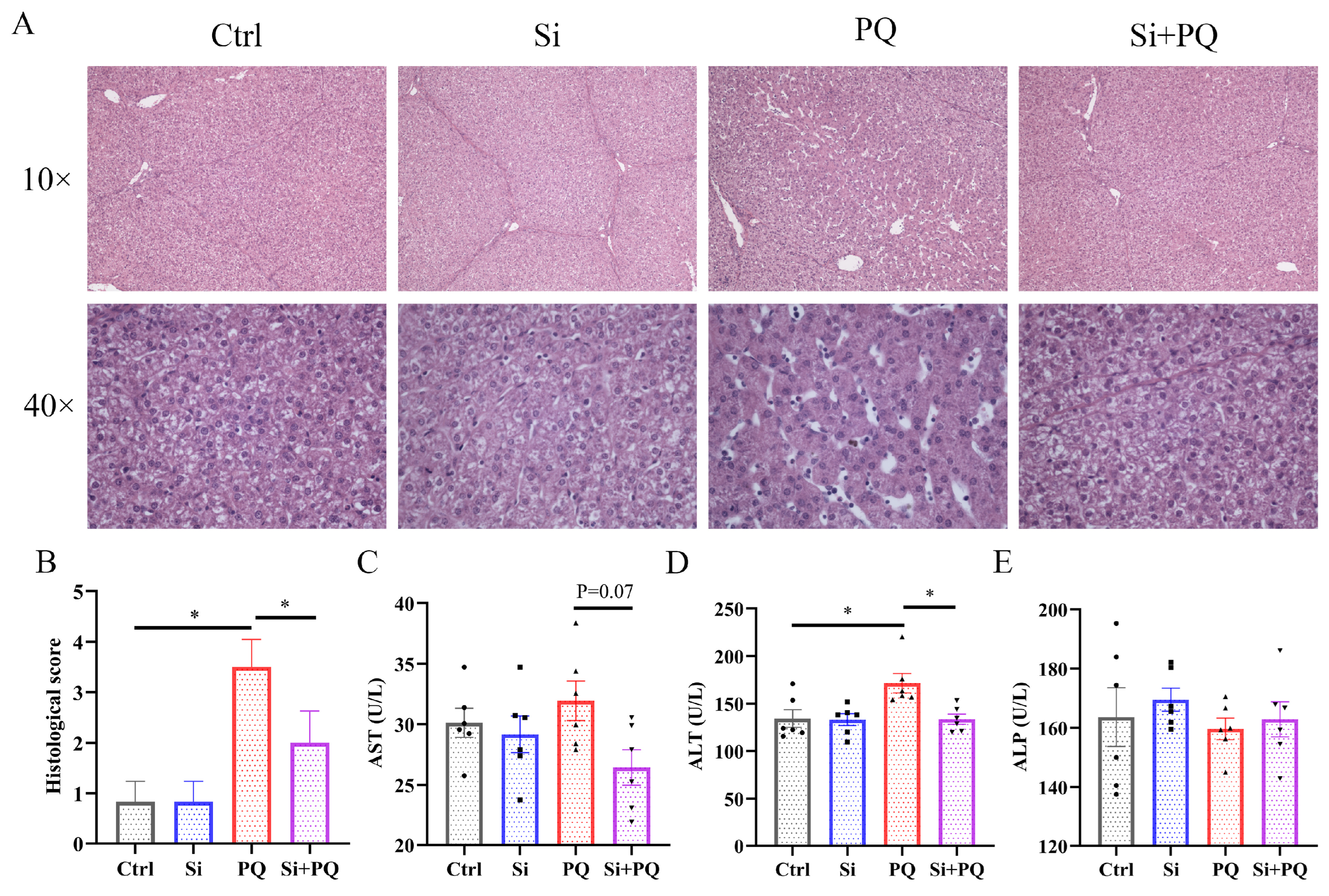
3.1. Effects of Dietary Silybin Supplementation on Liver Injury in Piglets Challenged with PQ
3.2. Effects of Dietary Silybin on Hepatic Oxidative Stress in Piglets Challenged with PQ
3.3. Effects of Dietary Silybin on Hepatic Activation of Nrf2 Signaling Pathway in Piglets Challenged with PQ
3.4. Effects of Dietary Silybin on Inflammatory Response in Piglets Challenged with PQ
3.5. Effects of Dietary Silybin on Mitochondrial Function in Piglets Challenged with PQ
3.6. Effects of Dietary Silybin Supplementation on Hepatocyte Apoptosis in Piglets Challenged with PQ
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, Y.; Li, Y.; Jia, P.; Ji, S.; Chen, Y.; Wang, T. Protective effects of pterostilbene against hepatic damage, redox imbalance, mitochondrial dysfunction, and endoplasmic reticulum stress in weanling piglets. J. Anim. Sci. 2020, 98, skaa328. [Google Scholar] [CrossRef] [PubMed]
- Machado, I.F.; Miranda, R.G.; Dorta, D.J.; Rolo, A.P.; Palmeira, C.M. Targeting Oxidative Stress with Polyphenols to Fight Liver Diseases. Antioxidants 2023, 12, 1212. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Paz, M.; Morán, L.; López-Alcántara, N.; Freixo, C.; Andrade, R.J.; Lucena, M.I.; Cubero, F.J. Oxidative Stress in Drug-Induced Liver Injury (DILI): From Mechanisms to Biomarkers for Use in Clinical Practice. Antioxidants 2021, 10, 390. [Google Scholar] [CrossRef]
- Wu, D.; Cederbaum, A.I. Oxidative stress and alcoholic liver disease. Semin. Liver Dis. 2009, 29, 141–154. [Google Scholar] [CrossRef]
- Yang, J.; Fernández-Galilea, M.; Martínez-Fernández, L.; González-Muniesa, P.; Pérez-Chávez, A.; Martínez, J.A.; Moreno-Aliaga, M.J. Oxidative Stress and Non-Alcoholic Fatty Liver Disease: Effects of Omega-3 Fatty Acid Supplementation. Nutrients 2019, 11, 872. [Google Scholar] [CrossRef]
- Guerbette, T.; Boudry, G.; Lan, A. Mitochondrial function in intestinal epithelium homeostasis and modulation in diet-induced obesity. Mol. Metab. 2022, 63, 101546. [Google Scholar] [CrossRef]
- Dong, Y.; Zhao, J.; Zhu, Q.; Liu, H.; Wang, J.; Lu, W. Melatonin inhibits the apoptosis of rooster Leydig cells by suppressing oxidative stress via AKT-Nrf2 pathway activation. Free Radic. Biol. Med. 2020, 160, 1–12. [Google Scholar] [CrossRef]
- Mooli, R.G.R.; Mukhi, D.; Ramakrishnan, S.K. Oxidative Stress and Redox Signaling in the Pathophysiology of Liver Diseases. Compr. Physiol. 2022, 12, 3167–3192. [Google Scholar]
- Surai, P.F. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef]
- Dobiasova, S.; Rehorova, K.; Kucerova, D.; Biedermann, D.; Kanova, K.; Petraskova, L.; Koucka, K.; Vaclavikova, R.; Valentova, K.; Ruml, T.; et al. Multidrug Resistance Modulation Activity of Silybin Derivatives and Their Anti-Inflammatory Potential. Antioxidants 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, W.; Zhai, T.; You, J.; Chen, Y. Silibinin ameliorates hepatic lipid accumulation and oxidative stress in mice with non-alcoholic steatohepatitis by regulating CFLAR-JNK pathway. Acta Pharm. Sin. B 2019, 9, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Salamone, F.; Volti, G.L. S1830 Silybin Enhances Mitochondrial Function and Inhibits NFkB Activation in Experimental NAFLD. Gastroenterology 2010, 138, S-798. [Google Scholar] [CrossRef]
- Serviddio, G.; Bellanti, F.; Stanca, E.; Lunetti, P.; Blonda, M.; Tamborra, R.; Siculella, L.; Vendemiale, G.; Capobianco, L.; Giudetti, A.M. Silybin exerts antioxidant effects and induces mitochondrial biogenesis in liver of rat with secondary biliary cirrhosis. Free Radic. Biol. Med. 2014, 73, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Lin, S.; Lin, Y.; Fang, Z.; Xu, S.; Feng, B.; Zhuo, Y.; Li, J.; Che, L.; Jiang, X.; et al. Effects of silymarin supplementation during transition and lactation on reproductive performance, milk composition and haematological parameters in sows. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Hwang, H.S.; Jang, S.Y.; Yu, S.; Kim, I.H. Supplemental impact of silymarin in growing pig diet on the growth performance, total tract digestibility, faecal microflora, faecal noxious gas emission and absorption rate in blood. J. Anim. Physiol. Anim. Nutr. 2024, 108, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, S.; Park, J.H.; Cho, S.; Kim, I.H. Silymarin seed extract supplementation enhances the growth performance, meat quality, and nutrients digestibility, and reduces gas emission in broilers. Anim. Biosci. 2022, 35, 1215–1222. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, B.; Lv, X.; Dong, Y.; Li, L.; Xia, Z. Effects of silybin supplementation on growth performance, serum indexes and liver transcriptome of Peking ducks. Front. Vet. Sci. 2024, 10, 1325115. [Google Scholar] [CrossRef]
- Cai, L.; Gao, G.; Yin, C.; Bai, R.; Li, Y.; Sun, W.; Pi, Y.; Jiang, X.; Li, X. The Effects of Dietary Silybin Supplementation on the Growth Performance and Regulation of Intestinal Oxidative Injury and Microflora Dysbiosis in Weaned Piglets. Antioxidants 2023, 12, 1975. [Google Scholar] [CrossRef]
- Chen, J.; Su, Y.; Lin, F.; Iqbal, M.; Mehmood, K.; Zhang, H.; Shi, D. Effect of paraquat on cytotoxicity involved in oxidative stress and inflammatory reaction: A review of mechanisms and ecological implications. Ecotox. Environ. Safe 2021, 224, 112711. [Google Scholar] [CrossRef]
- Dilberger, B.; Baumanns, S.; Schmitt, F.; Schmiedl, T.; Hardt, M.; Wenzel, U.; Eckert, G.P. Mitochondrial Oxidative Stress Impairs Energy Metabolism and Reduces Stress Resistance and Longevity of C. elegans. Oxid. Med. Cell. Longev. 2019, 2019, 6840540. [Google Scholar] [CrossRef]
- Zhang, R.; Zhao, C.; Gong, X.; Yang, J.; Zhang, G.; Zhang, W. Ginkgolide C Alleviates Acute Lung Injury Caused by Paraquat Poisoning via Regulating the Nrf2 and NF-κB Signaling Pathways. Oxid. Med. Cell. Longev. 2022, 2022, 7832983. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhang, Y.; Zhao, Z.; Shen, H.; Zhao, H.; Zhao, M. Isorhynchophylline ameliorates paraquat-induced acute kidney injury by attenuating oxidative stress and mitochondrial damage via regulating toll-interacting expression. Toxicol. Appl. Pharmacol. 2021, 420, 115521. [Google Scholar] [CrossRef]
- Liu, X.; Yang, H.; Liu, Z. Signaling pathways involved in paraquat-induced pulmonary toxicity: Molecular mechanisms and potential therapeutic drugs. Int. Immunopharmacol. 2022, 113, 109301. [Google Scholar] [CrossRef]
- Qi, M.; Wang, N.; Xiao, Y.; Deng, Y.; Zha, A.; Tan, B.; Wang, J.; Yin, Y.; Liao, P. Ellagic acid ameliorates paraquat-induced liver injury associated with improved gut microbial profile. Environ. Pollut. 2022, 293, 118572. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. Osteoarthr. Cartil. 2012, 20, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Dou, X.; Yan, D.; Liu, S.; Gao, L.; Shan, A. Thymol Alleviates LPS-Induced Liver Inflammation and Apoptosis by Inhibiting NLRP3 Inflammasome Activation and the AMPK-mTOR-Autophagy Pathway. Nutrients 2022, 14, 2809. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Wei, Z.; Zhao, X.; Li, Y.; Li, X.; Jiang, X. Gallic acid mitigates LPS-induced inflammatory response via suppressing NF-κB signalling pathway in IPEC-J2 cells. J. Anim. Physiol. Anim. Nutr. 2022, 106, 1000–1008. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.D.; de Freitas, M.L.; Dalmolin, L.; Oliveira, L.P.; Fleck, M.A.; Pagliarini, P.; Acker, C.; Roman, S.S.; Brandão, R. Diphenyl diselenide prevents hepatic alterations induced by paraquat in rats. Environ. Toxicol. Pharmacol. 2013, 36, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhang, Z.; Yang, S.; Wang, J.; Yang, X.; Tan, D. Betanin attenuates paraquat-induced liver toxicity through a mitochondrial pathway. Food Chem. Toxicol. 2014, 70, 100–106. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, X.; Li, L.; Wei, G.; Zhao, M. Hydrogen Sulfide Protects against Paraquat-Induced Acute Liver Injury in Rats by Regulating Oxidative Stress, Mitochondrial Function, and Inflammation. Oxid. Med. Cell. Longev. 2020, 2020, 6325378. [Google Scholar] [CrossRef] [PubMed]
- Baradaran, A.; Samadi, F.; Ramezanpour, S.S.; Yousefdoust, S. Hepatoprotective effects of silymarin on CCl4-induced hepatic damage in broiler chickens model. Toxicol. Rep. 2019, 6, 788–794. [Google Scholar] [CrossRef]
- Wan, J.-Y.; Gong, X.; Zhang, L.; Li, H.-Z.; Zhou, Y.-F.; Zhou, Q.-X. Protective effect of baicalin against Lipopolysaccharide/d-galactosamine-induced liver injury in mice by up-regulation of Heme oxygenase-1. Eur. J. Pharmacol. 2008, 587, 302–308. [Google Scholar] [CrossRef]
- Zhang, Q.; Ahn, J.M.; Kim, I.H. Micelle silymarin supplementation to sows’ diet from day 109 of gestation to entire lactation period enhances reproductive performance and affects serum hormones and metabolites. J. Anim. Sci. 2021, 99, skab354. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Li, S.; Jiang, N.; Shao, X.; Zhang, M.; Jin, H.; Zhang, Z.; Shen, J.; Zhou, Y.; Zhou, W.; et al. PINK1-parkin pathway of mitophagy protects against contrast-induced acute kidney injury via decreasing mitochondrial ROS and NLRP3 inflammasome activation. Redox Biol. 2019, 26, 101254. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Yin, R.X.; Gao, H.; Huang, F.; Wu, J.Z.; Pan, S.L.; Lin, W.X.; Yang, D.Z. Association of the SPTLC3 rs364585 polymorphism and serum lipid profiles in two Chinese ethnic groups. Lipids Health Dis. 2017, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Duan, G.; Huang, P.; Zheng, C.; Zheng, J.; Yu, J.; Zhang, P.; Wan, M.; Li, F.; Guo, Q.; Yin, Y.; et al. Development and Recovery of Liver Injury in Piglets by Incremental Injection of LPS. Antioxidants 2023, 12, 1143. [Google Scholar] [CrossRef]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef]
- Farmer, C.; Lapointe, J.; Palin, M.-F. Effects of the plant extract silymarin on prolactin concentrations, mammary gland development, and oxidative stress in gestating gilts1. J. Anim. Sci. 2014, 92, 2922–2930. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yamamoto, M. Molecular Mechanisms Activating the Nrf2-Keap1 Pathway of Antioxidant Gene Regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef]
- Ryan, S.W.; Quynh, T.T.; Samreen, F.; Carrie, H.S.; Thomas, W.K.; Thomas, R.S. Pharmacogenomics of Chemically Distinct Classes of Keap1-Nrf2 Activators Identify Common and Unique Gene, Protein, and Pathway Responses In Vivo. Mol. Pharmacol. 2018, 93, 297. [Google Scholar] [CrossRef]
- Hussein, R.M.; Sawy, D.M.; Kandeil, M.A.; Farghaly, H.S. Chlorogenic acid, quercetin, coenzyme Q10 and silymarin modulate Keap1-Nrf2/heme oxygenase-1 signaling in thioacetamide-induced acute liver toxicity. Life Sci. 2021, 277, 119460. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Y.; Liu, D. Silibinin ameliorats H2O2-induced cell apoptosis and oxidative stress response by activating Nrf2 signaling in trophoblast cells. Acta Histochem. 2020, 122, 151620. [Google Scholar] [CrossRef]
- Vezza, T.; Molina-Tijeras, J.A.; Rodríguez-Nogales, A.; Garrido-Mesa, J.; Cádiz-Gurrea, M.d.l.L.; Segura-Carretero, A.; González-Tejero, M.R.; Rodríguez-Cabezas, M.E.; Gálvez, J.; Algieri, F. The Antioxidant Properties of Salvia verbenaca Extract Contribute to Its Intestinal Antiinflammatory Effects in Experimental Colitis in Rats. Antioxidants 2023, 12, 2071. [Google Scholar] [CrossRef]
- Cui, W.; Zhou, S.; Wang, Y.; Shi, X.; Liu, H. Cadmium exposure activates the PI3K/AKT signaling pathway through miRNA-21, induces an increase in M1 polarization of macrophages, and leads to fibrosis of pig liver tissue. Ecotoxicol. Environ. Saf. 2021, 228, 113015. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, Q.-H.; Li, Y.-X.; Huang, Y.-F.; Xie, J.-H.; Xu, L.-Q.; Dou, Y.-X.; Su, Z.-R.; Zeng, H.-F.; Chen, J.-N. Protective effects of silymarin on triptolide-induced acute hepatotoxicity in rats. Mol. Med. Rep. 2018, 17, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Jiang, X.; Jia, X.; Jiang, X.; Che, L.; Lin, Y.; Zhuo, Y.; Feng, B.; Fang, Z.; Li, J.; et al. Silymarin Modulates Microbiota in the Gut to Improve the Health of Sow from Late Gestation to Lactation. Animals 2022, 12, 2202. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, J.U.; Gomez-Quiroz, L.; Arreguin Camacho, L.O.; Pinna, F.; Lee, Y.-H.; Kitade, M.; Domínguez, M.P.; Castven, D.; Breuhahn, K.; Conner, E.A.; et al. Curcumin effectively inhibits oncogenic NF-κB signaling and restrains stemness features in liver cancer. J. Hepatol. 2015, 63, 661–669. [Google Scholar] [CrossRef]
- Yoon, W.J.; Moon, J.Y.; Song, G.; Lee, Y.K.; Han, M.S.; Lee, J.S.; Ihm, B.S.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Artemisia fukudo essential oil attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2010, 48, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Abhilash, P.A.; Harikrishnan, R.; Indira, M. Ascorbic acid is superior to silymarin in the recovery of ethanol-induced inflammatory reactions in hepatocytes of guinea pigs. J. Physiol. Biochem. 2013, 69, 785–798. [Google Scholar] [CrossRef]
- Sharma, S.; Le Guillou, D.; Chen, J.Y. Cellular stress in the pathogenesis of nonalcoholic steatohepatitis and liver fibrosis. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 662–678. [Google Scholar] [CrossRef]
- Nunnari, J.; Suomalainen, A. Mitochondria: In Sickness and in Health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef]
- Bellance, N.; Lestienne, P.; Rossignol, R. Mitochondria: From bioenergetics to the metabolic regulation of carcinogenesis. Front. Biosci.-Landmark 2009, 14, 4015–4034. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; Ji, S.; Jia, P.; Chen, Y.; Li, Y.; Wang, T. Resveratrol and its derivative pterostilbene attenuate oxidative stress-induced intestinal injury by improving mitochondrial redox homeostasis and function via SIRT1 signaling. Free Radic. Biol. Med. 2021, 177, 1–14. [Google Scholar] [CrossRef]
- Anand, R.; Wai, T.; Baker, M.J.; Kladt, N.; Schauss, A.C.; Rugarli, E.; Langer, T. The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission. J. Cell Biol. 2014, 204, 919–929. [Google Scholar] [CrossRef]
- Tie, F.; Fu, Y.; Hu, N.; Wang, H. Silibinin Protects against H2O2-Induced Oxidative Damage in SH-SY5Y Cells by Improving Mitochondrial Function. Antioxidants 2022, 11, 1101. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Wang, Y.-K.; Hong, Y.-J.; Huang, Z.-Q. Protective effects of silybin on human umbilical vein endothelial cell injury induced by H2O2 in vitro. Vasc. Pharmacol. 2005, 43, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.F.A.; Wong-Brown, M.W.; Bowden, N.A. BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis. 2019, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Abd Eldaim, M.A.; Barakat, E.R.; Alkafafy, M.; Elaziz, S.A.A. Antioxidant and anti-apoptotic prophylactic effect of silymarin against lead-induced hepatorenal toxicity in rats. Environ. Sci. Pollut. Res. 2021, 28, 57997–58006. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, M.; Wang, Y.; Zhang, L.; Zhao, H.; Zhao, M. Silymarin attenuated paraquat-induced cytotoxicity in macrophage by regulating Trx/TXNIP complex, inhibiting NLRP3 inflammasome activation and apoptosis. Toxicol. Vitr. 2018, 46, 265–272. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Accession Number | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|
| GAPDH | NM_001206359.1 | F: GCTTGTCATCAATGGAAAGG | 86 |
| R: CATACGTAGCACCAGCATCA | |||
| TNF-α | NM_214022.1 | F: CTCACGTCCTTCTGGTTTAG | 96 |
| R: CCCTGATTTCTAAGTGTTGC | |||
| IL-6 | NM_214399.1 | F: AATGTCGAGGCTGTGCAGATT | 82 |
| R: TGGTGGCTTTGTCTGGATTCT | |||
| IL-8 | NM_213867.1 | F: CCGTGTCAACATGACTTCCAA | 75 |
| R: GCCTCACAGAGAGCTGCAGAA | |||
| IL-10 | NM_214041.1 | F: GACGATGAAGATGAGGAAGA | 54 |
| R: AGGTTTTTCTTTGGTTTCCC | |||
| CAT | NM_214301.2 | F: CCTGCAACGTTCTGTAAGGC | 72 |
| R: GCTTCATCTGGTCACTGGCT | |||
| SOD1 | NM_001190422.1 | F: GAAGACAGTGTTAGTAACGG | 93 |
| R: CAGCCTTGTGTATTATCTCC | |||
| GPX1 | NM_214201.1 | F: TCTCCAGTGTGTCGCAATGA | 104 |
| R: TCGATGGTCAGAAAGCGACG | |||
| GPX4 | NM_214407.1 | F: GATTCTGGCCTTCCCTTGC | 173 |
| R: TCCCCTTGGGCTGGACTTT | |||
| NDUFS2 | XM_005663166.3 | F: CTAAACGCGCAGAGATGAAGA | 108 |
| R: CCTCAATGGCAGTGTATGTGG | |||
| NDUFV2 | NM_001097475.2 | F: CCCAGATACTCCATTTGATTTCA | 169 |
| R: AATTTCTGCCACCTTGTTCATG | |||
| SDHA | XM_021076930.1 | F: TCTCTGAGGCCGGGTTTAACACA | 124 |
| R: CACCTCCAGTTGTCCTCCTCCAT | |||
| UQCRB | NM_001185172.1 | F: GGATGACGATGTAAAAGAAGCCA | 141 |
| R: TCCTCCTCATATTTTGTCCACTG | |||
| ATP5H | XM_021066093.1 | F: CATTGACTGGGTAGCCTTTG | 115 |
| R: CTTCTCAGGTAGAGCAGCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, L.; Ming, D.; Chen, W.; Zhao, Y.; Li, Y.; Sun, W.; Pi, Y.; Jiang, X.; Li, X. Silybin Alleviated Hepatic Injury by Regulating Redox Balance, Inflammatory Response, and Mitochondrial Function in Weaned Piglets under Paraquat-Induced Oxidative Stress. Antioxidants 2024, 13, 324. https://doi.org/10.3390/antiox13030324
Cai L, Ming D, Chen W, Zhao Y, Li Y, Sun W, Pi Y, Jiang X, Li X. Silybin Alleviated Hepatic Injury by Regulating Redox Balance, Inflammatory Response, and Mitochondrial Function in Weaned Piglets under Paraquat-Induced Oxidative Stress. Antioxidants. 2024; 13(3):324. https://doi.org/10.3390/antiox13030324
Chicago/Turabian StyleCai, Long, Dongxu Ming, Wenning Chen, Ying Zhao, Yanpin Li, Wenjuan Sun, Yu Pi, Xianren Jiang, and Xilong Li. 2024. "Silybin Alleviated Hepatic Injury by Regulating Redox Balance, Inflammatory Response, and Mitochondrial Function in Weaned Piglets under Paraquat-Induced Oxidative Stress" Antioxidants 13, no. 3: 324. https://doi.org/10.3390/antiox13030324